Előadást letölteni
Az előadás letöltése folymat van. Kérjük, várjon

1
ANTIGÉNTŐL FÜGGŐ FOLYAMATOK
IZOTÍPUS VÁLTÁS

2
Ig IZOTÍPUSOK Cµ IgM Cγ1 IgG Cγ2 IgG Cγ3 IgG Cγ4 IgG Cα IgA Cε IgE IgM
C Cδ C3 C1 Cε2 C1 C 2 C4 Cε1 C2 C Cδ C Cδ Ig IZOTÍPUSOK Cµ IgM Cγ1 IgG Cγ2 IgG Cγ3 IgG Cγ4 IgG Cα IgA Cε IgE C Cδ C C IgM

3
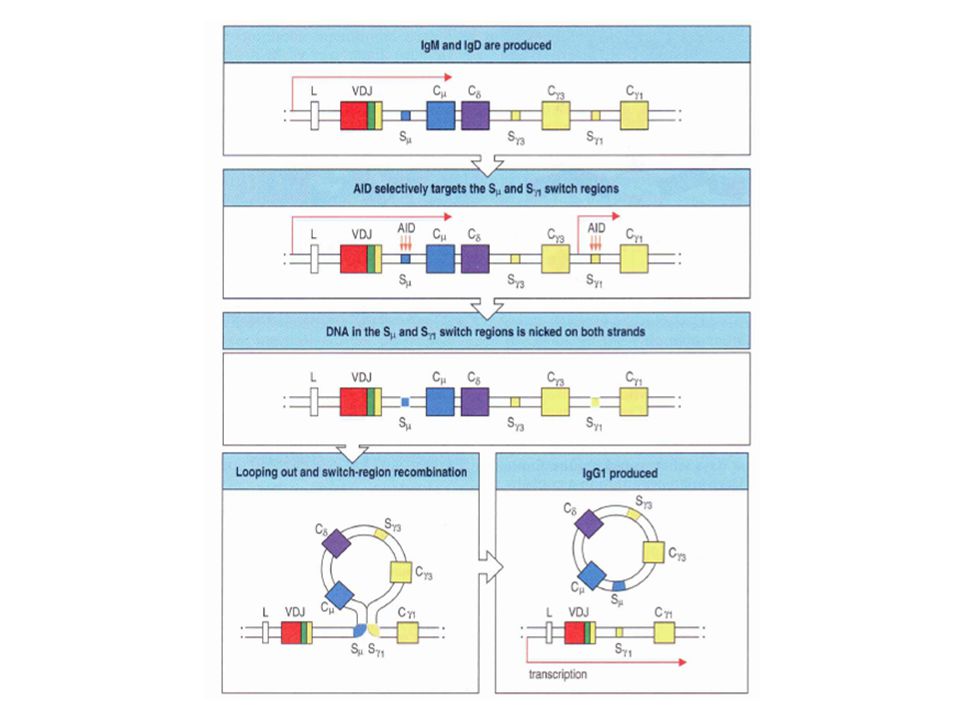
Switch régiók Ca2 Ce Cg4 Cg2 Ca1 Cg1 Cg3 Cd Cm
Sg3 Sg1 Sa1 Sg2 Sg4 Se Sa2 Sm A C-régiók előtt a DNS-ben ismétlődő szekvenciákból álló „switch” régiók helyezkednek el. (Kivétel a Cd régió). Az Sm 150 [(GAGCT)n(GGGGGT)] ismétlődő szakaszból áll (n=3 – 7) Az izotípus váltás mechanizmusa hasonló a V(D)J rekombinációhoz Az izotípus váltás nem a csontvelőben, hanem az antigénnel való találkozást követően T-sejt segítséggel megy végbe
. Az Sm 150 [(GAGCT)n(GGGGGT)] ismétlődő szakaszból áll. (n=3 – 7) Az izotípus váltás mechanizmusa hasonló a V(D)J rekombinációhoz. Az izotípus váltás nem a csontvelőben, hanem az antigénnel való találkozást követően T-sejt segítséggel megy végbe.")
4
Switch rekombináció Ca2 Ce Cg4 Cg2 Ca1 Cg1 Cg3 Cd Cm V23D5J4 V23D5J4
Sg3 Cm Cd Cg3 V23D5J4 Cg1 Sg1 Ca1 Cm Cd Cg3 V23D5J4 Cg3 V23D5J4 Ca1 Cg3 V23D5J4 IgG3 termelés IgM IgG3 V23D5J4 Ca1 IgA1 termelés IgG3 IgA1 V23D5J4 Ca1 IgA1 termelés IgM IgA1 Minden rekombinációnál a konstans régiók kivágódnak A génsorrend miatt egy IgE – termelő B sejt nem tud IgM, IgD, IgG1-4 vagy IgA1 termelésre váltani

