Előadást letölteni
Az előadás letöltése folymat van. Kérjük, várjon

1
MIÉRT NEM MÉRHETŐ? E + S P + E mol/dm3!!!!
BIM BSc 2007 ENZIMKINETIKA E S P E mol/dm3!!!! MIÉRT NEM MÉRHETŐ? Egy egység az az enzim mennyiség, amely 1 mol szubsztrátot alakít át vagy 1 mol terméket képez 1 perc alatt adott reakció körülmények között. SI rendszerben: 1 Katal: 1 mol szubsztrátot alakít át 1 s alatt. hatalmas enzim mennyiség nKat = 10-9 Kat 1 Kat = 6*107 U, 1U =1.6*10-8 Kat, U= 1/60 Kat E/tf E/mg fajlagos aktivitás Michaelis és Menten

2
Michaelis-Menten kinetika
E + S ES E P k 1 -1 2 -2 feltételezések: *k-2 =0 *első lépés gyorsan egyensúlyra jut= RAPID EKVILIBRIUM *stabil ES komplex, EP komplex elhanyagolható *egy aktiv centrum, egy szubsztrát aktivitás helyett cc használható S E0 vagyis E0 / S < <1 k1SE = k-1 (ES) a „minket érdeklő” reakciósebesség: az (ES) disszociációs állandója anyagmérleg osszuk el ezt a kettőt egymással
 a „minket érdeklő reakciósebesség: az (ES) disszociációs. állandója. anyagmérleg. osszuk el ezt a kettőt egymással.")
3
Michaelis-Menten kinetika
Rendezzük át!

4
Michaelis-Menten kinetika

5
terméket adó komplexek összes komplex terméket nem adó komplex szabad enzim MEMO M-M séma elképzelt séma alapján alapján

6
M és M

7
Maud Leonora Menten (1879–1960) was born in Port Lambton, Ontario, and in 1911 at the University of Toronto she became one of the first Canadian women to be qualified in medicine, from which she went on to obtain a PhD at the University of Chicago for aspects of cancer biochemistry. Her work with Leonor Michaelis on invertase was an interlude in a career devoted mostly to pathology and the more medical aspects of biochemistry and physiology. She coauthored more than 70 publications, and was the first to use electrophoretic mobility to study human hemoglobins. She spent most of her working life at the University of Pittsburgh, but went back to Canada after her retirement, where her research continued at the Medical Institute of British Columbia until it was brought to an end by ill health. She then returned to Ontario to spend the remainder of her life not far from where she was born. Leonor Michaelis (1875–1949) was born in Berlin. He worked for a year as assistant to Paul Ehrlich, and afterwards studied clinical medicine and developed an early interest in controlling the hydrogen ion concentration. In the years preceding the First World War he used his mastery of this subject to become one of the leaders in studying enzyme-catalyzed reactions. This was an impressively productive period for him, and his best-known paper is just one of 94 publications, including five books, in the five years from 1910 to Several of his papers from these years are still cited, and one of the books, Die Wasserstoffionenkonzentration, became the standard work on pH, buffers and related topics. In the 1920s he spent three years as professor of biochemistry in Nagoya, Japan, and subsequently moved to the USA; from 1926 to 1929 he was at the Johns Hopkins University, and afterwards he was at the Rockefeller Institute for Medical Research. Until the end of his life he remained active in research, concerned mainly with the study of free radicals. In addition to his scientific achievements, Michaelis was a violinist of near-professional standard. Azzi relates that during his time in Japan he was asked by Shinichi Suzuki, the son of a violin maker, if he was suited to a career as a soloist. Michaelis advised him to go into teaching, thereby catalyzing the birth of the Suzuki method of teaching the violin. (Fundamentals of Enzyme Kinetics, 4th Edition Athel Cornish-Bowden January 2012)
 was born in Berlin. He worked for a year as assistant to Paul Ehrlich, and afterwards studied clinical medicine and developed an early interest in controlling the hydrogen ion concentration. In the years preceding the First World War he used his mastery of this subject to become one of the leaders in studying enzyme-catalyzed reactions. This was an impressively productive period for him, and his best-known paper is just one of 94 publications, including five books, in the five years from 1910 to Several of his papers from these years are still cited, and one of the books, Die Wasserstoffionenkonzentration, became the standard work on pH, buffers and related topics. In the 1920s he spent three years as professor of biochemistry in Nagoya, Japan, and subsequently moved to the USA; from 1926 to 1929 he was at the Johns Hopkins University, and afterwards he was at the Rockefeller Institute for Medical Research. Until the end of his life he remained active in research, concerned mainly with the study of free radicals. In addition to his scientific achievements, Michaelis was a violinist of near-professional standard. Azzi relates that during his time in Japan he was asked by Shinichi Suzuki, the son of a violin maker, if he was suited to a career as a soloist. Michaelis advised him to go into teaching, thereby catalyzing the birth of the Suzuki method of teaching the violin. (Fundamentals of Enzyme Kinetics, 4th Edition Athel Cornish-Bowden. January 2012)")
8
BRIGGS-HALDANE KINETIKA
d(ES)/dt =0 steady state S Eo vagy Eo / S 1 és (k1ES k-1(ES) ill k1ES k2(ES))
/dt =0. steady state. S Eo vagy Eo / S 1 és. (k1ES k-1(ES) ill. k1ES k2(ES))")
9
B-H megoldása szimu- láció

