Előadást letölteni
Az előadás letöltése folymat van. Kérjük, várjon

1
Barcelona

2
A szegregáció törvénye
Mendel első törvénye: A szegregáció törvénye Monohibrid keresztezés A gaméták tisztaságának a törvénye Mendel második törvénye: Dihibrid keresztezés A független kombináció törvénye

3
Genetikai arányok számolási módjai
„n” szegregáló allélpár beltenyésztése során az utódokban : a különböző genotípusok száma 3n a különböző fenotípusok száma 2n A Punnett tábla nagyon szemléletes módszer az F2 nemzedék geno- és fenotípus arányainak leírására, de kettőnél több faktor esetén kényelmetlen.

4
AaBb beltenyésztésével szemléltetve:
Az „ág” diagram kényelmes módszer az utód geno- és fenotípusok arányainak leírására. AaBb beltenyésztésével szemléltetve: Beltenyésztés utódainak genotípusai Beltenyésztés utódainak fenotípusai Gaméták ¼ BB ¾ B- ½ B ¼ AA ½ Bb ¾ A- ½ A ¼ bb ½ b ½ Aa ¼ aa ½ a

5
AA bb Cc DD ee Ff genotípus?
Nagyszámú gén esetén egy bizonyos genotípus létrejöttének valószínűségét egyszerűen számíthatjuk ki: Pl.: Aa Bb Cc Dd Ee Ff x Aa Bb Cc Dd Ee Ff keresztezés utódai között milyen valószínűséggel fordul elő AA bb Cc DD ee Ff genotípus? Az egyes kívánt allél párok létrejöttének valószínűségei: AA ¼ bb ¼ Cc ½ DD ¼ ee ¼ Ff ½ A keresett ganotípus előfordulásának valószínűsége az allél párok valószínűségének szorzata: p(AA bb Cc DD ee Ff) = ¼ x ¼ x ½ x ¼ x ¼ x ½ = 1/1024.
 = ¼ x ¼ x ½ x ¼ x ¼ x ½ = 1/1024.")
6
A mendeli arányoknak nevezzük a következőket:
Mendel munkásságának újrafelfedezése (1900) után nagyon sok különböző élőlényen ellenőrizték és találták érvényesnek a törvényszerűségeket. A mendeli arányoknak nevezzük a következőket: 3 : 1 9 : 3 : 3 : 1 1 : 1 : 1: 1
 után nagyon sok különböző élőlényen ellenőrizték és találták érvényesnek a törvényszerűségeket. A mendeli arányoknak nevezzük a következőket: 3 : 1. 9 : 3 : 3 : 1. 1 : 1 : 1: 1.")
7
Az ember egyszerű mendeli genetikája
A mendeli genetika alkalmazása az ember tulajdonságainak öröklésére különleges módszereket igényel. Az ebben a rendszerben alkalmas módszer a pedigré analízis vagy törzsfa szerkesztés. A törzsfa szerkesztéshez egységes jelképrendszert használnak. férfi nő házasság gyerekek

8
A családfa szerkesztésben használt szimbólumok

9
Normális fenotípus változatok
Minden populáció mutat változékonyságot. Változatok: folyamatos, átfedő eloszlás magasság, súly, stb. mennyiségi genetika tárgyai nem folyamatos, nem átfedő eloszlás - mendeli genetika tárgyai - embernél: barna és kék szem sötét és szőke haj sötét és vörös haj állgödör és állgödör hiánya lenőtt és szabad fülcimpa

