Előadást letölteni
Az előadás letöltése folymat van. Kérjük, várjon

2
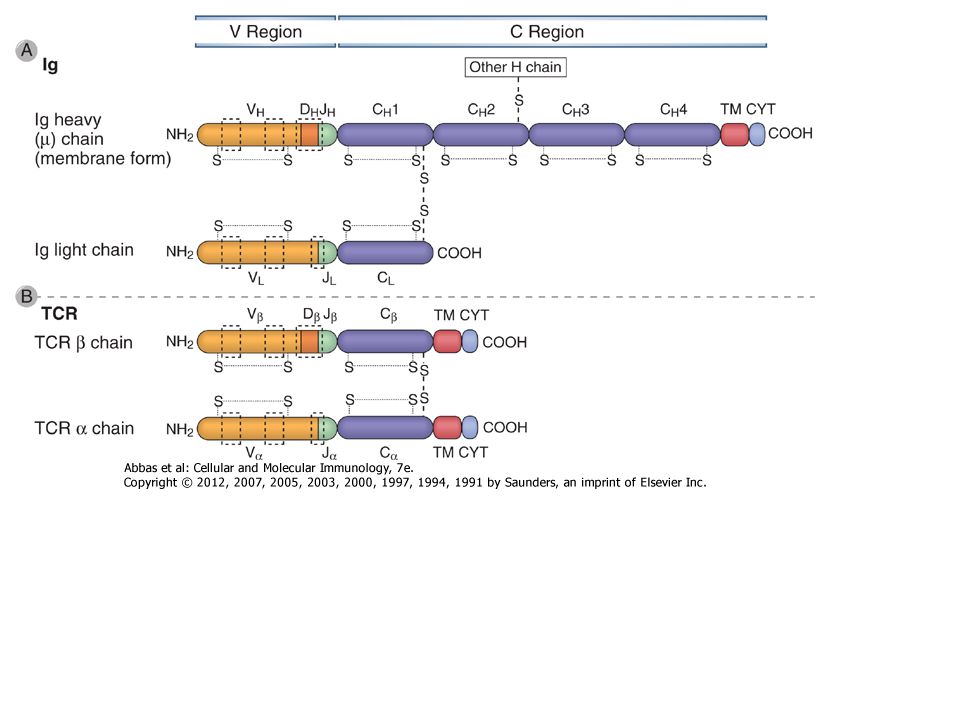
Az immunoglobulin szerkezete
Könnyű és nehéz láncok Diszulfid hidak láncok közötti láncon belüli Variabilis és konstans régiók Kapocs régió Domének VL & CL VH & CH1 - CH (CH4 pl. IgM) Oligoszaharid
 Oligoszaharid.")
3
HIPERVARIÁBILIS RÉGIÓK
könnyű lánc hipervariábilis régiója könnyű lánc H nehéz lánc CDR2 CDR1 CDR3 nehéz lánc hipervariábilis régiója CDR1 CDR3 CDR2 L

4
Hiper variábilis régiók---Complementary Determining Region (CDR)
A B-sejtek És a T-sejtek antigén felismerő receptorain is vannak

5
ADAPTÍV IMMUNITÁS SEJTJEI A LIMFOCITÁK SOKFÉLESÉGE
Hányféle SPECIFICITÁS ? Kb millió különböző ( ) antigén felismerő receptorral rendelkező egyedi B-limfocita klón Kb. 10 – 1000 millió különböző ( ) antigén felismerő receptorral rendelkező egyedi T-limfocita klón
 antigén felismerő receptorral rendelkező egyedi B-limfocita klón. Kb. 10 – 1000 millió különböző ( ) antigén felismerő receptorral rendelkező egyedi T-limfocita klón.")
6
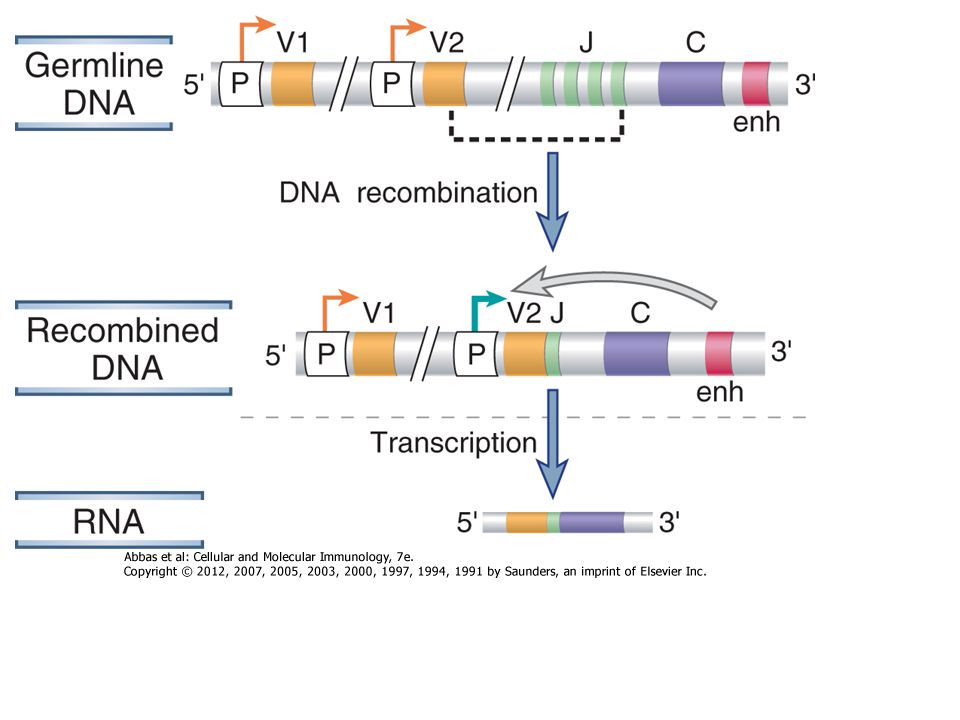
Hogyan képes a genom mintegy 20,000 génje több millió