5
Az ellenanyag izotípus váltás
Az immunválasz során egy ellenanyag specificitása (VDJ és VJ) nem változik (az affinitás érés során változhat az affinitása) Az ellenanyagok effektor funkciói az immunválasz során jelentősen változnak Az ellenanyagok képesek a variábilis domén megtartása mellett más konstans régióra váltani, ami más effektor funkciókra teszi képessé az molekulát J régiók Ca2 Ce Cg4 Cg2 Ca1 Cg1 Cg3 Cd Cm Az emberi nehéz lánc gének C régióinak sorrendje, ami meghatározza az izotípus váltás lehetőségeit
 nem változik (az affinitás érés során változhat az affinitása) Az ellenanyagok effektor funkciói az immunválasz során jelentősen változnak. Az ellenanyagok képesek a variábilis domén megtartása mellett más konstans régióra váltani, ami más effektor funkciókra teszi képessé az molekulát. J régiók. Ca2. Ce. Cg4. Cg2. Ca1. Cg1. Cg3. Cd. Cm. Az emberi nehéz lánc gének C régióinak sorrendje, ami meghatározza az. izotípus váltás lehetőségeit.")
6
Minden izotípus rekombináció produktív
C Cδ C2 C4 C C Átrendezett DNS IgM-termelő sejt Switch regions C Cδ, C2, C4 IZOTÍPUS VÁLTÁS Minden izotípus rekombináció produktív Más szignál szekvenciák és enzimek mint a VDJ átrendeződésnél Antigén stimuláció után Nem véletlenszerű Külső jelek irányítják C C Átrendezett DNS IgE-termelő sejt Első RNA átirat C mRNS Hiper IgM szindróma 2. típus Activation Induced Cytidine Deaminase (AID) RNS editing enzim NINCS HIPERMUTÁCIÓ ÉS IZOTÍPUS VÁLTÁS nehéz lánc
 RNS editing enzim. NINCS HIPERMUTÁCIÓ ÉS IZOTÍPUS VÁLTÁS. nehéz lánc.")
8
SZOMATIKUS HIPERMUTÁCIÓ

9
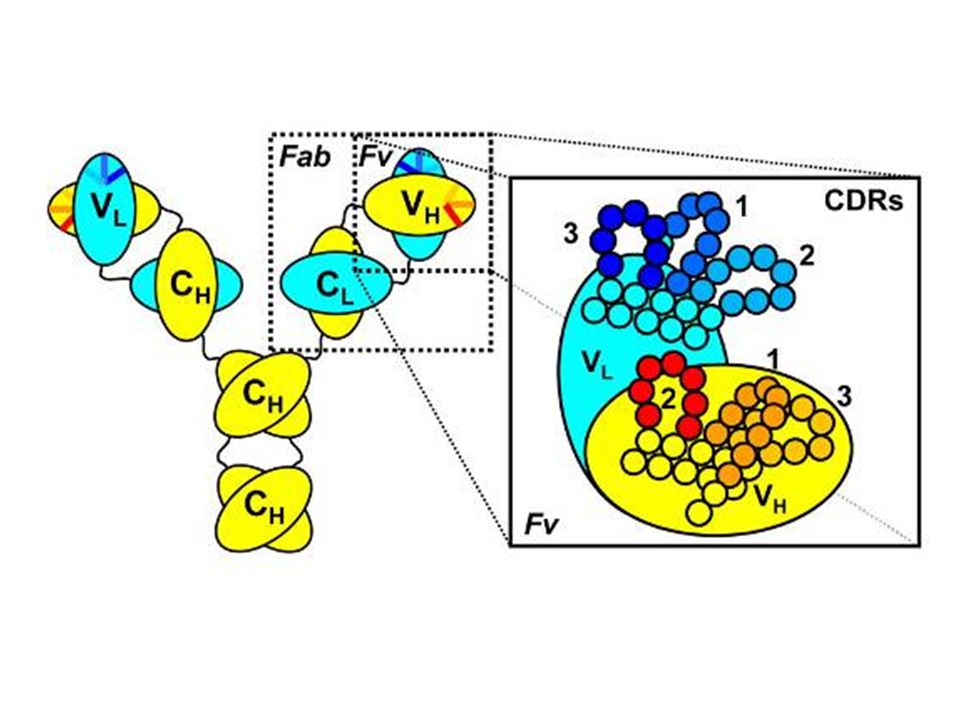
VL CDR1 CDR2 CDR3 Complementary Determining Region = hipervariábilis régió

