Előadást letölteni
1
Információhordozó makromolekulák
DNS, RNS, gének, klónozás

2
Az Escherichia coli sejtet alkotó molekulák
A sejt össztömegéhez viszonyítva (%) Szárazanyag tartalom (%) Különböző molekulafélék (specieszek) száma víz 70 1 fehérjék 15 50 3000 DNS 3,3 RNS 6 20 >3000 poliszaharidok 3 10 5 lipidek 2 6,6 Monomer alegységek és intermedierek 500 Szervetlen ionok
 Szárazanyag tartalom (%) Különböző molekulafélék (specieszek) száma. víz fehérjék DNS. 3,3. RNS >3000. poliszaharidok lipidek. 2. 6,6. Monomer alegységek és intermedierek Szervetlen ionok.")
3
Az élő sejteket makromolekulák építik fel, amelyek kisebb egységek (Mr<500) polimerizációjával jönnek létre. Aminosavak → fehérjék Mr= nukleotidok → nukleinsavak Mr<109 Egyszerű cukrok → poliszaharidok Mr=106 (Glicerin+ zsírsavak → lipidek Mr= ) Nukleinsavak Információhordozó makromolekulák Fehérjék Az információáramlás iránya: a molekuláris biológia központi dogmája
 Nukleinsavak. Információhordozó makromolekulák. Fehérjék. Az információáramlás iránya: a molekuláris biológia központi dogmája.")
4
Információáramlás a makromolekulák között
Centrális dogma transzkripció replikáció transzláció fehérje prionok DNS RNS reverz transzkripció genom transzkriptóma proteóma Mindhárom típusú információhordozó makromolekula lényegében ugyanazt az információt tárolja. Miért van szükség ugyanannak az információnak három különböző alakban való megjelenésére?

5
A nukleinsavak felépítése
Enyhe lúg (enzim) Nukleotidok Erélyes lúg (enzim) Nukleozidok + H3PO4 Enzim Heterociklusosok + pentóz (purin, pirimidin) (ribóz vagy dezoxiribóz)
 Nukleotidok. Erélyes lúg (enzim) Nukleozidok + H3PO4. Enzim. Heterociklusosok + pentóz. (purin, pirimidin) (ribóz vagy dezoxiribóz)")
6
A nukleinsavak felépítésében résztvevő ötszénatomos cukrok (pentózok)
A nukleinsavak felépítésében résztvevő ötszénatomos cukrok (pentózok). A dezoxiribóz 2’ szénatomjához OH csoport helyett H kapcsolódik. Az RNS-ben ribóz, a DNS-ben dezoxiribóz taláható.
. A dezoxiribóz 2’ szénatomjához OH csoport helyett H kapcsolódik. Az RNS-ben ribóz, a DNS-ben dezoxiribóz taláható.")
7
A nukleinsavak (DNS és RNS) felépítésében résztvevő bázisok alapvegyületei: a purin és a pirimidin. Ezek a szerves bázisok heterociklusos (nitrogént tartalmazó) vegyületek.
 vegyületek..")
8
A nukleinsavakban található bázisok: adenin, guanin, citozin, uracil (RNS), timin (DNS)
, timin (DNS)")
9
A nukleinsavak építőkövei a nukleotidok
A nukleinsavak építőkövei a nukleotidok. Ezek egy heterociklusos bázisból, cukorból és foszfátból állnak.