10
Polimorfizmus: Dimorfizmus:
két vagy több normális fenotípus előfordulása a populációban A polimorfizmus nagyon gyakori a természetes populációkban és DNS szintjén is tanulmányozható. Dimorfizmus: csak két fenotípus feniltiokarbamid(PTC) keserű ízének vagy ízetlennek érzékelése a PTC keserű izének érzése mint domináns allél öröklődik
 keserű ízének vagy ízetlennek érzékelése. a PTC keserű izének érzése mint domináns allél öröklődik.")
11
Orvosi genetika Recesszíven öröklődő allélok és fenotípusaik (rendellenességek) : fenolketonuria (PKU), a beteg pp, az egészséges ember PP vagy Pp albinizmus, cisztikus fibrózis, izomsorvadás. recesszív rendellenességek örökletességére jellemző: a terheltség egészséges szülők gyermekeiben bukkan fel, nőkben és férfiakban egyaránt jelentkezik a pedigréből a genotípusok is kikövetkeztethetők több generációt mutató pedigrékben a terhelt fenotípust mutató egyedek, testvérek között, csoportosan jelennek meg, nem fordulnak elő minden generációban a recesszív betegségek megjelenése gyakoribb a rokonházasságokban. .

12
Recesszív emberi rendellenesség öröklődését mutató családfa
A kis utódszámok miatt nem várunk pontos mendeli arányokat. nem terhelt szülők utódaiban jelenik meg Ha az utódok mindkét nemében azonos gyakoriságú, az arra utal, hogy nem kötött nemhez.

13
Orvosi genetika Dominánsan öröklődő allélok és fenotípusaik (rendellenességek) : Az öröklésmenetben minden nemzedékben megjelenik a terheltség, mindkét nemben. A dominánsan öröklődő rendellenességek esetén a normális, egészséges allél a recesszív

14
A pseudo-achondropasia fenotípusa
Az öröklésmenetben minden nemzedékben megjelenik a terheltség, mindkét nemben. A dominánsan öröklődő rendellenességek esetén a normális, egészséges allél a recesszív akondroplázia, a törpeség egyik formája Az egészséges egyedek genotípusa dd, a betegek Dd vagy DD.

15
Polidaktilia Brachydactylia

16
Mendel munkája a genetikai analízis prototípusa.
Megmutatja, hogyan lehetséges biológiai folyamatokat tanulmányozni genetikai markerek használatával. Megmutatja, hogyan tisztázható a gének funkciója változataik tanulmányozása segítségével Messze ható következményei vannak a mezőgazdaságra és a gyógyításra.

17
Törvényszerűségek A gének létezésére öröklődő változatok közötti keresztezések utódai között megfigyelhető bizonyos arányból következtethetünk. Egy bizonyos jelleg különbségét gyakran egyetlen gén különbség okozza. Magasabb rendű szervezetek sejtjeiben minden gén két példányban van meg. A szaporítósejtek kialakulása során a gén-pár minden tagja külön ivarsejtbe kerül. A szaporító sejtek kialakulása során különböző gének viselkedése gyakran egymástól független.