11
A VARIÁBILIS RÉGIÓ SZERKEZETE
Hipervariábilis (HVR) vagy komplementaritást meghatározó régiók (CDR) HVR3 FR1 FR2 FR3 FR4 HVR1 HVR2 Variabilitás Index 25 75 50 100 Amino savak sorszáma N – C terminális 150 Váz (framework) szakaszok
 vagy komplementaritást meghatározó régiók (CDR) HVR3. FR1. FR2. FR3. FR4. HVR1. HVR2. Variabilitás Index Amino savak sorszáma N – C terminális Váz (framework) szakaszok.")
12
SZOMATIKUS HIPERMUTÁCIÓ
0. nap Ag Plazmasejt klónok 1 2 3 4 5 6 7 8 ELSŐDLEGES I.V. AFFINITÁS ÉRÉS 1 2 3 4 5 6 7 8 14. nap Ag 1 2 3 4 5 6 7 8 MÁSODLAGOS I.V.

13
A szomatikus hipermutáció affinitás éréshez vezet
Clone 1 Clone 2 Clone 3 Clone 4 Clone 5 Clone 6 Clone 7 Clone 8 Clone 9 Clone 10 CDR1 CDR2 CDR3 6. nap CDR1 CDR2 CDR3 8. nap 12. nap 18. nap Hátrányos mutáció Előnyös mutáció Semleges mutáció Kisebb affinitás – Nincs klonális szelekció Nagyobb affinitás – Klonális szelekció Azonos affinitás – Nem hat a klonális delécióra A hipermutációs folyamat aktivált T limfociták közreműködését igényli. A mutáció ‘hot spots’ (CDR régiók) körül halmozódnak. AID és DNS repair enzimek közreműködése eredményezi.
 körül halmozódnak. AID és DNS repair enzimek közreműködése eredményezi.")
14
ELLENANYAG ÁLTAL KÖZVETÍTETT EFFEKTOR FUNKCIÓK
Neutralizáció – az ellenanyag kötődése gátolja a patogén sejtfelszíni kötődését, belépését a sejtbe vagy szaporodását Opszonizáció – az ellenanyag kötődése elindítja a komplement aktivációt és lehetővé teszi a sejthez való kötődést a komplement (CR1) és Ig (FcR) kötő receptorokon keresztül Az ellenanyag izotípusok eltérő komplement aktiváló és FcR kötő sajátsággal rendelkeznek
 és Ig (FcR) kötő receptorokon keresztül. Az ellenanyag izotípusok eltérő komplement aktiváló és FcR kötő sajátsággal rendelkeznek.")
15
A SZEKRETÁLT ELLENANYAGOK HOZZÁKÖTŐDNEK AZ ANTIGÉNHEZ = IMMUNKOMPLEX KÉPZŐDÉS
KOMPLEMENT AKCTIVÁCIÓ OPSZONIZÁCIÓ IMMUNKOMPLEX Makrofág FcR CR Ig Fc régió Konformáció változás? Asszociáció? KOMPLEMENT AKTIVÁCIÓ – klasszikus útvonal SEJTHEZ KÖTŐDÉS – citofil sajátság FAGOCITÓZIS LEBONTÁS IZOTÍPUS FÜGGŐ IgG1 és IgG3 >> IgG2 és IgG4

16
ELLENANYAG KÖZVETÍTETTE EFFEKTOR FUNKCIÓK
SPECIFIKUS ELLENANYAG Bakteriális toxin Baktérium az interstitiumban Baktérium a plazmában Toxin receptor Neutralizáció Opszonizáció Komplement activáció KOMPLEMENT Internalizáció Fagocitózis Fagocitózis és lizis

17
AZ ELLENANYAGOK EFFEKTOR FUNKCIÓI
GÁTLÁS Baktérium kötődése az epitél sejtekhez Vírus kötődése a receptorhoz Bakteriális toxinok kötődése a célsejthez NEUTRALIZÁCIÓ Az ellenanyagok kis hányada PLAZMA SEJT OPSZONIZÁCIÓ Az ellenanyag kötődése fokozza a fagocitózist FcR FcR CR1 Komplement C3b KOMPLEMENT AKTIVÁCIÓ Opszonizáció C3b által FAGOCITA SEJTEK BEKEBELEZÉS, LEBONTÁS

18
A B – SEJT DIFFERENCIÁCIÓT A T-SEJTEK SEGÍTIK
ANTIGÉN CITOKINEK PLAZMA SEJT IZOTÍPUS VÁLTÁS ÉS AFFINITÁS ÉRÉS CSAK T-SEJT SEGÍTSÉGGEL MEGY VÉGBE HOGYAN LÁTJÁK A T-SEJTEK AZ ANTIGÉNT?

