Előadást letölteni
Az előadás letöltése folymat van. Kérjük, várjon

1
Az “sejt gépei” az enzimek
papír + O füst + hamu + hő + CO2 + H2O A kémiai reakciók mindig a szabadenergia csökkenés irányába mennek végbe. Miért nem alakul át minden anyag a számára legalacsonyabb energiájú, legstabilabb állapotába? Válasz: aktivációs energiagát Az enzimek ezt az aktivációs energiagátat csökkentik.

2
Az enzimek ezt az aktivációs energiagátat csökkentik.

3
Az enzimek különösen nagy mértékben gyorsítják meg a reakciókat.
Minek tudható ez be? A katalitikus hely környezetében megnövelik a szubsztrátkoncentrációt A szubsztrát megkötéséből eredő kötési energia hozzájárul a közvetlen katalízishez Az enzimek jóval nagyobb affinitással bírnak az átmenet állapotú szubsztrát, mint a stabil végtermék iránt Sav és báziskatalízis

4
Enzimtulajdonságok Csak termodinamikailag lehetséges reakciót katalizálnak Mivel „csak” az aktivációs energiát csökkentik: Biokatalizátorok

5
Kapcsolt reakciók DG értéke negatív (exergonikus reakció): spontán, energiabevitel nélkül végbemegy DG értéke pozitív (endergonikus reakció): nem megy végbe spontán Végbemehet, ha egy exergonikus reakcióval összekapcsoljuk és az eredő szabadenergiaváltozás negatív.
: nem megy végbe spontán. Végbemehet, ha egy exergonikus reakcióval összekapcsoljuk és az eredő szabadenergiaváltozás negatív.")
7
Katalitikus antitestek
Orientációs hatás a b c a., az enzim megköti és pontosan orentálja egymáshoz a szubsztrátokat b., a szubstrát megkötésével az enzim átrendezi annak elektroneloszlását, részlegesen + és - részeket eredményezve. c., az enzim megfeszíti a megkötött szubsztrát molekulát, ezzel az átmeneti állapot felé tolva Katalitikus antitestek

8
Enzimtulajdonságok Csak termodinamikailag lehetséges reakciót katalizálnak Mivel „csak” az aktivációs energiát csökkentik: Biokatalizátorok Maguk nem változnak a reakció során Az enzimek fehérjemolekulák Holoenzim = apoenzim + koenzim

9
Az enzimhez kapcsolódó nem fehérje alkotók, prosztetikus csoportok
Az enzimek működésükhöz nem fehérje, nem aminosav részeket is igényelnek. - fémionok - szerves molekulák kovalens kötődéssel pl.: biotin, liponsav nem kovalens kötődéssel pl.: NAD, NADP+, tetrahidrofólsav

10
Csak termodinamikailag lehetséges reakciót katalizálnak
Enzimtulajdonságok Csak termodinamikailag lehetséges reakciót katalizálnak Mivel „csak” az aktivációs energiát csökkentik: Biokatalizátorok Maguk nem változnak a reakció során Az enzimek fehérjemolekulák, melyeket nem fehérje természetű részek egészíthetnek ki. Holoenzim = apoenzim + koenzim Specifikusak: - Szubsztrát - Reakció pl.: hexokináz, trombin Érzékenyek a környezeti hatások, körülményrkre (pH, hőmérséklet, ionerősség, koncentráció)
")
11
Fehérjeszerkezet és enzimműködés
E + S [ES] 1. Kulcs-zár elmélet: a szubsztrát pontosan az enzik kötőhelyének kiegészítése. Emil Fischer 1890 aktív centrum koenzimek kötő hely katalítikus hely

12
Az aktív hely kialakításában az enzim szerkezetének csak kis része vesz részt.
E-S kötődés: gyenge másodlagos kötések: ionos, hidrogén kötés, hidrofób kölcsönhatások 2. Indukált illeszkedési elmélet (Induced fit): A szubsztrát bekötése módosítja az enzim térszerkezetét. Koshland 1958. Az új enzim konformáció hozzájárul a megnövekedett katalítikus aktivitáshoz.
: A szubsztrát bekötése módosítja az enzim térszerkezetét. Koshland Az új enzim konformáció hozzájárul a megnövekedett katalítikus aktivitáshoz.")
13
Enzimkinetikai alapok
E + S ES E + P Reakciósebesség: az időegység alatt átalakított szubsztrát, vagy képződő termék mennyisége. A nem katalizált reakciók sebessége a reakcióban részt vevő anyagok koncentrációjának függvénye. Enzimes reakciók: Az adott idő alatt megkötött és átalakított szubsztrát mennyisége behatárolt. Az enzim egy pontban telítödik (Vmax)
")
14
„A postásnak is csak két keze van.”
vmax vmax/2 Nem katalizált reakció KM