18
Az öröklődés kromoszóma elmélete
A kromoszóma elmélet megfogalmazását (1902) W. Sutton és T. Boveri-nak tulajdonítják. A gének és kromoszómák párhuzamos viselkedése vezetett a feltételezéshez, hogy a gének a kromoszómákon helyezkednek el.
 W. Sutton és T. Boveri-nak tulajdonítják. A gének és kromoszómák párhuzamos viselkedése vezetett a feltételezéshez, hogy a gének a kromoszómákon helyezkednek el.")
19
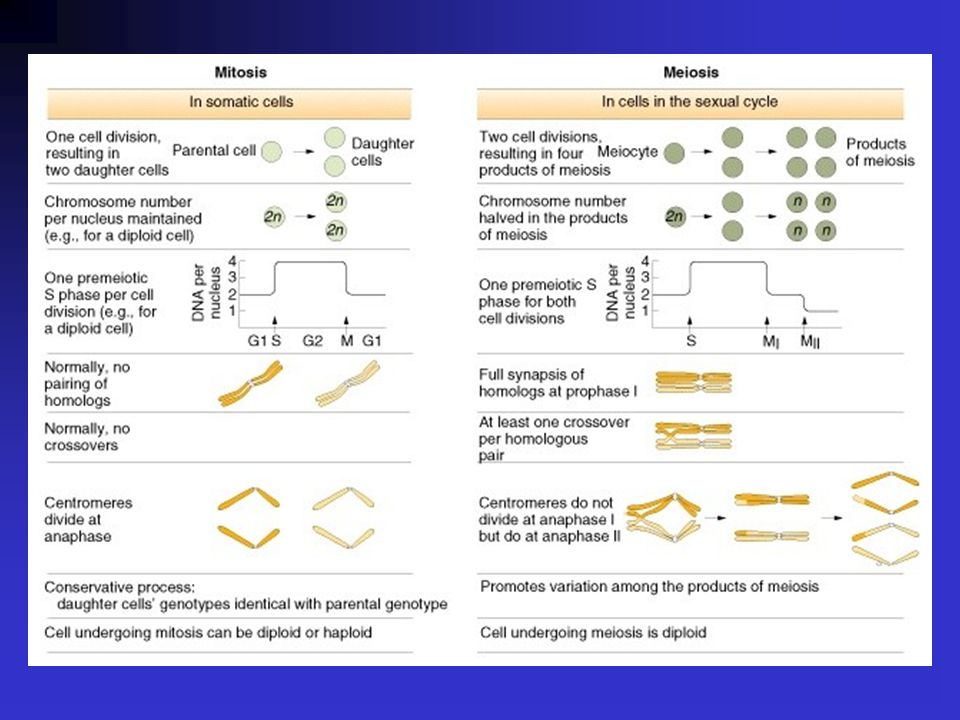
Párhuzamok a mendeli gének és a kromoszómák viselkedése között a meiózisban.

20
Az Öröklődés egyszeriben biztosítja az állandósgot és változatosságot:
a konzervatív mitózis a “liberális” meiózis (rekombináció) Sutton és Boveri elméletének korabeli kritikája: Változatlanok maradnak-e a kromoszómák az interfázis során? Biztos-e hogy a kromoszómák homológ páronként és nem véletlenszerűen párosodnak? Biztos-e, hogy a kromoszómák különböző géneket tartamaznak?
 Sutton és Boveri elméletének korabeli kritikája: Változatlanok maradnak-e a kromoszómák az interfázis során Biztos-e hogy a kromoszómák homológ páronként és nem véletlenszerűen párosodnak Biztos-e, hogy a kromoszómák különböző géneket tartamaznak")
21
A szegregáció törvénye A gaméták tisztaságának a törvénye
Az elméletet alátámasztó korabeli megfigyelések: E. Carothers (1913): sáska tesztiszben egy heteromorf kromoszóma párt tanulmányozott. Anafázisban megfigyelte a heteromorf pár eloszlását a pólusokon, és azt egyenlőnek találta. A heteromorf és a páratlan kromoszómák azonos gyakoriságú mintázata kerülnek a gamétákba. Mendel első törvénye: A szegregáció törvénye A gaméták tisztaságának a törvénye
: sáska tesztiszben egy heteromorf kromoszóma párt tanulmányozott. Anafázisban megfigyelte a heteromorf pár eloszlását a pólusokon, és azt egyenlőnek találta. A heteromorf és a páratlan kromoszómák azonos gyakoriságú mintázata kerülnek a gamétákba. Mendel első törvénye: A szegregáció törvénye. A gaméták tisztaságának a törvénye.")
22
Az egyes kromoszómák információ tartalma különböző
Az elméletet alátámasztó korabeli megfigyelések: Az egyes kromoszómák információ tartalma különböző A. Blackeslee (1922) 12 vonalat állított elő Datura növényből (12 kromoszóma párja van), amelyek egy-egy számfeletti kromoszómát tartalmaztak (triszomok). Minden törzs fenotípusa más-más. Ebből arra következtetett, hogy a kromoszómák különböző géneket tartamaznak. Datura növények termései
 12 vonalat állított elő Datura növényből. (12 kromoszóma párja van), amelyek egy-egy számfeletti. kromoszómát tartalmaztak (triszomok). Minden törzs fenotípusa más-más. Ebből arra következtetett, hogy a kromoszómák különböző géneket tartamaznak. Datura növények termései.")
23
Reciprok keresztezések eltérő F1 fenotípusok.
Közvetett bizonyítékok ! Reciprok keresztezések eltérő F1 fenotípusok. ? Th. H. Morgan (1909) nemhez – kötött öröklődés
 nemhez – kötött öröklődés.")
24
N. Stevens (1905) A heteromorf kromoszóma-pár (X és Y) szegregációja a Tenebrio hímek meiózisa során
 A heteromorf kromoszóma-pár (X és Y) szegregációja a Tenebrio hímek meiózisa során.")
25
Két rovarfaj hímjeinek és nőstényeinek kromoszóma szerelvényei.
N. Stevens (1905) Két rovarfaj hímjeinek és nőstényeinek kromoszóma szerelvényei.
 Két rovarfaj hímjeinek és nőstényeinek kromoszóma szerelvényei.")
26
A muslicák nemét az X és Y kromoszómák határozzák meg.
Fehér és piros-szemű Drosophilák reciprok keresztezéséből származó különböző eredmények magyarázata. A muslicák nemét az X és Y kromoszómák határozzák meg. Közvetett bizonyíték