Különböző specificitású ellenanyag kódolására? Egymástól elválasztott gén-szegmensekből csak egy funkcionális B-sejt receptor szerelődik össze minden B sejten

7
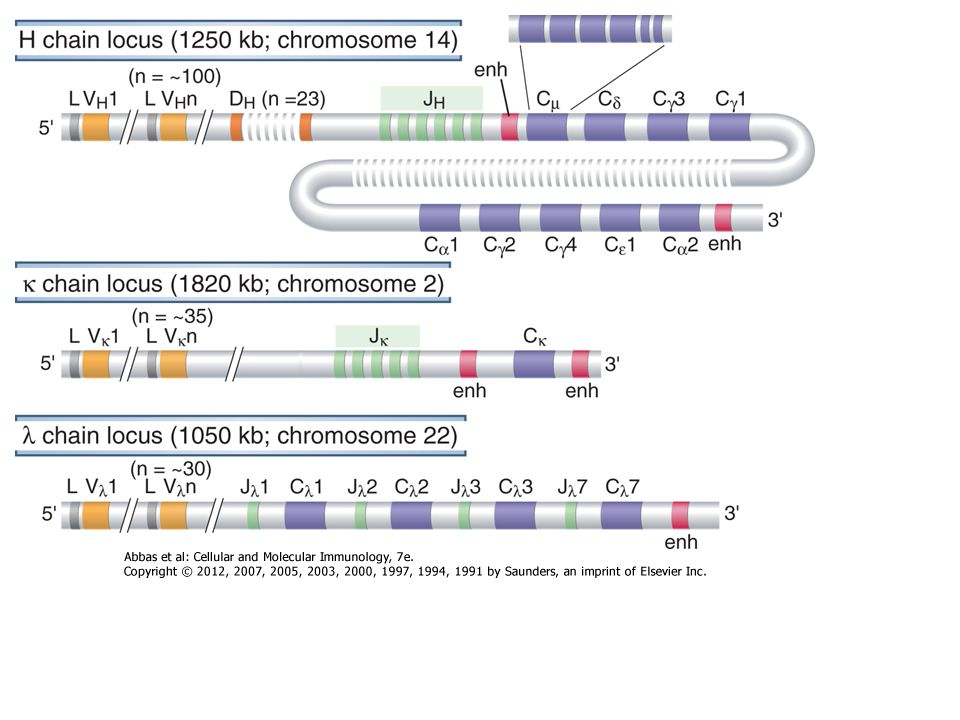
AZ IMMUNOGLOBULINOK MOLEKULÁRIS GENETIKÁJA
Dreyer & Bennett feltételezése (1965) Egy adott izotípusú ellenanyag valószínűleg: Az egyetlen C régiót kódoló gén (szegmens) a csíravonalban (ivarsejtben levő örökítő anyag) el van választva a V régió génektől A V génekből többféle áll rendelkezésre Feltételezhető egy mechanizmus, amely a V és C géneket fúzionálja egy teljes immunglobulin génné A feltételezés ellentétben állt az akkor elfogadott nézettel, amennyiben a DNS (genetikai állomány) egy adott egyed minden sejtjében azonos 7
 Egy adott izotípusú ellenanyag valószínűleg: Az egyetlen C régiót kódoló gén (szegmens) a csíravonalban (ivarsejtben levő örökítő anyag) el van választva a V régió génektől. A V génekből többféle áll rendelkezésre. Feltételezhető egy mechanizmus, amely a V és C géneket fúzionálja egy teljes immunglobulin génné. A feltételezés ellentétben állt az akkor elfogadott nézettel, amennyiben a DNS (genetikai állomány) egy adott egyed minden sejtjében azonos. 7.")