15
Ha két postás 1 óra alatt 250 levelet tud kézbesíteni.

16
Akkor négy postás 1 óra alatt hány levelet kézbesít?
Megjegyzés: a körülmények azonosak

17
vmax3 = k2E03 vmax2 = k2E02 vmax1 = k2E01

18
A Michaelis-Menten modell
Maud Menten Leonor Michaelis k1 k2 k-1 E + S ES E + P Kiindulási feltételek, egyszerűsítések: Az első lépés gyors ekvilibriuma A második lépés irreverzibilis Kezdeti sebességet vizsgálunk E<<[S]

19
E + S ES E + P ES fogyás = ES képződés k-1ES + k2ES = k1 ES
E = EO - ES ES = ES * k1/(k-1 + k2) = (EO - ES)S * k1/(k-1 + k2) Km = (k-1 + k2)/ k1 ES = (EO - ES)S * 1/ Km ES Km + ES S = EO S ES = EO S / (Km + S) v = k2ES v = k2EO S / (Km + S) v = vmax = k2EO v = vmaxS / (Km + S) k1 k2 k-1 E + S ES E + P
 = (EO - ES)S * k1/(k-1 + k2) Km = (k-1 + k2)/ k1. ES = (EO - ES)S * 1/ Km. ES Km + ES S = EO S ES = EO S / (Km + S) v = k2ES v = k2EO S / (Km + S) v = vmax = k2EO v = vmaxS / (Km + S) k1. k2. k-1. E + S ES E + P.")
20
vmax v = 0,5vmax 0,5 vmax = [S]vmax/ [S] +KM [S] +KM=2 [S] KM= [S] vmax/2 KM
![vmax v = 0,5vmax 0,5 vmax = [S]vmax/ [S] +KM [S] +KM=2 [S] KM= [S] vmax/2 KM](http://slideplayer.hu/slide/1922172/7/images/20/vmax+v+%3D+0%2C5vmax+0%2C5+vmax+%3D+%5BS%5Dvmax%2F+%5BS%5D+%2BKM+%5BS%5D+%2BKM%3D2+%5BS%5D+KM%3D+%5BS%5D+vmax%2F2+KM.jpg "vmax v = 0,5vmax 0,5 vmax = [S]vmax/ [S] +KM [S] +KM=2 [S] KM= [S] vmax/2 KM")
21
Az enzimaktivitás szubsztrát- koncentráció- függésének és jellemző kinetikai paramétereinek meghatározása A reakciósebesség függése a szubsztrátkoncentrációtól A szubsztrát, a termék és az enzim koncentrációjának időbeli alakulása

22
Lineweaver-Burk féle linearizált ábrázolásmód
v = vm[S]/(KM+[S]) 1/v = ([S] + KM)/vm[S] 1/v = [S]/vm[S] + KM/vm[S] 1/v = 1/vm + 1/[S] * KM/vm 1/v tga = KM/vm 1/vm 1/S
 1/v = ([S] + KM)/vm[S] 1/v = [S]/vm[S] + KM/vm[S] 1/v = 1/vm + 1/[S] * KM/vm. 1/v. tga = KM/vm. 1/vm. 1/S.")
23
A kinetikai paraméterek értelmezése:
vmax: arányos a jelenlévő enzim mennyiségével, lényegében az enzimaktivitás mérőszáma, az enzim mennyiségéről ad információt. vmax3 = k2E03 vmax2 = k2E02 vmax1 = k2E01 Függ az enzim mennyiségétől, nem enzimtulajdonság. A k2 sebességi állandó azonban már enzimtulajdonság.