19
Sejtfelszíni MHC-peptid komplex
A T LIMFOCITÁK ANTIGÉN FELISMERŐ MŰKÖDÉSÉHEZ PEPTID ANTIGÉNEKRE ÉS MHC MOLEKULÁKAT KIFEJEZŐ SAJÁT SEJTEKRE VAN SZÜKSÉG T Y Sejtfelszíni MHC-peptid komplex T-sejt válasz APC oldott Ag Peptid antigén Natív membrán Ag Sejtfelszíni peptidek Nincs T-sejt válasz

20
AZ ANTIGÉN BEMUTATÁS/PREZENTÁCIÓ
FELTÉTELEI MHC molekulák kifejeződése a sejtfelszínen Vírus antigének intracelluláris szintézise Fehérje antigének felvétele az extracelluláris térből A fehérje antigének denaturációja/feldolgozása a T-sejtek számára Az intracellulárisan feldolgozott fehérje antigének fragmentumai (peptidek) sejtfelszíni bemutatása az MHC molekulák által Az TCR-ral rendelkező T limfociták a fehérje antigénekből származó peptidek felismerésére szakosodtak
 sejtfelszíni bemutatása az MHC molekulák által. Az TCR-ral rendelkező T limfociták a fehérje antigénekből származó peptidek felismerésére szakosodtak.")
21
AZ MHC RÉGIÓ ÁLTAL KÓDOLT
GÉNEK ÉS FEHÉRJÉK

22
EGY VAGY TÖBB Ig DOMÉNT TARTALMAZÓ FEHÉRJÉK V vagy C doménhez hasonló
AZ IMMUNOGLOBULIN SZUPERGÉN CSALÁD TAGJAI EGY VAGY TÖBB Ig DOMÉNT TARTALMAZÓ FEHÉRJÉK V vagy C doménhez hasonló FUNKCIÓ FELISMERÉS Ig, TCR, MHC-I, MHC-II ADHÉZIÓ ICAM-1, ICAM-2, VCAM-1, NCAM KÖTÉS CD4, CD8, CD28, B7, IL-1RI, PDGFR, FcRII, poly-IgR

23
AZ MHC FEHÉRJÉK SZERKEZETE
Peptidkötő hely 1 2 1 1 3 2m 2 2 Transzmembrán Citoplazmatikus 1 2 3 1 2 2m 1 2 MHC II MHC I 3 és 2m Ig szupergén család 2 és 2

24
AZ EMBERI MHC (HLA) TÉRKÉPE A HUMAN GENOME PROJECT ALAPJÁN
3,838,986 bp 224 gén 6 kromoszóma MHC szekvenáló konszorcium Nature 401, 1999

25
AZ MHC MOLEKULÁK BIOLÓGIAI FUNKCIÓJA

26
AZ MHC KORLÁTOZÁS JELENSÉGE
A TCR MHC + peptid komplexeket ismer fel Egy adott TCR egy adott MHC – peptid komplex felismerésére képes Ugyanazt a peptidet egy másik MHC molekulához kötődve ugyanaz a TCR nem ismeri fel (más TCR felismerheti) Ugyanazt az MHC molekulát egy másik peptiddel együtt ugyanaz a TCR nem ismeri fel (más TCR felismerheti)
 Ugyanazt az MHC molekulát egy másik peptiddel együtt ugyanaz a TCR nem ismeri fel (más TCR felismerheti)")
27
AZ I TÍPUSÚ MHC MOLEKULA TÉRSZERKEZETE
PEPTID PEPTID 1 3 2 2m MINDEN MAGVAS SEJTEN KIFEJEZŐDIK

28
AZ II TÍPUSÚ MHC MOLEKULA TÉRSZERKEZETE
PEPTIDE PEPTID 2 1 2 1 A HIVATÁSOS ANTIGÉN PREZENTÁLÓ SEJTEKEN JELENIK MEG MÁS SEJTEKEN IS INDUKÁLHATÓ (endotél, mikroglia, asztocita)
")
29
AZ I TÍPUSÚ MHC MOLEKULA PEPTID KÖTŐ HELYE

30
AZ II TÍPUSÚ MHC MOLEKULA PEPTID KÖTŐ HELYE

31
A PEPTIDKÖTŐ HELY GEOMETRIÁJA
b2m a-lánc Peptid a-lánc b-lánc Peptid Az MHC-I molekula 8-10 aminosav hosszúságú peptideket köt Az MHC-II molekula >13 aminosav hosszúságú peptideket köt

32
A PEPTIDKÖTŐ HELY SZERKEZETE
A „törzs” régió aminosav oldalláncai egyenletesen elosztott zsebekbe illeszkednek P2 és P9 nagy hidrofób zsebbe illeszkednek

33
A peptidek leoldhatók az MHC molekulákról
Stabil MHC-peptid komplexek izolálása Peptid szeparálás és szekvenálás Savval eluált peptidek „üres” MHC molekulák

