Előadást letölteni
Az előadás letöltése folymat van. Kérjük, várjon

1
A B-sejt differenciáció antigén jelenlétében lezajló folyamatai A B-sejt repertoire és az ellenanyag diverzitás növelése a periférián Alternatív splicing Somatikus hypermutáció Izotípus váltás

2
A membrán asszociált és szekretált Ig-ok közötti váltás Alternativ slicing eredménye.

3
SZOMATIKUS HIPERMUTÁCIÓ

4
CDR1CDR2CDR3 VL Complementary Determining Region = hipervariábilis régió

5
STRUCTURE OF THE VARIABLE REGION Hypervariable (HVR) or complimentarity determining regions (CDR) Framework regions (FR)
 or complimentarity determining regions (CDR) Framework regions (FR)")
6
Somatic hypermutation is targeted to the rearranged gene segments that encode immunoglobulin V regions. AID: activation induced cytidine deaminase AID Cytosine to uracil change….. UNG Uracil-DNA Glycosilase, abasic nucleotide is excised by the nuclease APE1 Base excised and replaced with any nucleotide MUTATION!!!!

7
SZOMATIKUS HIPERMUTÁCIÓ 0. nap Ag 14. nap Ag ELSŐDLEGES I.V. MÁSODLAGOS I.V. Plazmasejt klónok 1234567812345678 9 101 1 12 13 14 15 16 17 18 19 20 21 22 23 24 AFFINITÁS ÉRÉS

8
IZOTÍPUS VÁLTÁS (CLASS SWITCH)
")
9
C Cδ C 3 C 1 Cε2 C 1 C 2 C 4 Cε1 C 2 C Cδ IgM CC CC Ig IZOTÍPUSOK CµIgM Cγ1IgG Cγ2IgG Cγ3IgG Cγ4IgG CαIgA CεIgE

10
C2C2CC C4C4C2C2C1C1C1C1C3C3CC CC Switch régiók Az S 150 [(GAGCT)n(GGGGGT)] ismétlődő szakaszból áll (n=3 – 7) Az izotípus váltás mechanizmusa hasonló a V(D)J rekombinációhoz Az izotípus váltás nem a csontvelőben, hanem az antigénnel való találkozást követően T-sejt segítséggel megy végbe S3S3S1S1S1S1S2S2S4S4SS S2S2 SS A C-régiók előtt a DNS-ben ismétlődő szekvenciákból álló „switch” régiók helyezkednek el. (Kivétel a C régió).
![C2C2CC C4C4C2C2C1C1C1C1C3C3CC CC Switch régiók Az S 150 [(GAGCT)n(GGGGGT)] ismétlődő szakaszból áll (n=3 – 7) Az izotípus váltás mechanizmusa hasonló a V(D)J rekombinációhoz Az izotípus váltás nem a csontvelőben, hanem az antigénnel való találkozást követően T-sejt segítséggel megy végbe S3S3S1S1S1S1S2S2S4S4SS S2S2 SS A C-régiók előtt a DNS-ben ismétlődő szekvenciákból álló „switch régiók helyezkednek el.](http://images.slideplayer.hu/8/2109743/slides/slide_10.jpg "(Kivétel a C régió)..")
11
C2C2CC C4C4C2C2C1C1C1C1C3C3CC CC CC CC C3C3 V 23 D 5 J 4 S3S3 CC CC C3C3 C1C1 S1S1 C1C1 C3C3 C1C1 C3C3 IgG3 termelés IgM IgG3 V 23 D 5 J 4 C1C1 IgA1 termelés IgG3 IgA1 V 23 D 5 J 4 C1C1 IgA1 termelés IgM IgA1 Switch rekombináció Minden rekombinációnál a konstans régiók kivágódnak A génsorrend miatt egy IgE – termelő B sejt nem tud IgM, IgD, IgG1-4 vagy IgA1 termelésre váltani