8
! A VH (nehéz lánc variábilis domén) GÉN SZEGMENSEK SZOMATIKUS ÁTRENDEZŐDÉSE 65 VH 27 D 6 JH VH1 VH2 VH3 D D D D JH JH JH JH D VH1 VH2 VH3 A B sejt fejlődés során JH VH1 VH2 JH D

9
AZ ANTIGÉN RECEPTOROK SOKFÉLESÉGE Egy egyed különböző B limfocitái
! AZ ANTIGÉN RECEPTOROK SOKFÉLESÉGE Egy egyed különböző B limfocitái VH D JH VL JL V-Domének C-Domének Néhány effektor funkció!! VH-D-JH VL-JL

13
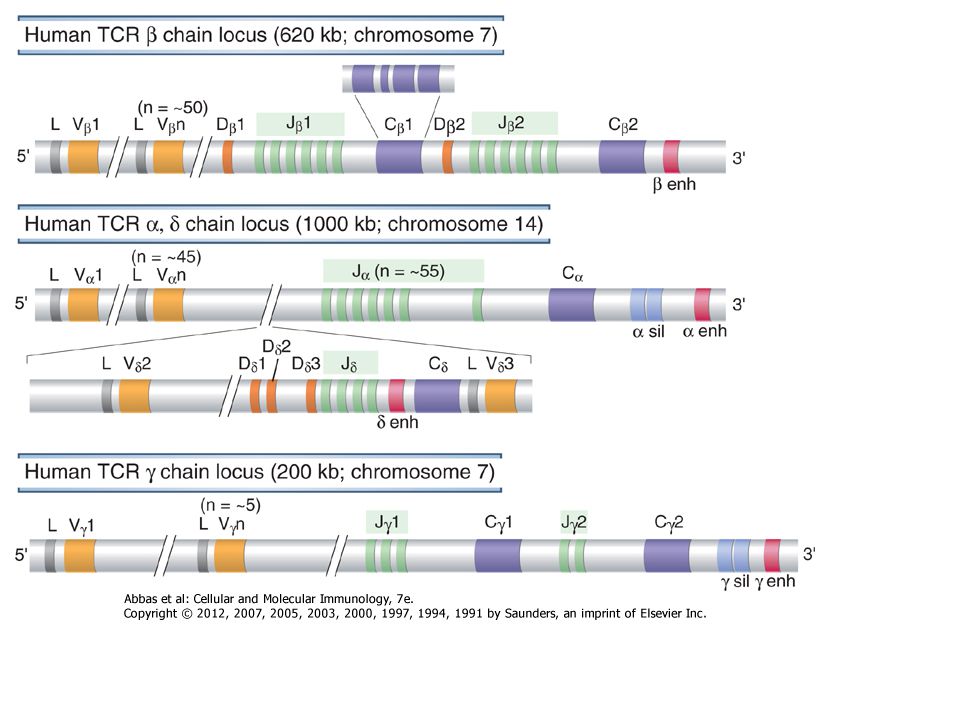
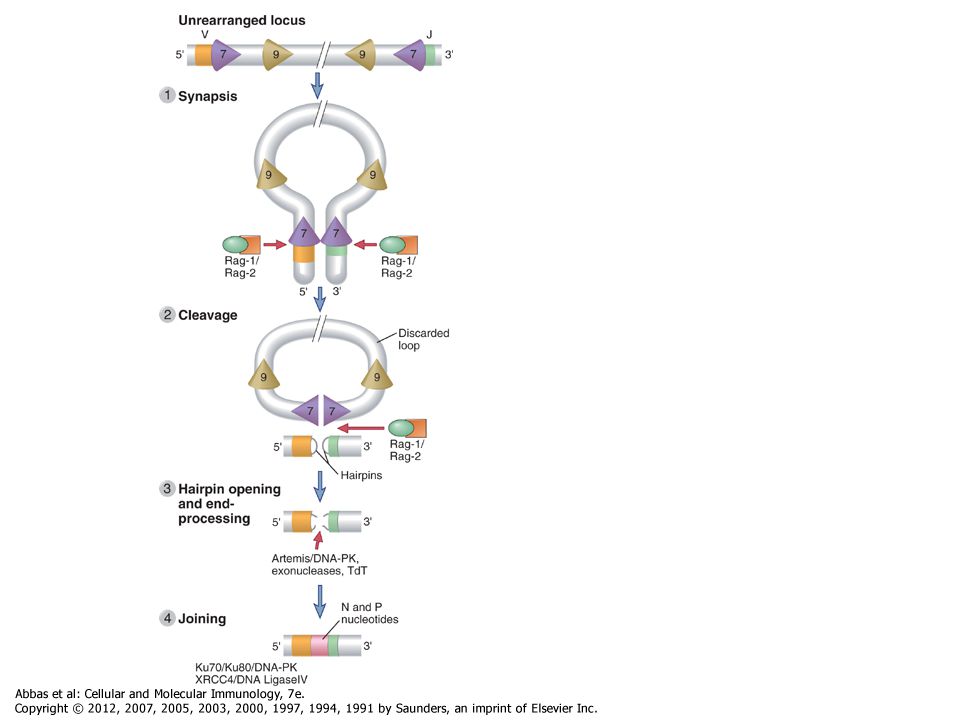
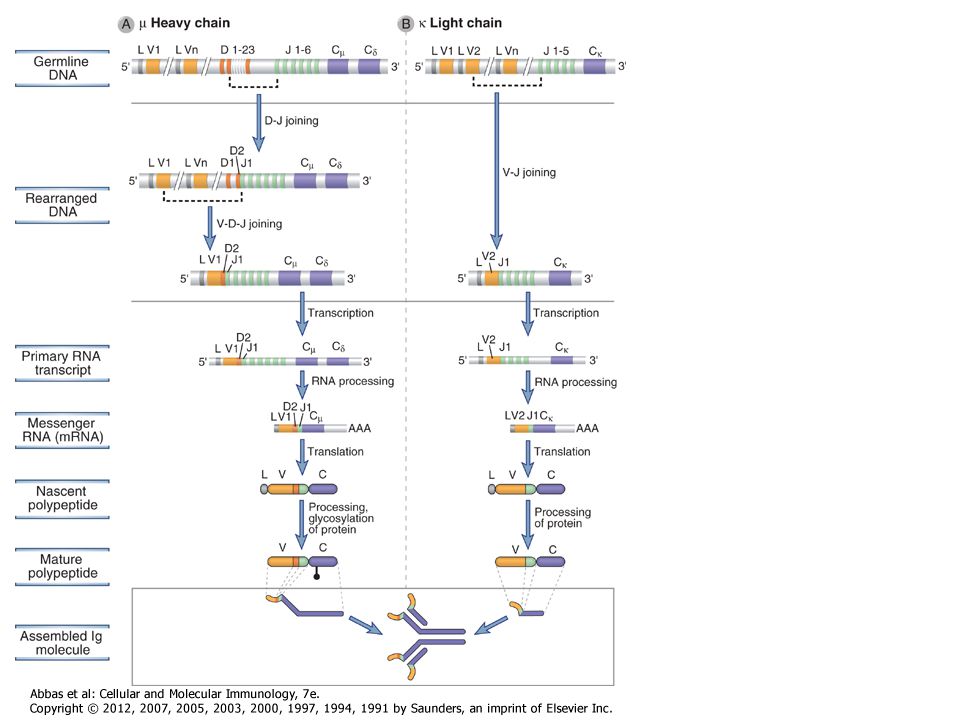
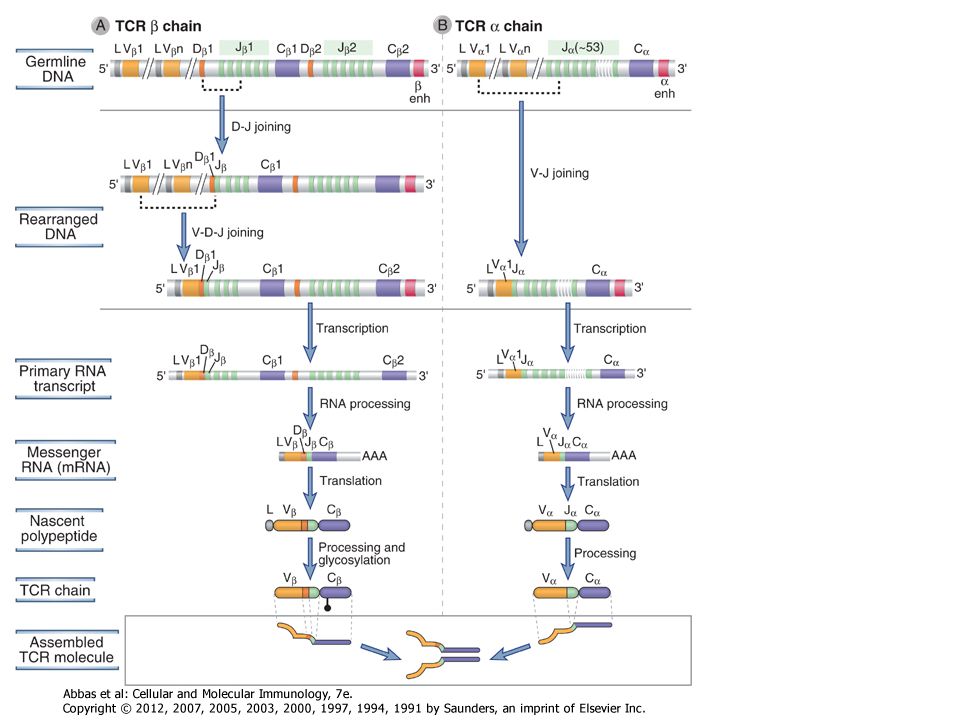
The process of V(D)J recombination at any Ig or TCR locus involves selection of one V gene, one J segment, and one D segment (when present) in each lymphocyte and rearrangement of these gene segments together to form a single V(D)J exon that will code for the variable region of an antigen receptor protein (Fig. 8-9). In the Ig light chain and TCR α and γ loci, which lack D segments, a single rearrangement event joins a randomly selected V gene to an equally randomly selected J segment. The IgH and TCR β and δ loci contain D segments, and at these loci two distinct rearrangement events must be separately initiated, first joining a D to a J and then a V segment to the fused DJ segment. Each rearrangement event involves a number of sequential steps. First, the chromatin must be opened in specific regions of the antigen receptor chromosome to make gene segments accessible to the enzymes that mediate recombination. Next, two selected gene segments must be brought next to one another across a considerable chromosomal distance. Double-stranded breaks are then introduced at the coding ends of these two segments, nucleotides are added or removed at the broken ends, and finally the processed ends are ligated to produce clonally unique but diverse antigen receptor