10
BRIGGS-HALDANE KINETIKA
Km=(k-1 + k2) / k1 Michaelis állandó
 / k1. Michaelis állandó.")
11
ha num(k1)> >num(k2) a két konst. azonos!
DISZKUSSZIÓ Michaelis -Menten Briggs-Haldane ha num(k1)> >num(k2) a két konst. azonos!
> >num(k2) a két konst. azonos!")
12
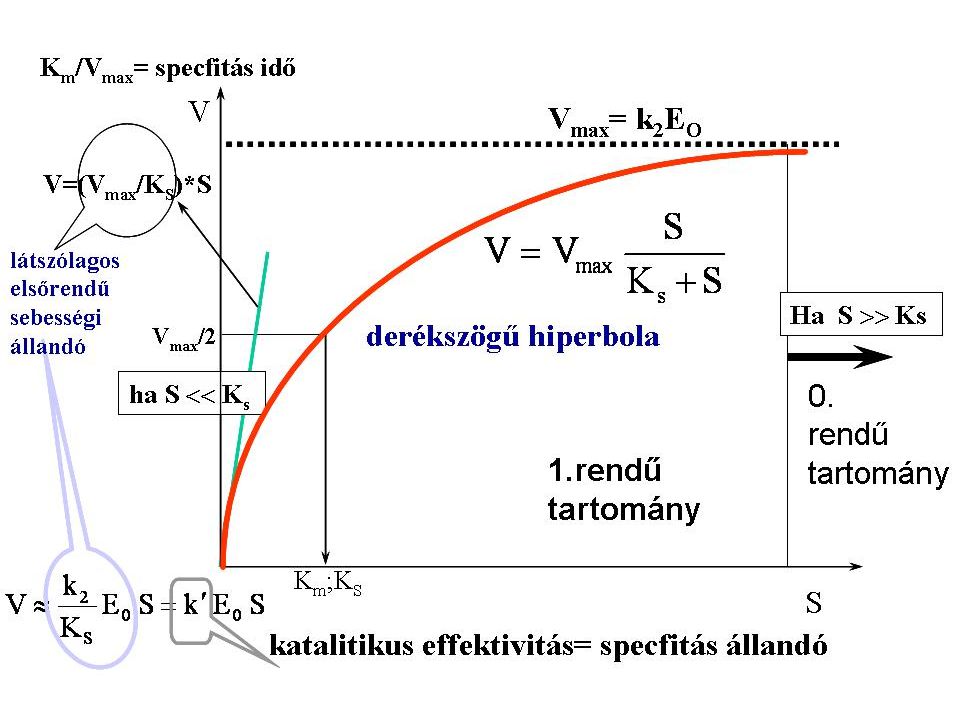
Km/Vmax= specfi(ci)tás idő
DISZKUSSZIÓ V=(V max /K S )*S 1.rendû tartomány 0. rendű V = k 2 E O K m ; K Km/Vmax= specfi(ci)tás idő Ha S Ks derékszögű hiperbola S Ks látszólagos elsőrendű sebességi állandó katalitikus effektivitás= specfi(ci)tás állandó
*S. 1.rendû. tartomány. 0. rendű. V. = k. 2. E. O. K. m. ; K. Km/Vmax= specfi(ci)tás idő. Ha S Ks. derékszögű hiperbola. S Ks. látszólagos elsőrendű sebességi állandó. katalitikus effektivitás= specfi(ci)tás állandó.")
14
hiperbola V V max értelmes tartomány S K m -K m