12
Az ellenanyag izotípus váltás Az immunválasz során egy ellenanyag specificitása (VDJ és VJ) nem változik (az affinitás érés során változhat az affinitása) Az ellenanyagok effektor funkciói az immunválasz során jelentősen változnak Az ellenanyagok képesek a variábilis domén megtartása mellett más konstans régióra váltani, ami más effektor funkciókra teszi képessé az molekulát J régiók C2C2CC C4C4C2C2C1C1C1C1C3C3CC CC Az emberi nehéz lánc gének C régióinak sorrendje, ami meghatározza az izotípus váltás lehetőségeit
 nem változik (az affinitás érés során változhat az affinitása) Az ellenanyagok effektor funkciói az immunválasz során jelentősen változnak Az ellenanyagok képesek a variábilis domén megtartása mellett más konstans régióra váltani, ami más effektor funkciókra teszi képessé az molekulát J régiók C2C2CC C4C4C2C2C1C1C1C1C3C3CC CC Az emberi nehéz lánc gének C régióinak sorrendje, ami meghatározza az izotípus váltás lehetőségeit")
13
Átrendezett DNS IgM-termelő sejt Átrendezett DNS IgE-termelő sejt Első RNA átirat C mRNS nehéz lánc C Cδ C 2 C 4 C C CC Cδ, C 2, C 4 C C Switch regions IZOTÍPUS VÁLTÁS Minden izotípus rekombináció produktív Más szignál szekvenciák és enzimek mint a VDJ átrendeződésnél Antigén stimuláció után Nem véletlenszerű Külső jelek irányítják Hiper IgM szindróma 2. típus Activation Induced Cytidine Deaminase NINCS HIPERMUTÁCIÓ ÉS IZOTÍPUS VÁLTÁS

14
ELLENANYAG ÁLTAL KÖZVETÍTETT EFFEKTOR FUNKCIÓK Neutralizáció – az ellenanyag kötődése gátolja a patogén sejtfelszíni kötődését, belépését a sejtbe vagy szaporodását Opszonizáció – az ellenanyag kötődése elindítja a komplement aktivációt és lehetővé teszi a sejthez való kötődést a komplement (CR1) és Ig (FcR) kötő receptorokon keresztül Az ellenanyag izotípusok eltérő komplement aktiváló és FcR kötő sajátsággal rendelkeznek
 és Ig (FcR) kötő receptorokon keresztül Az ellenanyag izotípusok eltérő komplement aktiváló és FcR kötő sajátsággal rendelkeznek")
15
Az emberi Ig izotípusok effektor funkciók szempontjából különbözők

16
IMMUNKOMPLEX A SZEKRETÁLT ELLENANYAGOK HOZZÁKÖTŐDNEK AZ ANTIGÉNHEZ = IMMUNKOMPLEX KÉPZŐDÉS Makrofág FcRCR KOMPLEMENT AKCTIVÁCIÓ OPSZONIZÁCIÓ FAGOCITÓZIS LEBONTÁS Ig Fc régió Konformáció változás? Asszociáció? KOMPLEMENT AKTIVÁCIÓ – klasszikus útvonal SEJTHEZ KÖTŐDÉS – citofil sajátság IZOTÍPUS FÜGGŐ IgG1 és IgG3 >> IgG2 és IgG4

17
ELLENANYAG KÖZVETÍTETTE EFFEKTOR FUNKCIÓK SPECIFIKUS ELLENANYAG Bakteriális toxin Toxin receptor Neutralizáció Internalizáció Baktérium az interstitiumban Baktérium a plazmában OpszonizációKomplement activáció FagocitózisFagocitózis és lizis KOMPLEMENT

18
AZ ELLENANYAGOK EFFEKTOR FUNKCIÓI PLAZMA SEJT NEUTRALIZÁCIÓ Az ellenanyagok kis hányada GÁTLÁS Baktérium kötődése az epitél sejtekhez Vírus kötődése a receptorhoz Bakteriáláis toxinok kötődése a célsejthez OPSZONIZÁCIÓ Az ellenanyag kötődése fokozza a fagocitózist Fc R Fc R CR1 Komplement C3b KOMPLEMENT AKTIVÁCIÓ Opszonizáció C3b által FAGOCITA SEJTEK BEKEBELEZÉS, LEBONTÁS