27
C. Bridges (1913) Közvetlen bizonyíték P xwxwY xw+0 F1 F2
Az elsődleges és másodlagos kivételeket eredményező Drosophila keresztezések. P xwxwY xw+0 F1 1/2ooo F2

28
Az elsődleges kivétel utódok létrejöttének magyarázata feltételezve, hogy a maternális szülőben az X kromoszómák nem váltak szét (non - diszjunkció). Red = piros-szemű White = fehér szemű

29
A kromoszóma elmélet kísérletes bizonyítéka
Az elsődleges kívétel nőstények és vad hímekkel keresztezése eredményeként kapott utódok neme és szem-fenotípusa megfelel annak, amit azon feltételezések alapján várunk, hogy a szemszín gén az X kromoszómán helyezkedik el, és az elsődleges kivétel nőstény XXY kromoszómákat hordoz. Ezen genetikai bizonyítékon túl Bridges megvizsgálta az utódok kromoszómáit is citológiailag, és azokat az elvárásnak megfelelőnek találta. Ezeket az eredményeket a kromoszóma elmélet citogenetikai bizonyítékainak tekintjük.

30
Nemi kromoszómák és a nemhez kötött öröklés
Az XY típusú nem-meghatározás: XX XY X0 XXY ember nő férfi Turner szindróma Kleinefelter szindróma muslica nőstény hím Steril

31
X-hez kötött recesszív allélek pedigréje mutatja, hogy az apákban kifejeződő rendellenesség nem jelentkezik lányaikban, azonban azok fiaiban újra megjelenik. A III-3 és III-4 egyedek fenotípusuk alapján nem különböztethetők meg.

32
X-hez kötött recesszív allél okozza a hemofiliát
X-hez kötött recesszív allél okozza a hemofiliát. A vérzékenységet okozó allél valószínűleg Viktória királynő ivarsejtjeiben keletkezett. A családon belüli házasodások miatt a rendellenesség elterjedt az egész családban.

33
Négy tesztikuláris feminizáció szindrómában szenvedő lánytestvér
Négy tesztikuláris feminizáció szindrómában szenvedő lánytestvér. Mind a négy paciens kromoszóma szerelvénye 44 autoszóma plusz X és Y kromoszóma. Örököltek egy X kromoszómához kötött recesszív allélt ami miatt érzéketlenek az androgénre (férfi nemi hormon).
.")
34
A férfiakban megjelenő X-hez kötött domináns fenotípust a lányaik öröklik.

35
X kromoszómához kötött domináns rendellenesség öröklődésének családfája. A terhelt nők általában heterozigóták, a rendellenességet a fiaik felére és a lányaikra örökítik. Ilyen rendellenesség a hipofoszfatémia, a D vitamin rezisztens angolkór egy fajtája.