34
Az MHC molekulákról leoldott peptidek eltérő szekvenciákkal rendelkeznek de közös motívumokat tartalmaznak Egy adott MHC I molekulához kötődő peptidek állandó aminosav mintázatot mutatnak A közös szekvencia részlet a MOTIF R T Y Q L V N C P E I Y S F H A sok peptidre jellemző közös aminosavak illeszkednek az MHC molekula szerkezetéhez HORGONYZÓ AMINOSAVAK A V T Y K Q L P S A Y I K R G Y V Q L Nem azonosak de hasonlók Y & F aromás V, L & I hidrofób S I F N E K L A P G Y N L A horgonyzó aminosavak oldalláncai a zsebekbe illeszkednek Az eltérő MHC molekulák különböző konzervált aminosav mintázattal rendelkező peptideket képesek megkötni

35
Az allélikus polimorfizmus a peptid kötő helyre koncentrálódik
Class I Class II (HLA-DR) 1 3 2 2m 2 1 2 1 Az MHC polimorfizmus befolyásolja a peptid kötő képességet Az allelikus variránsok 20 aminosavban is eltérhetnek
 1. 3. 2. 2m. 2. 1. 2. 1. Az MHC polimorfizmus befolyásolja a peptid kötő képességet. Az allelikus variránsok 20 aminosavban is eltérhetnek.")
36
A legtöbb polimorfizmus pont mutáció
következménye DPB1*01011 TAC GCG CGC TTC GAC AGC GAC GTG GGG GAG TTC CGG GCG GTG ACG GAG CTG GGG CGG CCT GCT GCG GAG TAC TGG AAC AGC CAG AAG GAC ATC CTG GAG GAG DPB1* A DPB1* T- -T A- -A DPB1* T- -T AC -A DPB1*0202 CT- -T AG DPB1* T- -T A- -A- --C C DPB1* T DPB1* T- -T A- -A DPB1*0501 CT- -T AG DPB1* T- -T A- -A- --C C DPB1* T- -T A- -A DPB1* T- -T A- -A- --C DPB1* T- -T A- -A DPB1* A C DPB1* A C DPB1* DPB1* T- -T A- -A- --C C DPB1* A C DPB1* T- -T A- -A DPB1* T- -T A- -A- --C DPB1* T- -T A- -A DPB1* T- -T AG DPB1* T- -T A- -A- --C C DPB1* T- -T A- -A- --C C DPB1*2101 CT- -T AG DPB1*2201 CT- -T AG DPB1* T- -T DPB1* T AG DPB1* T- -T A- -A C DPB1* A DPB1* 30 HLA-DPb allél szekvenciája a 204 és 290 nukleotidok (35-68 aminosavak) közötti szakaszon A polimorf nukleotidok a peptidkötő helyhez tartoznak Y-F A-V Silent A-D A-E E-A I-L
 közötti szakaszon. A polimorf nukleotidok a peptidkötő. helyhez tartoznak. Y-F. A-V. Silent. A-D. A-E. E-A. I-L.")
37
AZ MHC GÉNEK ELHELYEZKEDÉSE
6 kromoszóma rövid karja MHC 15 kromoszóma 2m Nem- klasszikus MHC gének E, G, F Klasszikus MHC gének POLIMORF HLA – Human Leukocyte Antigen rendszer HLA –A, B, C I osztály MINDEN MAGVAS SEJTEN HLA – DR, DP, DQ II osztály HIVATÁSOS ANTIGÉN PREZENTÁLÓ SEJTEKEN III osztály

38
AZ MHC MOLEKULÁK SZÖVETI MEGOSZLÁSA
Szövet MHC I MHC II T sejt /- B sejt Makrofág Dendritikus sejt Tímusz epitél sejtek Neutrofilek Hepatociták Vese Agy Eritrocita A sejt aktiváció befolyásolja az MHC expressziót A szöveti kifejeződés jól tükrözi az MHC molekulák funkcióit Az I osztályú MHC molekulák a vírus és tumor ellenes immunitásban fontosak A II osztályú MHC molekulák az immunrendszer sejtjeinek aktiválásában, együttműködésében játszanak szerepet

39
AZ I ÉS II OSZTÁLYÚ MHC GÉNEK ÖRÖKLŐDÉSE HUMAN LEUKOCYTE ANTIGEN
HLA I osztály II osztály EVERY CELL α1β1 α2β2 PROFESSIONAL APC