10
A nukleinsavak polimer láncait foszfodiészter kötések tartják össze
A nukleinsavak polimer láncait foszfodiészter kötések tartják össze. A foszfátcsoport fiziológiás pH-n (7.4) negatív töltésű, ezért elektromos térben a DNS és az RNS a pozitív pólus felé vándorol. A nukleinsavaknak a cukor-foszfát lánc által kijelölt iránya van. Egy nukleinsavszekvencia, ha erre nincs más külön utasítás, mindig 5’→3’ irányt jelöl (pl. 5’ATGCGGTCATT3’).
 negatív töltésű, ezért elektromos térben a DNS és az RNS a pozitív pólus felé vándorol. A nukleinsavaknak a cukor-foszfát lánc által kijelölt iránya van. Egy nukleinsavszekvencia, ha erre nincs más külön utasítás, mindig 5’→3’ irányt jelöl (pl. 5’ATGCGGTCATT3’).")
11
A DNS és RNS láncoknak iránya van!
Konvenció: ATGC ≠ CGAT 5’→3’irány pH 7-en a foszfát csoportok ionizáltak → negatív töltés! A DNS kémiailag sokkal stabilabb mint az RNS. Az RNS lúggal könnyen hidrolizál a 2’ OH csoport jelenléte miatt (szomszédcsoport hatás).
.")
12
Az RNS kémiailag meglehetősen instabil molekula
Az RNS kémiailag meglehetősen instabil molekula. A 2’ OH csoport katalizálja a cukor-foszfát lánc lebomlását (szomszédcsoport hatás). A DNS molekula azért alkalmas a genetikai információ hosszú távú stabil tárolására, mert kémiailag sokkal ellenállóbb, mint az RNS (nincs 2’ OH).
. A DNS molekula azért alkalmas a genetikai információ hosszú távú stabil tárolására, mert kémiailag sokkal ellenállóbb, mint az RNS (nincs 2’ OH).")
13
A DNS két antiparalel láncból felépülő ún
A DNS két antiparalel láncból felépülő ún. kettős spirál szerkezetet alkot. A két láncot a bázisok közötti hidrogén kötések tartják össze. Az adenin és timin között kettő, míg a guanin és a citozin között három hidrogén kötés alakul ki. A hidrogén kötés viszonylag gyenge másodlagos kölcsönhatás, azonban nagy számban már igen komoly stabilizációs erőt jelent.

14
A DNS kettősspirál különböző megjelenési formái (konformációi)
A DNS kettősspirál különböző megjelenési formái (konformációi). A B-DNS a leggyakoribb forma.
. A B-DNS a leggyakoribb forma.")
15
Az RNS-nek, szemben a DNS-sel, bonyolult háromdimenziós szerkezete lehet. Az RNS ezért alkalmas szerkezeti és katalitikus funkciók ellátására is. Az ábra a 16S riboszómális RNS-t ábrázolja.

16
A DNS molekulában a tárolt információ a bázisok sorrendjében rejlik.
A bázisok sorrendjének meghatározása a DNS szekvenálás. Régen többféle módszert is alkalmaztak (pl. a kémiai lebontást → Maxam-Gilbert). Ma már kizárólag a Sanger-féle láncterminációs módszert ill. annak automatizált változatait használják. Ezzel a módszerrel határozták meg a humán genom szekvenciáját is.
. Ma már kizárólag a Sanger-féle láncterminációs módszert ill. annak automatizált változatait használják. Ezzel a módszerrel határozták meg a humán genom szekvenciáját is.")
17
A Sanger-féle szekvenálásnál használt didezoxi-nukleotid (ddNTP)
A Sanger-féle szekvenálásnál használt didezoxi-nukleotid (ddNTP). A didezoxi-nukleotidnak nem csak a 2’, hanem a 3’ szénatomjához sem kapcsolódik OH csoport. Mivel a DNS szintézis 5’→3’ irányba halad (az enzim a 3’ OH csoporthoz illeszti a következő nukleotidot), a ddNTP beépülése a DNS láncba további nukleotidok beépülését lehetetlenné teszi, vagyis a szintézis leáll, a DNS lánc csonka marad.
. A didezoxi-nukleotidnak nem csak a 2’, hanem a 3’ szénatomjához sem kapcsolódik OH csoport. Mivel a DNS szintézis 5’→3’ irányba halad (az enzim a 3’ OH csoporthoz illeszti a következő nukleotidot), a ddNTP beépülése a DNS láncba további nukleotidok beépülését lehetetlenné teszi, vagyis a szintézis leáll, a DNS lánc csonka marad.")
18
Nukleinsavak gélelektroforézise
Nukleinsavak gélelektroforézise. Elektromos térben a nukleinsavak a pozitív pólus felé vándorolnak. Ha az elektroforézis gélközegben végezzük, a gélben a kisebb molekulák gyorsabban, a nagyobbak lassabban fognak mozogni. Ezzel a módszerrel a nukleinsav molekulák nagyság szerint szétválaszthatók.