genes that can be efficiently transcribed. The C regions lie downstream of the rearranged V(D)J exon separated by the germline J-C intron. This rearranged exon is transcribed to form a primary (nuclear) RNA transcript. Subsequent RNA splicing brings together the leader exon, the V(D)J exon, and the C region exons, forming an mRNA that can be translated on membrane-bound ribosomes to produce one of the chains of the antigen receptor.
. In the Ig light chain and TCR α and γ loci, which lack D segments, a single rearrangement event joins a randomly selected V gene to an equally randomly selected J segment. The IgH and TCR β and δ loci contain D segments, and at these loci two distinct rearrangement events must be separately initiated, first joining a D to a J and then a V segment to the fused DJ segment. Each rearrangement event involves a number of sequential steps. First, the chromatin must be opened in specific regions of the antigen receptor chromosome to make gene segments accessible to the enzymes that mediate recombination. Next, two selected gene segments must be brought next to one another across a considerable chromosomal distance. Double-stranded breaks are then introduced at the coding ends of these two segments, nucleotides are added or removed at the broken ends, and finally the processed ends are ligated to produce clonally unique but diverse antigen receptor genes that can be efficiently transcribed. The C regions lie downstream of the rearranged V(D)J exon separated by the germline J-C intron. This rearranged exon is transcribed to form a primary (nuclear) RNA transcript. Subsequent RNA splicing brings together the leader exon, the V(D)J exon, and the C region exons, forming an mRNA that can be translated on membrane-bound ribosomes to produce one of the chains of the antigen receptor..")