36
Dóziskompenzáció X kromoszóma inaktiváció
A Barr test, kondenzált inaktivált X kromoszóma egy normális nő sejtmagjában. A férfiaknak nincs Barr testük. A Barr testek sejtenkénti száma mindig az összes X kromoszóma száma mínusz egy.

37
X – kromoszóma inaktíváció
- egerekben: az egyedfejlődés 3-6 napja során - emberben: korai embrióban, a sejtek differenciálódása előtt. Gén - dózis kompenzáció kromoszóma imprinting genetikai mozaikok The clonal inheritance of a condensed inactive X chromosome that occurs in female mammals.

38
Egy tarka macska. Mind a tarka mind a teknős mintázatú macskák nőstények. Egy X kromoszómához kötött bundaszín gén alléleire, O (orange = vörös) és o (fekete) heterozigóták. A vörös és fekete szektorokat az X kromoszóma inaktivációja okozza. A bunda fehér területeit a tarka macskán egy másik gén okozza.
 és o (fekete) heterozigóták. A vörös és fekete szektorokat az X kromoszóma inaktivációja okozza. A bunda fehér területeit a tarka macskán egy másik gén okozza.")
39
Három egymást követő nemzedéket alkotó,
a nemhez kötött ektodermális diszplaziára (verejtékmirigy hiány) heterozigóta nőben megfigyelhető szomatikus mozaicizmus. A verejtékmirigy hiányos terület kékkel jelölt.
 heterozigóta nőben megfigyelhető. szomatikus mozaicizmus. A verejtékmirigy hiányos terület kékkel jelölt.")
40
Ez egy olyan fenotípus, amit valószínűleg egy
Szőrös fülcimpa. Ez egy olyan fenotípus, amit valószínűleg egy Y kromoszómához kapcsolt gén okoz.

41
párhuzamos viselkedése
Autoszómális gének és kromoszómák párhuzamos viselkedése

43
Kromatin - DNS hiszton nem-hiszton fehérjék - RNS Kromoszóma

44
Milyen módszerrel tömörítődik a DNS a sejtmagban?
Egy kromoszóma egyetlen folytonos DNS molekula Milyen módszerrel tömörítődik a DNS a sejtmagban?

45
Kromatin - DNS hiszton nem-hiszton fehérjék - RNS Kromoszóma

46
A kromoszóma szakaszok egymástól elkülönülő szerkezeti és működési sajátosságait a nukleoszóma alapszerkezethez kapcsolódó nem-hiszton típusú kromoszómális fehérjék biztosítják. A nem-hiszton fehérjék kapcsolódhatnak - közvetlenül a DNS-hez (pl. topoizomeráz) - a hisztonokhoz, (és módosíthatják azokat pl. hiszton metilázok, foszfatázok). vagy közvetve másik nem-hiszton fehérjéhez (pl. transzkripciós faktorok).
 - a hisztonokhoz, (és módosíthatják azokat pl. hiszton metilázok, foszfatázok). vagy közvetve másik nem-hiszton fehérjéhez (pl. transzkripciós faktorok).")
47
Kétéltűek meiotikus „lámpakefe” kromoszómája.
(kétéltűek meiózis diplotén) 30 nm központi váz (scaffold)
 30 nm. központi váz. (scaffold)")
48
A Drosophilában az egyik térképezett szakasz hurok egységei
A Drosophilában az egyik térképezett szakasz hurok egységei. Megfigyelhetők a gének helyei és a scaffold kapcsolódó (SAR) szakaszok.
 szakaszok.")
49
Az eukarióta kromoszóma szerkezet modellje.
A scaffold váz további többszörös spiralizációval tömörödik. metafázis interfázis

50
interfázisos kromoszómák nyálmirigy óriáskromoszómák

51
Kromoszóma mutációk: deléció, duplikáció, inverzió, transzpozíció, transzlokáció deléció

52
replikációs origó Mitotikus kromoszómák

53
erőteljesebb kromatin feltekeredés. erőteljes génexpresszió.
kariotípus FISH Sávozási technikák: Giemsa – festés G – sötét sávok: erőteljesebb kromatin feltekeredés. G – világos sávok: erőteljes génexpresszió.