15
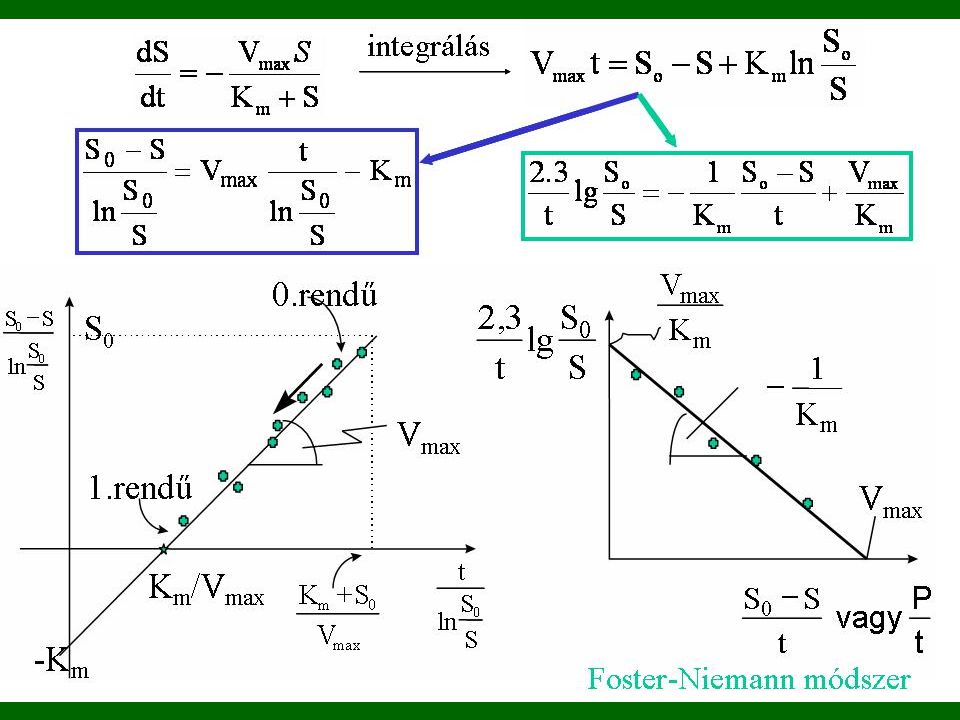
PARAMÉTERBECSLÉS 1 1. INTEGRÁLT ALAKBÓL Foster-Niemann

17
1/V 1/S -1/Km Paraméterbecslés 2
2. Lineweaver-Burk módszer linearizálás, dupla reciprok 1/S -1/Km tga=Km/Vmax 1/V 1/Vmax

18
S/V Km/Vmax Km S Paraméterbecslés 3
3. HANES v. LANGMUIR módszer linearizás tga=1/Vmax S/V S Km/Vmax Km

19
4. EADIE-HOFSTEE módszer linearizálás
Paraméterbecslés 4 V V/S Vmax tga=-Km Vmax/Km Vmax/Km V/S tga=-1/Km V Vmax