15
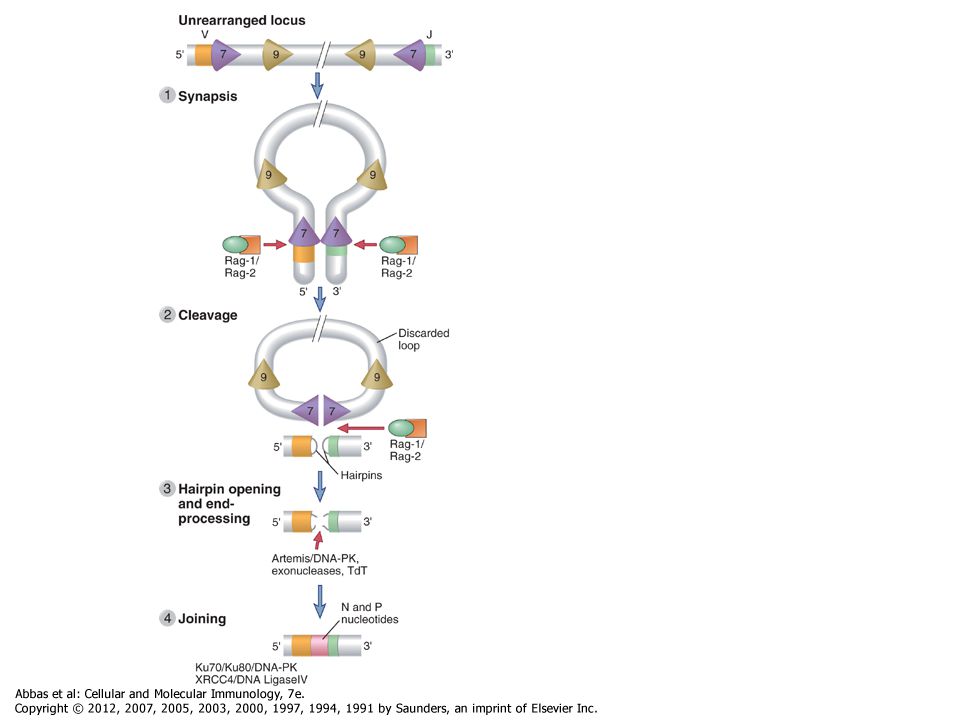
Figure 8-10 V(D)J recombination
Figure 8-10 V(D)J recombination. The DNA sequences and mechanisms involved in recombination in the Ig gene loci are depicted. The same sequences and mechanisms apply to recombinations in the TCR loci. A, Conserved heptamer (7 bp) and nonamer (9 bp) sequences, separated by 12- or 23-bp spacers, are located adjacent to V and J exons (for κ and λ loci) or to V, D, and J exons (in the H chain locus). The V(D)J recombinase recognizes these recombination signal sequences and brings the exons together. B, C, Recombination of V and J exons may occur by deletion of intervening DNA and ligation of the V and J segments (B) or, if the V gene is in the opposite orientation, by inversion of the DNA followed by ligation of adjacent gene segments (C). Red arrows indicate the sites where germline sequences are cleaved before their ligation to other Ig or TCR gene segments
J recombination. The DNA sequences and mechanisms involved in recombination in the Ig gene loci are depicted. The same sequences and mechanisms apply to recombinations in the TCR loci. A, Conserved heptamer (7 bp) and nonamer (9 bp) sequences, separated by 12- or 23-bp spacers, are located adjacent to V and J exons (for κ and λ loci) or to V, D, and J exons (in the H chain locus). The V(D)J recombinase recognizes these recombination signal sequences and brings the exons together. B, C, Recombination of V and J exons may occur by deletion of intervening DNA and ligation of the V and J segments (B) or, if the V gene is in the opposite orientation, by inversion of the DNA followed by ligation of adjacent gene segments (C). Red arrows indicate the sites where germline sequences are cleaved before their ligation to other Ig or TCR gene segments.")
18
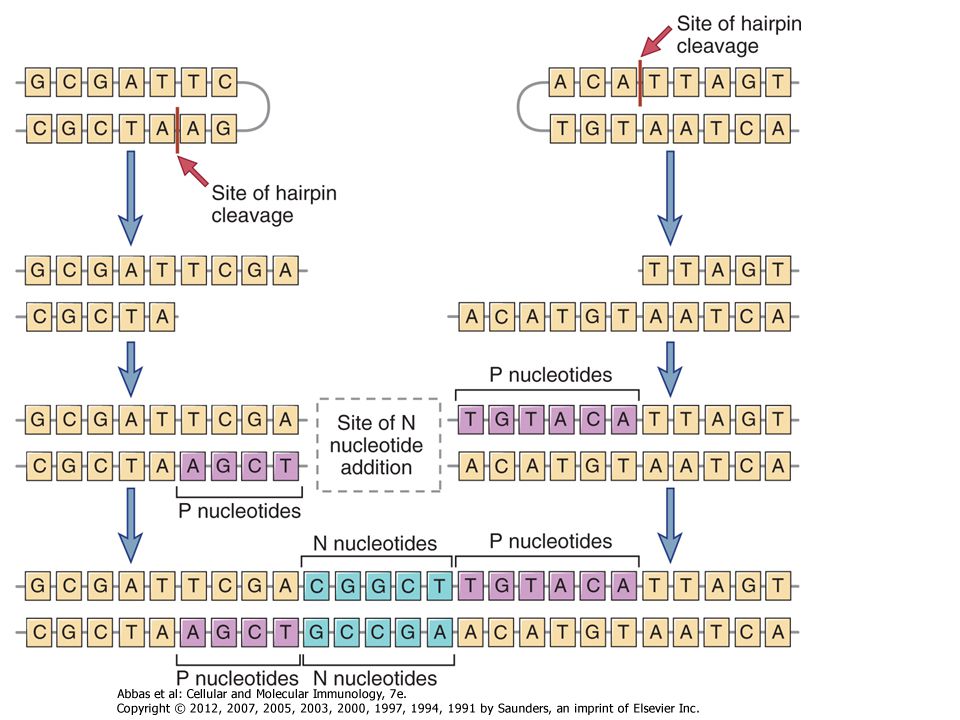
V D J TCGACGTTATAT AGCTGCAATATA TTTTT Kapcsolási sokféleség
Csíravonalban kódolt nukleotidok Palindrom (P) nukleotidok – újak Nem-template (N) kódolt nukleotidok - újak A V, D és J régiók között gyakorlatilag random szekvenciák alakulnak ki
 nukleotidok – újak. Nem-template (N) kódolt nukleotidok - újak. A V, D és J régiók között gyakorlatilag random szekvenciák alakulnak ki.")