19
PLAZMA SEJT ANTIGÉN CITOKINEK B -SEJT A B – SEJT DIFFERENCIÁCIÓT A T-SEJTEK SEGÍTIK IZOTÍPUS VÁLTÁS ÉS AFFINITÁS ÉRÉS CSAK T-SEJT SEGÍTSÉGGEL MEGY VÉGBE HOGYAN LÁTJÁK A T-SEJTEK AZ ANTIGÉNT?

20
T-SEJTEK FEJLŐDÉSE ÉS DIFFERENCIÁCIÓJA

21
A T-sejt prekurzorok a csontvelőből a Timuszba vándorolnak

22
A T-sejt prekurzorok nem expresszálnak T-sejt markereket. Azok a timusz epitéliummal történő interakvió után aktiválódnak

23
A Notch-1receptor és a belőle keletkező transzkripciós faktor szükséges a T-sejt vonal kialakulásához

24
Az αβ és γδ T-sejt vonalak közös (DN) progenitor sejtekből fejlődnek
 progenitor sejtekből fejlődnek")
25
A β-lánc átrendeződése (Ig nehéz lánc homológja) A TCR-átrendeződés—BCR-hez hasonlóan történik Béta lánc átrendeződés hatásfoka kb 80% RAG-1 RAG-2 gén Inaktiválódik Timocita proliferál, CD4 CD8 expresszió
 A TCR-átrendeződés—BCR-hez hasonlóan történik Béta lánc átrendeződés hatásfoka kb 80% RAG-1 RAG-2 gén Inaktiválódik Timocita proliferál, CD4 CD8 expresszió")
26
Az α-lánc átrendeződése (Ig könnyű lánc) A TCR-átrendeződés—BCR-hez hasonlóan történik
 A TCR-átrendeződés—BCR-hez hasonlóan történik")
27
C mIg H mIg L TCR TCR T-SEJT C V Antigén receptor TCR A B- ÉS T-SEJTEK ANTIGÉN FELISMERŐ RECEPTORAI HASONLÓ SZERKEZETŰEK TCR = + A -lánc variábilis régiója V – D – J gének rekombinációjával jön létre – analóg az IgH - lánccal Az -lánc variábilis régiója V – J gének rekombinációjával jön létre – analóg az IgL - lánccal Egy kötőhely Nincs szomatikus mutáció

28
GÉNEK/ KAPCSOLÓDÁS IMMUNOGLOBULIN H / VARIÁBILIS (V) 6570 DIVERZITÁS (D) 270 D (3 keret) ritka- JOINING (J) 65/4 JOINING + P + N 21 50% V GENE PÁROK 3.4x10 6 KAPCSOLÁS ~3x10 7 TELJES ~10 14 SZOMATIKUS HIPERMUTÁCIÓ AZ IMMUNOGLOBULINOK ÉS T-SEJT RECEPTOR GÉNEK SOKFÉLESÉGÉNEK ELMÉLETI LEHETŐSÉGEI T CELL RECEPTOR 52~70 20 gyakran- 1361 21 5.8x10 6 ~2x10 11 10 18 NINCS
 6570 DIVERZITÁS (D) 270 D (3 keret) ritka- JOINING (J) 65/4 JOINING + P + N 21 50% V GENE PÁROK 3.4x10 6 KAPCSOLÁS ~3x10 7 TELJES ~10 14 SZOMATIKUS HIPERMUTÁCIÓ AZ IMMUNOGLOBULINOK ÉS T-SEJT RECEPTOR GÉNEK SOKFÉLESÉGÉNEK ELMÉLETI LEHETŐSÉGEI T CELL RECEPTOR 52~70 20 gyakran x10 6 ~2x NINCS")
29
There are two types of MHC molecule, MHC class I and MHC class II.