54
centromeron kinetochor mikrotubulus kromoszóma

55
Mitotic chromosome organization
HP1 heterochromatin Outer kinetochore: POLO, BubR1, Rod, ZW10, CENP-meta, CENP-E Aur B, KLP67A, KLP10A, Orbit, DCLIP190 CID / CENP-A inner kinetochore identity MEI-S332 centromeric sister chromatin cohesion PROD pericentric heterochomatin

56
The molecular motor on the kinetochor of the chromosome attaches it to the microtubule.
During metaphase, the motor pulls on the microtubule which causes the chromosome to move towards the MTOC. This movement seperates the sister chromatids so that 1 copy of each chromosome goes to each new cell. As this happens, the microtubules de-polymerize and get shorter.

57
Az eukarióta kromoszómák
heterokromatikus és eukromatikus szakaszra oszthatók

58
A heterokromatin helyzete a Drosophila melanogasterben.
(DNS festékkel festve) C = centromer NO = nukleólusz organizátor eukromatin heterokromatin A heterokromatin genetikailag inaktív, a genetikai analízis adatai szinte teljes mértékben az eukromatinra vonatkoznak.
 C = centromer. NO = nukleólusz organizátor. eukromatin heterokromatin. A heterokromatin genetikailag inaktív, a genetikai analízis adatai szinte teljes mértékben az eukromatinra vonatkoznak.")
59
PEV “pozíció – függő – variegáció”
ADE2 gene codes for one of the enzymes of adenine biosynthesis, and the absence of the ADE2 gene product leads to the accumulation of a red pigment. The white gene controls eye pigment production and is named after the mutation that first identified it.

60
kromoszómák - eukromatin - heterokromatin feltekeredési állapota
pericentrikus telomerikus fakultatív: konstitutív:

61
reversible modification
of histone N-tails metilálás, acetilálás, foszforilálás Histone – code ?

62
A heterokromatin és az eukromatin közötti főbb különbségek
1., A heterokromatin tömörebb szerkezetű az eukromatinnál, és az egész sejtciklus alatt erősen kondenzált. 2., A heterokromatin alig tartalmaz géneket. 3., A heterokromatin a centromer közelében található, az eukromatin a centromertől távolabb. 4., Az eukromatin mennyisége fajon belül állandó, a heterokromatin mennyisége egyedenként, néhol sejtenként változhat. 5., A heterokromatin mindig utoljára replikálódik. 6., A heterokromatint szatellita DNS alkotja.

63
chromatin remodeling complexes
Elősegítik a DNS – replikációt, repair-t és gén-expressziót. Sejtosztódás ideje alatt inaktívak. In the absence of remodeling complexes, the interconversion between the three nucleosomal states is very slow because of a high activation energy barrier. Using ATP hydrolysis, chromatin-remodeling complexes (green) create an activated intermediate in which the histone-DNA contacts have been partly disrupted. This activated state can then decay to any one of the three nucleosomal configurations shown. In this way, the remodeling complexes greatly increase the rate of interconversion between different nucleosomal states. The remodeled state, in which the histone-DNA contacts have been loosened, has a higher free energy level than that of standard nucleosomes and will slowly revert to the standard nucleosome conformation, even in the absence of a remodeling complex. Cells have many different chromatin remodeling complexes, and they differ in their detailed biochemical properties; for example, not all can change the position of a nucleosome, but all use the energy of ATP hydrolysis to alter nucleosome structure.
 create an activated intermediate in which the histone-DNA contacts have been partly disrupted. This activated state can then decay to any one of the three nucleosomal configurations shown. In this way, the remodeling complexes greatly increase the rate of interconversion between different nucleosomal states. The remodeled state, in which the histone-DNA contacts have been loosened, has a higher free energy level than that of standard nucleosomes and will slowly revert to the standard nucleosome conformation, even in the absence of a remodeling complex. Cells have many different chromatin remodeling complexes, and they differ in their detailed biochemical properties; for example, not all can change the position of a nucleosome, but all use the energy of ATP hydrolysis to alter nucleosome structure.")
64
kromoszóma ciklus interfázis mitózis kondenzáció dekondenzáció
replikáció