22
B SEJT DIFFERENCIÁCIÓ A CSONTVELŐBEN

23
AZ IMMUNOGLOBULIN GÉN SZEGMENSEK ÁTRENDEZŐDÉSÉNEK SORRENDJE
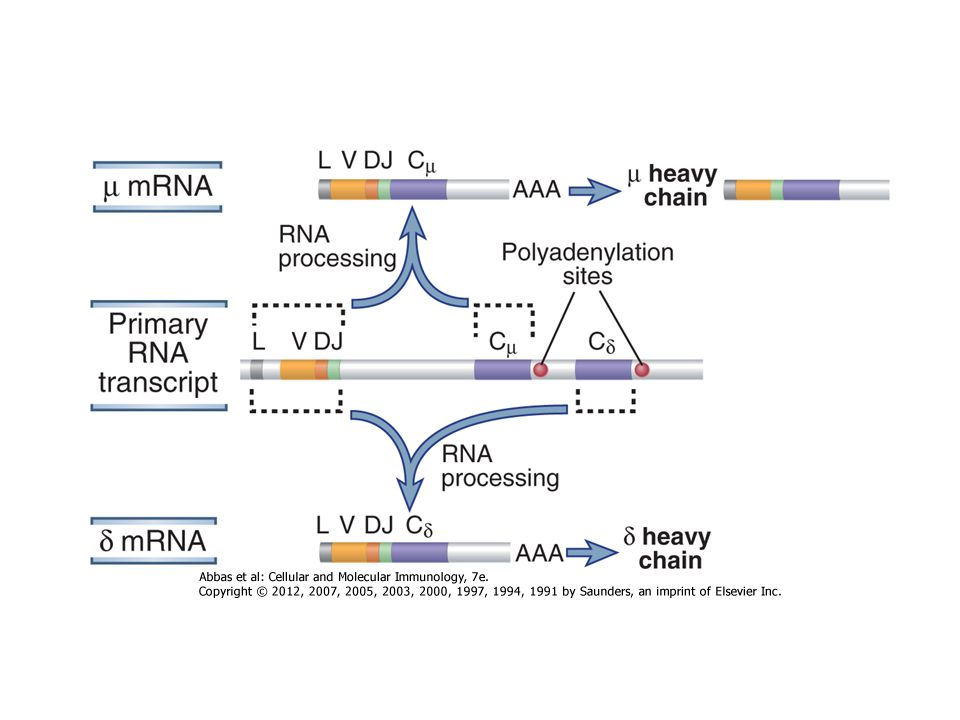
D – J rekombináció V – DJ rekombináció VDJ – δ transzkripció δ transzláció Pót könnyű lánc V – J rekombináció VJ – (vagy VJ - ) transzkripció vagy transzláció B-sejt Szecernált IgM ellenanyag mIgD mIgM
 transzkripció. vagy transzláció. B-sejt. Szecernált IgM ellenanyag. mIgD mIgM.")
24
Hogyan képes a B sejt csak egyfajta könnyű és egyfajta nehéz láncot kifejezni?
Annak ellenére, hogy minden B sejtben egy apai és egy anyai Ig lókusz is jelen van A gének nagy része ko-dominánsan kifejeződik Sikeres génátrendeződés gátolja a rekombinációt a másik kromoszómán ALLÉL KIZÁRÁS Csak egyféle H- és L-lánc szintetizálódik A sejtfelszínre 2 H- és 2 L-láncból álló dimerek kerülnek ki

25
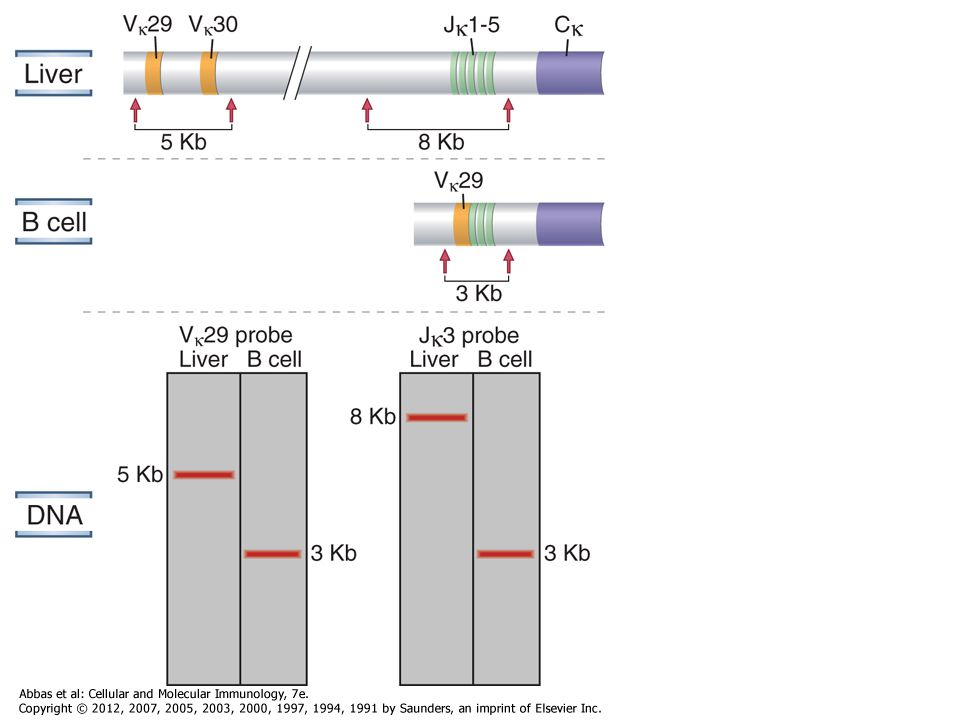
BIZONYÍTÉK AZ ALLÉL KIZÁRÁSRA
ALLOTÍPUS- a H-lánc C-régió polimorfizmusa (marker) Az Ig allotípusok a B-sejtek felszínén fluoreszcens festékkel kimutathatók a/a b/b a/b Y B a Y B b Y B a Y B b ÉS Y B a b H-lánc átrendeződés gátlása megakadályozza, hogy egy B-limfocitában kétféle specificitás alakuljon ki
 Az Ig allotípusok a B-sejtek felszínén fluoreszcens festékkel kimutathatók. a/a. b/b. a/b. Y. B. a. Y. B. b. Y. B. a. Y. B. b. ÉS. Y. B. a. b. H-lánc átrendeződés gátlása megakadályozza, hogy egy B-limfocitában kétféle specificitás alakuljon ki.")
26
AZ ALLÉL KIZÁRÁS SZÜKSÉGES A HATÉKONY KLONÁLIS SZELEKCIÓHOZ
Antibody S. typhi S. typhi Minden utódsejtnek ugyanazzal a specificitással kell rendelkeznie, máskülönben az immunválasz hatásfoka romlik A H-lánc átrendeződés gátlása az osztódó utódsejtekben megakadályozza az új specifitások létrejöttét

27
Y Y Y Y Y Y Az allél kizárás megakadályozza a
nem kívánatos specifitású ellenanyagok képződését Egyféle Ag receptor/sejt HA egy sejtben kétféle Ag receptor lenne Y B Y Saját antigen pl. agysejten B Y S. aureus S. aureus Y Anti S. aureus Ab Y Anti- agysejt Ab Y Anti S. aureus Ab A H-lánc átrendeződés gátlása biztosítja, hogy egy sejtben csak egyféle ellenanyag képződjön Ezáltal megakadályozza a nem kívánatos auto-reaktív választ

28
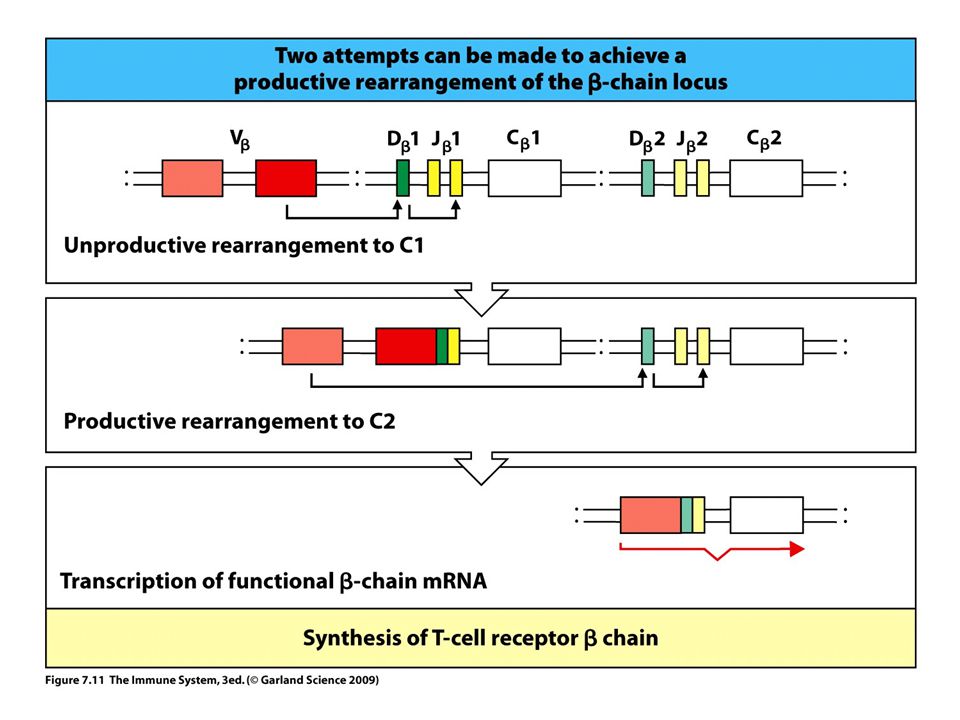
A GÉNÁTRENDEZŐDÉS LEHETŐSÉGEI
V-D-J 1.allél 2.allél mRNS Fehérje receptor Átrendeződés gátlása V-J V-J Nem sikeres Sikeres mRNS fehérje mRNS fehérje

29
A T SEJTEK ÉRÉSE ÉS DIFFERENCIÁLÓDÁSA A TÍMUSZBAN

33
The use of different combinations of V, D, and J gene segments and the addition and removal of nucleotides at the joints contribute to the tremendous diversity of antigen receptors, Combinatorial diversity. V(D)J rearrangement brings together multiple germline gene segments that may combine randomly, and different combinations produce different antigen receptors. The maximum possible number of combinations of these gene segments is the product of the numbers of V, J, and (if present) D gene segments at each antigen receptor locus. Therefore, the amount of combinatorial diversity that can be generated at each locus reflects the number of germline V, J, and D gene segments at that