40
AZ MHC MOLEKULÁK POLIMORFIZMUSA AZ EMBERI POPULÁCIÓBAN
? FELTÉTELEZÉS • Minden egyed 6 I típusú MHC molekulát fejez ki • Az összes MHC allotípus elvileg véletlenszerűen oszlik el a populációban • Az különböző allél bármely másik alléllal együttesen fordulhat elő ~6 x 1015 egyedi kombináció Csak az egypetéjű ikrek HLA lókuszai megegyezőek Az emberi populáció nagyon kevert (outbred) Az MHC genetika nagyon összetett POLIGENITÁS, POLIMORFIZMUS
 Az MHC genetika nagyon összetett. POLIGENITÁS, POLIMORFIZMUS.")
41
I osztály II osztály ~6 x 1015 egyedi kombináció
AZ EMBERI MHC (HLA) POLIMORFIZMUSA AZ EMBERI POPULÁCIÓBAN 872 I osztály 1652 allél 506 15.18 28.65 13.38 4.46 0.02 5.72 18.88 8.44 9.92 1.88 4.48 24.63 2.64 1.76 0.01 CAU AFR ASI Frequencia (%) HLA-A1 HLA- A2 HLA- A3 HLA- A28 HLA- A36 Allél csoportok 274 A polimorfizmus (allélek) száma A B C II osztály 688 allél 466 114 66 15 25 2 A valóságban az MHC allélek NEM véletlenszerűen oszlanak el a populációban Az allélek a fajták és a vonalak között haplotípusokban szegregálódnak a b DR DP DQ
 POLIMORFIZMUSA AZ EMBERI POPULÁCIÓBAN I osztály allél CAU. AFR. ASI. Frequencia (%) HLA-A1. HLA- A2. HLA- A3. HLA- A28. HLA- A36. Allél csoportok A polimorfizmus (allélek) száma. A. B. C. II osztály. 688 allél A valóságban az MHC allélek NEM véletlenszerűen oszlanak el a populációban. Az allélek a fajták és a vonalak között haplotípusokban szegregálódnak. a. b. DR. DP. DQ.")
42
A klasszikus MHC gének kapcsoltan, haplotípusokban öröklődnek
DP DQ DR Gyerekek DP-1,8 DQ-3,6 DR-5,4 B-7,2 C-9,8 A-11,10 DP-1,9 DQ-3,7 DR-5,5 B-7,3 C-9,1 A-11,9 DP-2,8 DQ-4,6 DR-6,4 B-8,2 C-10,8 A-12,10 DP-2,9 DQ-4,7 DR-6,5 B-8,3 C-10,10 A-12,9 B C A DP DQ DR X Szülők B C A DP DQ DR DP-1,2 DQ-3,4 DR-5,6 B-7,8 C-9,10 A-11,12 DP-9,8 DQ-7,6 DR-5,4 B-3,2 C-1,8 A-9,10 B C A DP DQ DR B C A DP DQ DR

43
Bemutatják a sejt belső környezetét
AZ MHC MOLEKULÁK SAJÁT VAGY ANTIGÉN EREDETŰ PEPTIDEKET KÖTVE JELENNEK MEG A SEJTFELSZÍNEN Vese epitél sejt Bemutatják a sejt belső környezetét B-sejt, makrofág, dendritikus sejt Bemutatják a sejt belső és külső környezetét Máj sejt I. típusú MHC A citoszólból és a sejtmagból származó adott méretű peptidek II. típusú MHC Membrán fehérjékből és az MHC molekulákból (70%) származó peptidek
 származó peptidek.")
44
AZ MHC ALLOTÍPUSOK KOMBINÁCIÓJA A POPULÁCIÓBAN ÉS AZ EGYEDBEN
* * * * * * 2. EGYED 1. EGYED

45
A FERTŐZÉSEK KIMENETELE EGY ÉS TÖBB POLIMORF MHC GÉN ESETÉN
Példa: Ha csak egyféle MHC molekula (MHC X) lenne a populációban A patogén kikerüli az MHC X általi felismerést MHC XX Többféle MHC-Gén v A populáció védett v A populációt a kihalás fenyegetné V – vírus fertőzés által okozott kár
 lenne a populációban. A patogén kikerüli az MHC X általi felismerést. MHC. XX. Többféle MHC-Gén. v. A populáció védett. v. A populációt a kihalás fenyegetné. V – vírus fertőzés által okozott kár.")
46
Mi az előnye az MHC típusok sokféleségének?
• A patogén mikroorganizmusok osztódása lényegesen gyorsabb, mint az emberi reprodukció • Adott idő alatt a patogén gének sokkal gyakrabban mutálódnak, mint az emberi gének és ezáltal gyakran kikerülhetik az MHC gének változásait • Az MHC típusok száma korlátozott A patogének flexibilitásával szemben • A populációban minden MHC típus sok variánst hoz létre • Ezek a variánsok nem feltétlenül nyújtanak védelmet minden egyed számára, de védik a populációt a kihalástól