65
A DNS szekvenciák típusai és szerveződésük
a kromoszómában

66
Repetitív DNS főbb típusai:
A prokarióta genom egyszerű szerveződésű, szinte csak kódoló részeket tartalmaz. Az eukarióta genomok jelentős része (átlag 20-50%) nem kódoló szekvenciákat tartalmaz. Ezek jelentős része több példányban lévő repetitív DNS. Repetitív DNS főbb típusai: Géncsaládok (szétszort vagy tandem elhelyezkedés) Nem kódoló repetitív szekvenciák - a szatellit DNS - a miniszatellit DNS - a mikroszatellit DNS - a telomerikus repeatek - az ugráló gének és maradványaik
 nem kódoló szekvenciákat tartalmaz. Ezek jelentős része több példányban lévő repetitív DNS. Repetitív DNS főbb típusai: Géncsaládok (szétszort vagy tandem elhelyezkedés) Nem kódoló repetitív szekvenciák. - a szatellit DNS. - a miniszatellit DNS. - a mikroszatellit DNS. - a telomerikus repeatek. - az ugráló gének és maradványaik.")
67
A legtöbb gén diploidokban 2 (homológ) példányban van jelen.
Egy izom fehérjére specifikus, fluoreszcensen jelölt próba használatával végzett kromoszóma in situ hibridizáció. Mindössze egyetlen lókusz mutat fluoreszcens pontot, ami megfelel a próba izom fehérje génjéhez kötődésének.

68
Szétszórt géncsaládok
Néhány gén a genomban szétszóródva 2-nél több példányban található. Ezeket géncsaládoknak nevezzük. Egy családon belül a gének nukleotid sorrendje különbözhet, sőt az evolúció során a homológok eltérő funkciót is nyerhetnek. (pl. humán hemoglobinok). Egy családon belül egyes gének mutációk révén teljesen el is veszthetik a funkcióképességüket. Így keletkeznek az át nem íródó pszeudogének. Példák: Aktin 5-30 példány Keratin >20 példány Miozin nehéz lánc 5-10 példány Tubulin 3-15 példány Peteburok fehérje 50 példány Globinok 5 példány Immunglobulin variábilis szakasz 500 példány Ovalbumin 3 példány
. Egy családon belül egyes gének mutációk révén teljesen el is veszthetik a funkcióképességüket. Így keletkeznek az át nem íródó pszeudogének. Példák: Aktin 5-30 példány Keratin >20 példány. Miozin nehéz lánc 5-10 példány Tubulin 3-15 példány. Peteburok fehérje 50 példány Globinok 5 példány. Immunglobulin variábilis szakasz 500 példány. Ovalbumin 3 példány.")
69
Tandem géncsaládok Bizonyos gének termékéből a sejt nagy mennyiséget igényel. Ezen gének családjai egymás mellett elhelyezkedő sorozatokat alkotnak. Ilyenek például a riboszómális RNS-ek génjei a tRNS génjei és a hiszton gének. A riboszómális RNS-t kódoló gének helye nukleólusz organizátor (NO), citológiailag a másodlagos befűződés helye a kromoszómán.
, citológiailag a másodlagos befűződés helye a kromoszómán.")
70
A hiszton gének a legtöbb élőlényben tandem géncsaládokat képeznek,
az ismétlődések száma lehet. Tengeri sün és muslica hiszton gének tandem ismétlődései. Az ábra az ismétlődések csak egy kis részét mutatja. A nyilak a transzkripció irányát jelölik.