locus. After synthesis of antigen receptor proteins, combinatorial diversity is further enhanced by the juxtaposition of two different, randomly generated V regions (i.e., VH and VL in Ig molecules and Vα and Vβ in TCR molecules). Therefore, the total combinatorial diversity is theoretically the product of the combinatorial diversity of each of the two associating chains. The actual degree of combinatorial diversity in the expressed Ig and TCR repertoires in any individual is likely to be considerably less than the theoretical maximum. This is because not all recombinations of gene segments are equally likely to occur, and not all pairings of Ig heavy and light chains or TCR α and β chains may form functional antigen receptors. Importantly, because the numbers of V, D, and J segments in each locus are limited (see Table 8-1), the maximum possible numbers of combinations are on the order of thousands. This is, of course, much less than the actual diversity of antigen receptors in mature lymphocytes. Junctional diversity. The largest contribution to the diversity of antigen receptors is made by the removal or addition of nucleotides at the junctions of the V and D, D and J, or V and J segments at the time these segments are joined. One way in which this can occur is if endonucleases remove nucleotides from the germline sequences at the ends of the recombining gene segments. In addition, new nucleotide sequences, not present in the germline, may be added at junctions. As described earlier, coding segments (e.g., V and J gene segments) that are cleaved by Rag-1 form hairpin loops whose ends are often cleaved asymmetrically by the enzyme Artemis so that one DNA strand is longer than the other (Fig. 8-13). The shorter strand has to be extended with nucleotides complementary to the longer strand before the ligation of the two segments. The short lengths of added nucleotides are called P nucleotides, and their templated addition introduces new sequences at the V-D-J junctions. Another mechanism of junctional diversity is the random addition of up to 20 non-template-encoded nucleotides called N nucleotides (see Fig. 8-13). N region diversification is more common in Ig heavy chains and in TCR β and γ chains than in Ig κ or λ chains. This addition of new nucleotides is mediated by the enzyme terminal deoxynucleotidyl transferase (TdT). In mice rendered deficient in TdT by gene knockout, the diversity of B and T cell repertoires is substantially less than in normal mice. The addition of P nucleotides and N nucleotides at the recombination sites may introduce frameshifts, theoretically generating termination codons in two of every three joining events. These genes cannot produce functional proteins, but such inefficiency is the price that is paid for generating diversity.