47
Az MHC típusok és variánsok molekuláris magyarázata
POLIGENITÁS Számos MHC I és II gén kódol különböző típusú MHC molekulákat eltérő peptid-kötő specificitással POLIMORFIZMUS >1% variábilitás egyetlen genetikai lókuszon a populációban az MHC-k a ma ismert leginkább polimorf gének Az MHC molekulák típusai és variábilitásai nem változnak az egyed élete folyamán Az MHC molekulák diverzitása populáció szinten érvényesül, szöges ellentétben a T és B sejt antigén receptorok diverzitásával, melyek egy egyeden belül érvényesülnek

48
AZ MHC FUNKCIÓI KLASSZIKUS MHC GÉN TERMÉKEK NEM KLASSZIKUS MHC GÉNEK
Saját fehérjékből származó peptidek prezentálása – immunológiai saját folyamatos megjelenítése Az immunológiai saját meghatározása Saját MHC + saját peptid – egyedekként változó MHC és saját peptid Antigénből származó peptidek prezentálása – idegen/megváltozott saját felismerése Az immunológiailag idegen meghatározása Saját MHC + idegen peptid – egyedekként változó MHC és idegen peptid Allogén válasz idegen MHC-val szembeni válasz (transzplantáció) Az MHC által korlátozott T-sejt felismerés következménye A T-limfociták differenciációja és szelekciója a tímuszban A T-limfociták életben tartása a periférián Az NK sejt felismerés célpontja NEM KLASSZIKUS MHC GÉNEK Specializált funkciók A KLASSZIKUS MHC GÉNEKKEL SZERKEZETI ROKONSÁGOT MUTATÓ FEHÉRJÉK
 Az MHC által korlátozott T-sejt felismerés következménye. A T-limfociták differenciációja és szelekciója a tímuszban. A T-limfociták életben tartása a periférián. Az NK sejt felismerés célpontja. NEM KLASSZIKUS MHC GÉNEK. Specializált funkciók. A KLASSZIKUS MHC GÉNEKKEL SZERKEZETI ROKONSÁGOT MUTATÓ FEHÉRJÉK.")
49
ANTIGÉN PREZENTÁCIÓ

50
A T sejtek antigén felismerő receptorainak becsült száma
1,000,000,000,000,000 ( ) Ennyi különböző antigén eredetű peptid Hogy képes 6 nem variábilis MHC molekula 1,000,000,000,000,000 különböző peptidet kellő erőséggel megkötni? Hol történik a peptid kötés?
 Ennyi különböző antigén eredetű peptid. Hogy képes 6 nem variábilis MHC molekula. 1,000,000,000,000,000 különböző peptidet. kellő erőséggel megkötni Hol történik a peptid kötés")
51
Olyan kötőhely, amely elég rugalmas bármely peptid megkötésére?
Flexibilis kötőhely? Olyan kötőhely, amely elég rugalmas bármely peptid megkötésére? A sejtfelszínen az ilyen kötőhely nem lenne képes • elég nagy erősséggel kialakítani az MHC – peptid – TCR hármas komplexet • megakadályozni a peptidek lecserélődését extracelluláris peptidek által

52
Flexibilis kötőhely? Laza, rugalmas Zárt
A kötőhely kialakulásának kezdeti, intracelluláris szakaszában a peptid irányítja az MHC molekula térszerkezetét Laza, rugalmas Zárt Egy adott MHC molekula számára lehetővé teszi, hogy • sok különböző peptiddel lépjen kapcsolatba • a peptidet a sejtfelszínen nagy affinitással kösse • stabil komplexeket képezzen a sejtfelszínen • csak olyan molekulákat szállítson a sejtfelszínre, amelyek intracellulárisan peptidet kötöttek

53
A T sejtek antigén felismerő receptorainak becsült száma
1,000,000,000,000,000 ( ) Hogy képes 6 nem variábilis molekula 1,000,000,000,000,000 különböző peptidet megkötni? MHC molekulák • Amíg a peptid kötés nem történik meg, flexibilis konformációt vesznek fel • A peptidkötést konformáció változás kíséri, ami növeli a komplex stabilitását • A peptid „befogására” kevés horgonyzó aminosav szolgál - a horgonyzó aminosavak közt különböző szekvenciák lehetnek - különböző hosszúságú peptidek kötődhetnek
 Hogy képes 6 nem variábilis molekula. 1,000,000,000,000,000 különböző peptidet megkötni MHC molekulák. • Amíg a peptid kötés nem történik meg, flexibilis konformációt vesznek fel. • A peptidkötést konformáció változás kíséri, ami növeli a komplex stabilitását. • A peptid „befogására kevés horgonyzó aminosav szolgál. - a horgonyzó aminosavak közt különböző szekvenciák lehetnek. - különböző hosszúságú peptidek kötődhetnek.")
54
IFN általi koordinált kifejeződés
AZ ENDOGÉN ANTIGÉN BEMUTATÁSI ÚT Tc-sejt MHC-I + saját peptid MHC-I + Ag peptid α-lánc+β2m MHC+peptid NYITOTT ZÁRT MHC-I, LMP2/7, TAP IFN általi koordinált kifejeződés α-lánc TAP1/2 gp96 calnexin FEHÉRJE SAJÁT ANTIGÉN Proteaszóma LMP2/LMP7