24
Átviteli szám: a vmax és az enzimkoncentráció hányadosa
kcat= vmax/E0 (1 és közötti szám, általában ~ 1000) egy enzimmolekula által 1 másodperc alatt átalakított szubsztrátmolekulák száma. KM: Megadja a szubsztrát koncentrációját az enzim környezetében. Állandó egy adott enzimre, alkalmas lehet az enzim azonosítására. Befolyásolásával szabályozható az enzim aktivitása. Az szubsztrát enzim irányába mutatott affinitását mutatja.
 egy enzimmolekula által 1 másodperc alatt átalakított. szubsztrátmolekulák száma. KM: Megadja a szubsztrát koncentrációját az enzim környezetében. Állandó egy adott enzimre, alkalmas lehet az enzim azonosítására. Befolyásolásával szabályozható az enzim aktivitása. Az szubsztrát enzim irányába mutatott affinitását mutatja.")
25
Enzimaktivitás: időegység alatt adott reakciókörülmények között átalakított szubsztrát, vagy képzett termék mennyisége. 1 Unit: 1mmol/perc 1 katal: mol/s (SI) Fajlagos aktivitás: egységnyi mennyiségű fehérjére eső enzimaktivitás (mmol/perc/mg fehérje)
 Fajlagos aktivitás: egységnyi mennyiségű fehérjére eső enzimaktivitás (mmol/perc/mg fehérje)")
27
Enzimgátlások Irreverzibilis: Az enzim valamely katalízisben szerepet játszó funkcionális csoportját teszik tönkre, vagy szorosan akár kovalens kötéssel hozzákapcsolódnak (pl.: nehézfémek). Az irreverzibilis gátlás ténylegesen csökkenti az aktív enzimmennyiséget. Reverzibilis: A reverzibilis gátlószerek dinamikus komplexet képeznek az enzimmel. A gátlás fajtáit megkülönböztetjük a vmax és KM kinetikai paraméterekre kifejtett hatásuk szerint. Kompetitív: KM nő Nemkompetitív: vmax csökken Unkompetitív: vmax, KM csökken
. Az irreverzibilis gátlás ténylegesen csökkenti az aktív enzimmennyiséget. Reverzibilis: A reverzibilis gátlószerek dinamikus komplexet képeznek az enzimmel. A gátlás fajtáit megkülönböztetjük a vmax és KM kinetikai paraméterekre kifejtett hatásuk szerint. Kompetitív: KM nő. Nemkompetitív: vmax csökken. Unkompetitív: vmax, KM csökken.")
28
Kompetitív: az enzimhez kötődve megakadályozzák a szubsztrát bekötődésétr, versengés a kötőhelyért. Általában szubsztrátanalógok, de nem feltétlenül. Magas szubsztrátkoncentrációnál leszorul a gátlószer az enzimről. Nemkompetitív: A gátlószernek nincs hatása a szubsztrát kötődésére. Azok egymástól függetlenül az enzim különböző helyeire kötődnek. A szubsztrátkoncentráció emelése nem képes eliminálni hatását. Unkompetitív: A gátlószer nem képes a szabad enzimhez, csak a már előzetesen létrejött ES komplexhez kapcsolódni.

29
Kompetitív gátlás: vmax változatlan, KM nő Nemkompetitív gátlás: vmax csökken, KM változatlan Unkompetitív gátlás: vmax csökken KM csökken

30
Az enzimgálások kettős reciprok (Lineweaver-Burk) ábrázolása
kompetitív nemkompetitív unkompetitív

31
Az enzimaktivitás közvetlen szabályozásának módjai
1. Allosztérikus enzimek Allos = másik Steros = szilárd, vagy három dimenziós Aktív hely: szubsztrát Regulációs hely: regulátor molekula Kommunikáció A regulátor molekula bekötődése befolyásolja az aktív helyet: konformációváltozás aktivitásváltozás

32
Az X molekula nem az aktív helyhez kötődik
Nem kell, hogy bármilyen kapcsolat legyen közte és a glukóz között. Az X molekula egyszerűen be- (+ reguláció) vagy kikapcsolja (- reguláció) az enzimet. Az allosztérikus fehérjék általános kapcsolóként működhetnek, segítve az anyagcsereutak összehangolását.
 vagy kikapcsolja. (- reguláció) az enzimet. Az allosztérikus fehérjék általános kapcsolóként működhetnek, segítve az anyagcsereutak összehangolását.")
33
2. Fehérjeszabályozás foszforiláció/defoszforiláció által
Allosztéria: pillanatszerű asszociáció/disszociáció (másodrendű kötések) Fehérje foszforiláció: Lassabb, kovalens módósítás 3. Limitált proteolízis proteolízis: peptidkötések bontása limitált: csak jól definiált helyen történő Általában proteáz enzimek aktiválása történik így, hogy csak az adott helyen (emésztőenzimek) és csak az adott körülmények között (véralvadási enzimek) legyenek aktivak. Proformában zimogénként szintetizálódnak és csak késöbb aktiválódnak.
 Fehérje foszforiláció: Lassabb, kovalens módósítás. 3. Limitált proteolízis. proteolízis: peptidkötések bontása. limitált: csak jól definiált helyen történő. Általában proteáz enzimek aktiválása történik így, hogy csak az adott helyen (emésztőenzimek) és csak az adott körülmények között (véralvadási enzimek) legyenek aktivak. Proformában zimogénként szintetizálódnak és csak késöbb aktiválódnak.")
34
A pH és a hőmérséklet hatása az enzimreakciók sebességére
Arhenius Hődenaturáció burkológörbék