71
Nem kódoló repetitív szekvenciák
- a szatellit DNS - a miniszatellit DNS - a telomerikus repeatek - az ugráló gének és maradványaik

72
Sokszor ismétlődő heterokromatikus DNS
szatellita DNS sávok genomi DNS CsCl oldat Genomikus DNS céziumklorid centrifugálása után gyakran a fő DNS száltól elkülönülve szatellit DNS sávok figyelhetők meg. A szatellit DNS-t elsősorban rövid tandem ismétlődések alkotják, amik akár több ezer kb hosszúak is lehetnek. Pl: AAGAC AAGAC AAGAC

73
A hatalmas szatellit DNS blokkok alkotják a kromoszómák
A hatalmas szatellit DNS blokkok alkotják a kromoszómák heterokromatinját a Drosophia 2. kr. heterokromatin összetétele: Cm 2L 2R (AAGAG)n (AAGAGAG)n (AATAG)n (AATAACATAG)n (AAGAC)n A repetitív DNS túlnyomó részét a heterokromatikus szatellit DNS adja. Valószínűleg ez az alapja a heterokromatin kondenzált szerkezetének. A muslica genomnak pl. 30%-a szatellit DNS, egyes kenguru fajokban a genom 50 % szatellit DNS.
n. (AAGAGAG)n. (AATAG)n. (AATAACATAG)n. (AAGAC)n. A repetitív DNS túlnyomó részét a heterokromatikus szatellit DNS adja. Valószínűleg ez az alapja a heterokromatin kondenzált szerkezetének. A muslica genomnak pl. 30%-a szatellit DNS, egyes kenguru fajokban a genom 50 % szatellit DNS.")
74
A miniszatellitt DNS vagy VNTR
Az eukromatinban szétszórt rövidebb-hosszabb tandem repetitív DNS blokkokat miniszatellit DNS-nek hívják, angolul variable number of tandem repeats-nek, vagy VNTR-nek. Ezek 1-5 kb hosszú szakaszok, amik nukleotidnyi ismétlődéseket tartalmaznak. A VNTR-ek vizsgálatát restrikciós enzimmel hasított genomikus DNS-en végzik. Olyan enzimet használnak, ami a VNTR-en belül nem hasít. Az ilyen DNS VNTR próbával végzett Southern hibridizációja egyedről egyedre különböző mintázatot ad. A mintázatot DNS ujjlenyomatnak nevezik. Az igazságügyi orvostanban fontos bizonyítási módszer.

75
VNTR-ek Egy bűntény színhelyéről származó vérminta és hét gyanúsítottból származó DNS ujjlenyomata.

76
Ugráló gének (transzpozonok)
Az eukarióta genom repetitiv szekvenciáinak jelentős részét önálló replikációra és áthelyződésre képes DNS-sorrendek, vagy ezek származékai alkotják. Ezek: Transzpozonok – később, külön tárgyaljuk Retrotranszpozonok, retrovírusokra emlékeztetnek. Például: Drosophila copia elemei, 5 kb hosszú, 50/genom élesztő Ty eleme, 6 kb, 30/genom emlősök LINE (long interspersed elements) 1-5 kb, Nem-retrovírus szerű retrotranszpozonok, rövid pár-száz nukleotidpár hosszú ismétlődések, SINE, short interspersed elements-nek is nevezik pl. Alu elemek. 200 bp, a humán genom 5 %-át teszik ki.
 1-5 kb, Nem-retrovírus szerű retrotranszpozonok, rövid pár-száz nukleotidpár. hosszú ismétlődések, SINE, short interspersed elements-nek is nevezik. pl. Alu elemek. 200 bp, a humán genom 5 %-át teszik ki.")
77
Az eukarióta kromoszóma szerkezet általános képe.
A kromoszóma egy kis része látszik, öt fehérjét kódoló génnel, a nukleólusz organizátor egyik vége, és a centromer heterokromatin vége. Az ismétlődő sorrendek különböző féleségei látszanak. (Minden kromoszóma normálisan a gének ezreit hordozza.)
")
78
the end

Hasonló előadás

 DNS-ből,>")







 Bihari Péter.>")