30
MHC class I and MHC class II molecules bind to different T-cell co-receptors.

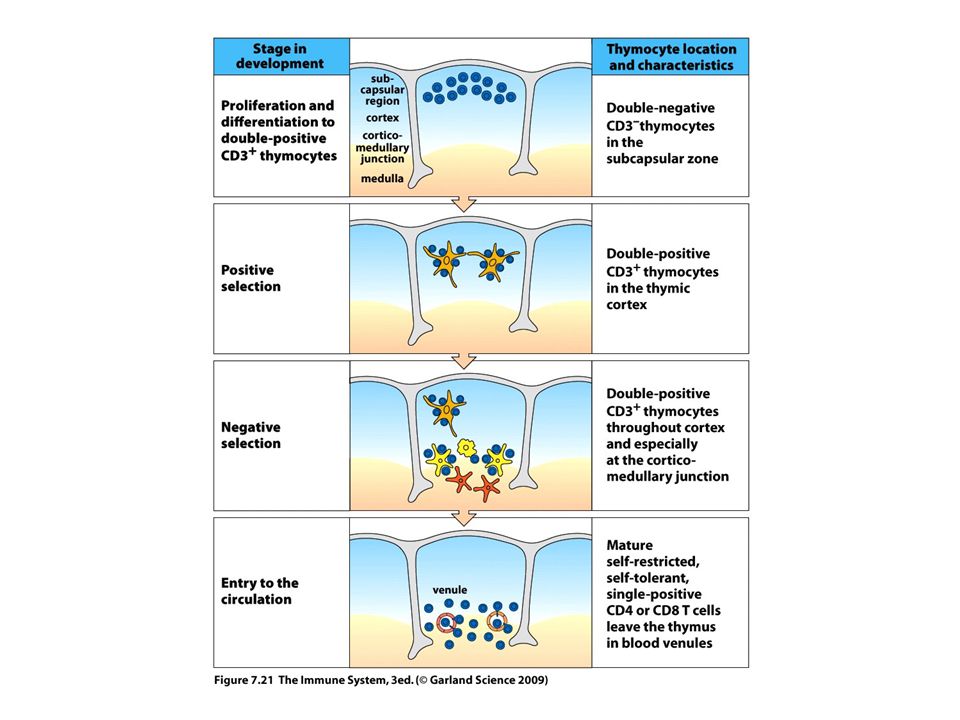
31
A T-sejt repertoire kialakítása. Pozitív és negatív szelekció a Tímuszban

32
Viszonylag kevés, kb 2%-a A TCR-eknek reagál az MHC-val, sok sejt elpusztul Kortex epitélium MHCI MHCII- pozitív Az α-lánc átrendeződés folytatódhat amíg funkcionális Αβ receptor jön létre ami reagál Alkalmas a pozitív szelekcióra A fejlődő T-sejtek pozitív szelekciója a tímuszban

33
A pozitív szelekció szerepe a „single poz”sejtek kialakulásában Kopasz limfocita szindróma Bare lymphocyte syndrome MHCI vagy MHCII hiány CD8+ vagy CD4+ sejtek Hiánya…. Ko-receptorok szerepe A single + T-sejtek kialaku- lásában

34
A fejlődő T-sejtek negatív szelekciója a tímuszban DC Macrophag a velőállo- Mányban. Plussz velőállomány Speciális epitél sejtjei amik egy AIRE-nek nevezett TF-t Expresszálnak---saját szövet- Specifikus fehérjék átírása AIRE mutáció: autoimmun Szindrómát okoz Autoimmune polyendocrinopathy -candidiasis-ectodermal dystrophy AIRE: Autoimmune regulator CENTRÁLIS TOLERANCIA

36
A regulátor T-sejtek különálló fejlődési vonalat képviselnek CD25+ FoxP3+ sejtek FoxP3-hiány: autoimmun betegség IPEX: immune dysregulation polyendocrinopathy, enteropathy, X-linked syndrome

Hasonló előadás





>")











>")
>")

 kialakulása Genetikai, Sejt- és Immunbiológiai Intézet Falus András.>")