J rearrangement brings together multiple germline gene segments that may combine randomly, and different combinations produce different antigen receptors. The maximum possible number of combinations of these gene segments is the product of the numbers of V, J, and (if present) D gene segments at each antigen receptor locus. Therefore, the amount of combinatorial diversity that can be generated at each locus reflects the number of germline V, J, and D gene segments at that locus. After synthesis of antigen receptor proteins, combinatorial diversity is further enhanced by the juxtaposition of two different, randomly generated V regions (i.e., VH and VL in Ig molecules and Vα and Vβ in TCR molecules). Therefore, the total combinatorial diversity is theoretically the product of the combinatorial diversity of each of the two associating chains. The actual degree of combinatorial diversity in the expressed Ig and TCR repertoires in any individual is likely to be considerably less than the theoretical maximum. This is because not all recombinations of gene segments are equally likely to occur, and not all pairings of Ig heavy and light chains or TCR α and β chains may form functional antigen receptors. Importantly, because the numbers of V, D, and J segments in each locus are limited (see Table 8-1), the maximum possible numbers of combinations are on the order of thousands. This is, of course, much less than the actual diversity of antigen receptors in mature lymphocytes. Junctional diversity. The largest contribution to the diversity of antigen receptors is made by the removal or addition of nucleotides at the junctions of the V and D, D and J, or V and J segments at the time these segments are joined. One way in which this can occur is if endonucleases remove nucleotides from the germline sequences at the ends of the recombining gene segments. In addition, new nucleotide sequences, not present in the germline, may be added at junctions. As described earlier, coding segments (e.g., V and J gene segments) that are cleaved by Rag-1 form hairpin loops whose ends are often cleaved asymmetrically by the enzyme Artemis so that one DNA strand is longer than the other (Fig. 8-13). The shorter strand has to be extended with nucleotides complementary to the longer strand before the ligation of the two segments. The short lengths of added nucleotides are called P nucleotides, and their templated addition introduces new sequences at the V-D-J junctions. Another mechanism of junctional diversity is the random addition of up to 20 non-template-encoded nucleotides called N nucleotides (see Fig. 8-13). N region diversification is more common in Ig heavy chains and in TCR β and γ chains than in Ig κ or λ chains. This addition of new nucleotides is mediated by the enzyme terminal deoxynucleotidyl transferase (TdT). In mice rendered deficient in TdT by gene knockout, the diversity of B and T cell repertoires is substantially less than in normal mice. The addition of P nucleotides and N nucleotides at the recombination sites may introduce frameshifts, theoretically generating termination codons in two of every three joining events. These genes cannot produce functional proteins, but such inefficiency is the price that is paid for generating diversity.")
Hasonló előadás







>")









>")
>")
 kialakulása Genetikai, Sejt- és Immunbiológiai Intézet Falus András.>")