35
Izoenzimek: Azonos funkciót ellátó (azonos reakciót katalizáló) azonban különböző (elsődleges) fehérjeszerkezetű enzimmolekulák. Az izoenzimek különbözhetnek: - szabályozásukban - sejten belüli elhelyezkedésükben - különböző szervek, szövetek közötti megoszlásukban -az általuk katalizált reakció kinetikai paramétereiben: vmax, KM -stabilitásukban

36
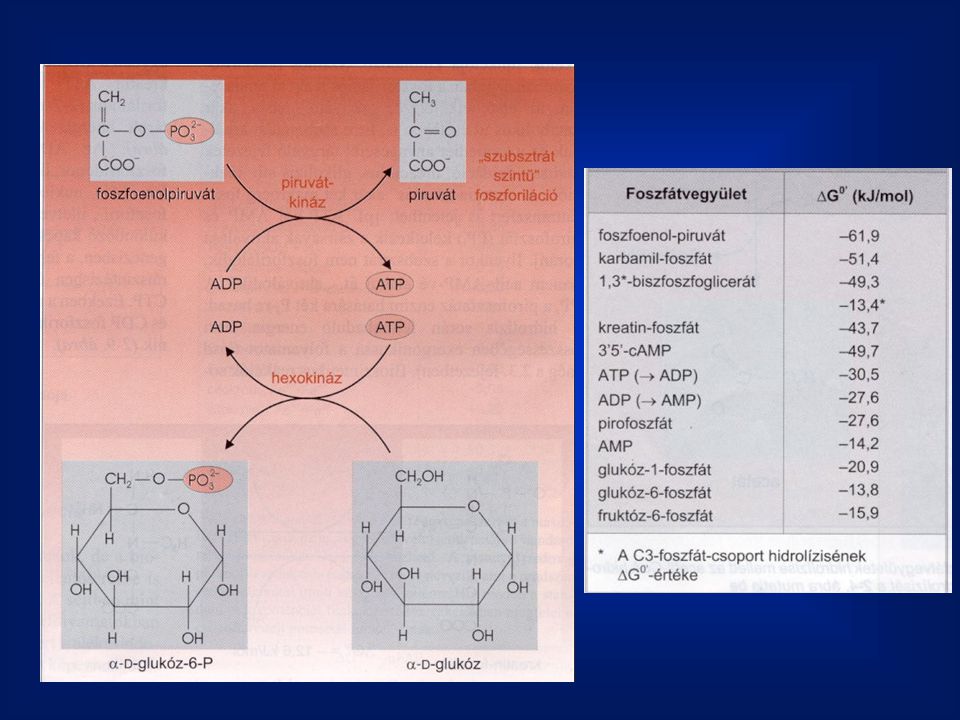
Az enzimek osztályai A hasonló reakciókat katalizáló enzimeket egyazon osztályba sorolták. (Enzyme Comission, EC). Négy számmal jelöli az enzimeket: 1. osztály, 2. alosztály, 3. csoport, 4. az adott enzim 1. Oxidoreduktázok pl.: etanol + NAD acetaldehid + NADH + H+ 2. Transzferázok pl.: glukóz + ATP glukóz-6 foszfát + ADP 3. Hidrolázok pl.: glukóz-6 foszfát + H2O glukóz + Pi 4. Liázok pl.: 2-foszfoglicerát foszfoenol-piruvát + H2O 5. Izomerázok pl.: glukóz-6 foszfát frutóz-6 foszfát 6. Ligázok pl.: glutamát + NH3 + ATP glutamin + ADP + Pi
. Négy számmal jelöli az enzimeket: 1. osztály, 2. alosztály, 3. csoport, 4. az adott enzim. 1. Oxidoreduktázok pl.: etanol + NAD+ acetaldehid + NADH + H+ 2. Transzferázok pl.: glukóz + ATP glukóz-6 foszfát + ADP. 3. Hidrolázok pl.: glukóz-6 foszfát + H2O glukóz + Pi. 4. Liázok pl.: 2-foszfoglicerát foszfoenol-piruvát + H2O. 5. Izomerázok pl.: glukóz-6 foszfát frutóz-6 foszfát. 6. Ligázok pl.: glutamát + NH3 + ATP glutamin + ADP + Pi.")
37
E.C.1.1.1.1. alkohol-dehidrogenáz
1. Oxidoreduktázok E.C alkohol-dehidrogenáz O H NAD+ NADH+H+ CH3 CH2 OH CH3 C A reakció a körülményektől függően az ellenkező irányba is folyhat. E.C tejsav dehidrogenáz Az állatokban és a mikroorganizmusokban található enzim nem azonosak, az állati szterteospecifikus, csak L-tejsavat termel, míg a mikroorganizmusokban található racém keveréket állít elő.