20
L-B, L-H, E-H ábrázolások
1/V S/V 1/S -1/Km tga=Km/Vmax tga=1/Vmax Km/Vmax 1/Vmax Km S V Vmax tga=-Km Vmax/Km V/S

21
A M-M és B-H egyenletekben V kezdeti reakciósebességet jelent!!!
V0 értelmezése A M-M és B-H egyenletekben V kezdeti reakciósebességet jelent!!! V0=(dS/dt)t=0 S t P V0= (dP/dt)t=0
t=0. S. t. P. V0= (dP/dt)t=0.")
22
Egyéb grafikus módszerek 1
V max = v + S K m Direkt lineáris ábrázolás Eisenthal és Cornish-Bowden (1974) Egy megfigyelés V V max V4 V3 V2 V1 -[S] [S] -4S -3S -2S -1S
 Egy megfigyelés. V. V. max. V4. V3. V2. V1. -[S] [S] -4S. -3S. -2S. -1S.")
23
Egyéb grafikus módszerek 2
V max = v + S K m (a hiperbola megszerkesztése) V Vmax S -Km
 V. Vmax. S. -Km.")
24
A k2 meghatározása és Vmax E0-függése
Vmax3=k2EO3 Vmax2=k2EO2 Vmax1=k2EO1 Km EO1 EO2 EO3 EO S V tg = k2 Tehát így mérünk enzim aktivitást!!!

25
Dimenziómentes hiperbola
V’ =V/Vmax V’ S’=S/SO Km’=Km/SO = + V max és S valamint K bevezetésekkel m

26
A kinetikai paraméterek értelmezése
1 Vmax IUBMB:nem max,hanem Limit!!! HATÁRSEBESSÉG

27
A kinetikai paraméterek értelmezése
1 Vmax IUBMB:nem max, hanem limit!!! HATÁRSEBESSÉG NEM ENZIMTULAJDONSÁG Vmax= k2 . E = AKTIVITÁS k [s-1] ENZIMTULAJDONSÁG = turnover number, váltásszám Kiterjesztés minden enzimre és minden kinetikára kcat: [ s-1 ] Egy enzimmolekula átalakítási frekvenciája S-telítés esetén Vmax= kcat . E0 érzéketlen Km , KS Közelítőleg az S az élő sejtben az enzim affinitása A = B ??? Változott a KS Inhibitor?Aktivátor? Enzimanalitika S>>KS Túl érzékeny

28
A kinetikai paraméterek értelmezése 2
k dm3mol-1min-1 max. érték(1011) kis molekulák diffúzió-sebessége k min-1 k min-1 Km mol/dm3
 kis molekulák diffúzió-sebessége k min-1. k min-1. Km mol/dm3.")
29
α-amiláz: 500 s-1 , glükoamiláz: 160 s-1 , glükózizomeráz: 3 s-1

30
Legtöbb enzim e két szélső eset között
kcat alsó határa metabolikus enzimeknél Legtöbb enzim e két szélső eset között Természetes enzimeknél: >105 Mesterséges e-nél (DNA-zyme, abzyme:<103

31
Ha az enzim móltömege 60000 és fajlagos aktivitása 1 U/mg, akkor kcat éppen 1 s-1
Ms= U/mg kcat=1 s-1

32
REVERZIBILIS REAKCIÓK 1
Sok enzim katalizálta reakció - főként a biopolimer hidrolízisek - nagymértékben a jobboldali irányba eltolt egyensúllyal rendelkeznek,→ gyakorlatilag k-2 valóban elhanyagolható. De például a glükóz fruktóz (glükóz izomeráz)gyakorlatban is egyensúlyi reakcióként viselkedik E + S ES E + P k1 k-1 k2 k-2 KP 1/KS
gyakorlatban is egyensúlyi reakcióként viselkedik. E + S ES E + P. k1. k-1. k2. k-2. KP. 1/KS.")
33
REVERZIBILIS REAKCIÓK 2
Végezzük el a következő osztásokat: HALDANE összefüggés

34
REVERZIBILIS REAKCIÓK 3
? MI TÖRTÉNIK? S → P vagy P → S S MITŐL FÜGG? Keq , S , P értéke P Vnetto = Velőre - Vvissza = k2 (ES) - k-2(EP) Reverzibilis M-M egyenlet
 - k-2(EP) Reverzibilis M-M egyenlet.")
35
Ez a reverzibilis M-M enzimkinetika általános alakja

Hasonló előadás