55
AZ EXOGÉN ANTIGÉN BEMUTATÁSI ÚT
Th-sejt MHC-II + Ag peptid MHC-II + saját peptid ZÁRT NYITOTT Ii+αβ CLIP DMA/B INVARIÁNS LÁNC (Ii) Chaperon – konformáció Peptidkötőhely gátlása Szállító/visszatartó molekula DMA/DMB 1. A peptidet befogadó konformáció fenntartása 2. A CLIP és az exogén fehérjékből származó peptidek lecserélése
 Chaperon – konformáció. Peptidkötőhely gátlása. Szállító/visszatartó molekula. DMA/DMB. 1. A peptidet befogadó konformáció fenntartása. 2. A CLIP és az exogén fehérjékből származó peptidek lecserélése.")
56
ANTIGEN ÁTALAKÍTÁS ÉS BEMUTATÁS
MHC I MHC II Kötött peptid forrás saját vagy idegen fehérjék méret 8-10 aminosav 13-25 aminosav heterogenitás korlátozott átfedő peptidek természetes citoplazmatikus és magi fehérjék ~70% MHC eredetű, membrán- és extra- celluláris fehérjék Peptid képződés helye citoplazma vezikulumok endo/lizoszóma enzimek proteaszóma LMP-2, LMP-7 szabályozó egységek savas proteázok katepszinek transzport TAP – peptid méret C-terminális cytoplasm ER nincs MHC transzport Ii - irányít, visszatart ER vezikuláris rendszer speciális vezikulum CIIV MHC - peptide kölcsönhatás ER speciális vezikulum, CIIV chaperonok calnexin, tapazin, calretuculin Ii, DMA/B MHC - peptid komplexek a sejtfelszínen stabil komplexek a sejt belső környezetét tükrözik kevés nem stabil üres molekula nincs peptid disszociáció, csere stabil complexek a sejt belső/külső környezetét tükrözik kevés recirkuláló és CLIP-peptidet kötő molekula kismértékű peptid csere

57
AZ MHC GÉNEK ELHELYEZKEDÉSE
6 kromoszóma rövid karja MHC 15 kromoszóma 2m Nem- klasszikus MHC gének E, G, F Klasszikus MHC gének POLIMORF HLA – Human Leukocyte Antigen rendszer HLA –A, B, C I osztály MINDEN MAGVAS SEJTEN HLA – DR, DP, DQ II osztály HIVATÁSOS ANTIGÉN PREZENTÁLÓ SEJTEKEN III osztály

58
AZ MHC EGYÉB GÉNJEI (nem klasszikus) nem polimorf
Ib MHC gének I típusú, 2 mikroglobulinnal asszociált MHC szerű molekulák Korlátozott szöveti kifejeződés HLA-G trofoblaszt, kapcsolódik a CD94 NK-sejt receptorhoz, gátolja a magzat és tumorok NK-sejt általi pusztítását HLA-E bizonyos sejtek membránján, HLA-A, B, C gének szignál szekvenciáját köti, kapcsolódik a CD94 NK-sejt receptorhoz HLA-F magzati máj, eozinofil felszín, ismeretlen funkció MHC II régió Az antigén feldolgozásban szereplő géneket kódolnak HLA-DM/, HLA-DO hivatásos APC-ben Proteaszóma komponensek (LMP-2 és 7), peptid transzporterek (TAP-1 és 2) Sok pseudogén MHC III régió Komplement fehérjék kódolása C4A és C4B, C2 és B FAKTOR TUMOR NEKRÓZIS FAKTOR- / Immunológiailag irreleváns gének 21-hidroxiláz, RNA helikáz, kazein kináz hősokk fehérje 70, szialidáz
, peptid transzporterek (TAP-1 és 2) Sok pseudogén. MHC III régió. Komplement fehérjék kódolása C4A és C4B, C2 és B FAKTOR. TUMOR NEKRÓZIS FAKTOR- / Immunológiailag irreleváns gének. 21-hidroxiláz, RNA helikáz, kazein kináz. hősokk fehérje 70, szialidáz.")
Hasonló előadás






>")









>")
>")
 kialakulása Genetikai, Sejt- és Immunbiológiai Intézet Falus András.>")