38
E.C. 1.6.6.1-1.6.6.3. nitrát-reduktázok
FAD tartalmú enzimek. A szervezetben a nitrátokat redukálják nitritekké. Hasonló enzimek alakítják át a húspácsók nitát tartalmának átalakítását. E.C aminosav-oxidázok Egyben dezaminálják az aminosavak.

39
Elektrontranszfer, a fontosabb elektronszállító molekulák
NAD: nikotinamid adenin-dinukleotid FAD: flavin adenin-dinukleotid Ubikinon

40
E.C. 1.1.3.4. glükóz-oxidáz: a levgő oxigénjével a glükózt közvetlenül glükonsavvá oxidálja.
Technológiai felhasználásai: - a glükóz eltávolítása - az oxigén megkötése pl.: sajtok, húsok oxidatív elszíneződését, glükóz-oxidázt tartalmazó csomagolás segítségével valósítják meg. E.C citokróm-oxidáz: terminális oxidáció a mitokondriumban E.C lipoxigenáz: Az 1-cisz, 4-cisz pentadién kettős kötéseket tartalmazó zsírsavakat oxidálja peroxidokká.

41
2. Transzferázok: E.C hexokináz E.C foszfofruktokináz

42
E.C. 2.7.5.3. foszfogliceromutáz
A foszfátcsoport eltávolítását hidrolázok a foszfoprotein foszfatázok katalizálják.

43
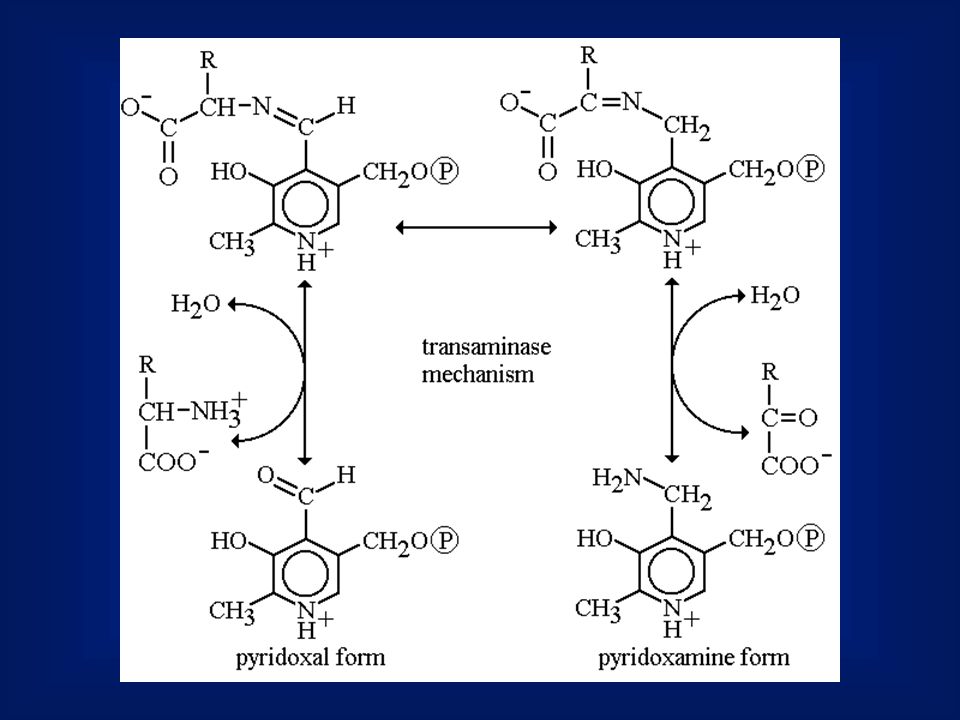
Aminotranszferázok

45
4. Liázok 3. Hidrolázok

46
5. Izomerázok 6. Ligázok

Hasonló előadás














>")




.>")