19
A Sanger-féle láncterminációs DNS szekvenálás
A Sanger-féle láncterminációs DNS szekvenálás. A szekvenálandó egyláncú DNS-t (templát) DNS-polimeráz enzimmel kettős láncúvá egészítjük ki. Szintézis közben olyan nukleotid származékokat (didezoxi nukleotidok, ddNTP) adunk az elegyhez, amelyek megakadályozzák a lánc tovább épülését, ha beépülnek a DNS láncba. Megfelelő koncentráció arányok beállításával elérhetjük, hogy minden lehetséges fragmentum keletkezzék. Ezután a keletkezett DNS molekulákat gélelektroforézis segítségével nagyság szerint szétválasztjuk (növekvő sorrendbe állítjuk őket). A gél aljától fölfelé haladva leolvasható a szekvencia (az eredeti DNS lánc komplementer szekvenciája).
 DNS-polimeráz enzimmel kettős láncúvá egészítjük ki. Szintézis közben olyan nukleotid származékokat (didezoxi nukleotidok, ddNTP) adunk az elegyhez, amelyek megakadályozzák a lánc tovább épülését, ha beépülnek a DNS láncba. Megfelelő koncentráció arányok beállításával elérhetjük, hogy minden lehetséges fragmentum keletkezzék. Ezután a keletkezett DNS molekulákat gélelektroforézis segítségével nagyság szerint szétválasztjuk (növekvő sorrendbe állítjuk őket). A gél aljától fölfelé haladva leolvasható a szekvencia (az eredeti DNS lánc komplementer szekvenciája).")
20
Radioaktívan jelölt DNS molekulák szétválasztása poliakrilamid gélelektroforézissel. A gélen alulról fölfelé haladva a szekvencia leolvasható.

21
Automata DNS szekvenátor működési elve

22
A GÉN FOGALMA Morgan, XX. század eleje: A gén a kromoszóma egy része (darabja), amely meghatározza az élőlény egy tulajdonságát (fenotípus). Beadle és Tatum, 1940: egy gén - egy enzim hipotézis egy gén – egy fehérje hipotézis egy gén – egy polipeptid Avery, 1944: A gének anyaga DNS. Mai definíció: A gén egy olyan DNS szakasz, amely egy géntermék (polipeptid vagy RNS) szintéziséhez szükséges információt tárolja. A szűken vett definíció csak a struktúrgént jelenti (polipeptid vagy RNS elsődleges szekvenciáját kódoló DNS), a tágabb definícióba beleértjük a regulátor szekvenciákat (promóterek, enhancerek, stb.) is.
 szintéziséhez szükséges információt tárolja. A szűken vett definíció csak a struktúrgént jelenti (polipeptid vagy RNS elsődleges szekvenciáját kódoló DNS), a tágabb definícióba beleértjük a regulátor szekvenciákat (promóterek, enhancerek, stb.) is.")
23
A gén Stop kodon TAA, TAG, TGA Start kodon ATG (Met)
5’ „nemkódoló” szakasz promóter, enhancer, riboszómakötőhely, stb Srtuktúrgén ORF: open reading frame Eukarióták esetén intronokat is tartalmaz 3’ „nemkódoló” szakasz poliadeniláció, transzkripciós terminátor, stb.

24
RNS gének Vannak gének, amelyek olyan RNS-ek szekvenciáját kódolják, amelyek nem fordítódnak le fehérjévé. Riboszómális RNS (rRNS): A legintenzívebben átíródó gének közé tartoznak minden szervezetben (nucleolus = sejtmagvacska). Transzfer RNS (tRNS): A fehérjeszintézishez (transzláció) nélkülözhetetlenek. Kis nukleáris RNS (snRNS): RNS molekulák „érése” (splicing) Kis nukleoláris RNS (snoRNS): nt, rRNS processzálás, alternatív splicing, telomeráz RNS, stb. Mikro RNS (miRNS): ~22 nt, Hosszabb prekurzorokból keletkeznek, génkifejeződést szabályozzák: RNS interferencia (Orvosi Nóbel díj, 2006) Ezeknek a géneknek a felépítése jelentősen eltér a fehérjét kódoló gének felépítésétől, ezért sokkal nehezebb őket megtalálni a genomban. Pl. az miRNS géneket csak néhány éve fedezték fel!
: A legintenzívebben átíródó gének közé tartoznak minden szervezetben (nucleolus = sejtmagvacska). Transzfer RNS (tRNS): A fehérjeszintézishez (transzláció) nélkülözhetetlenek. Kis nukleáris RNS (snRNS): RNS molekulák „érése (splicing) Kis nukleoláris RNS (snoRNS): nt, rRNS processzálás, alternatív splicing, telomeráz RNS, stb. Mikro RNS (miRNS): ~22 nt, Hosszabb prekurzorokból keletkeznek, génkifejeződést szabályozzák: RNS interferencia (Orvosi Nóbel díj, 2006) Ezeknek a géneknek a felépítése jelentősen eltér a fehérjét kódoló gének felépítésétől, ezért sokkal nehezebb őket megtalálni a genomban. Pl. az miRNS géneket csak néhány éve fedezték fel!")
25
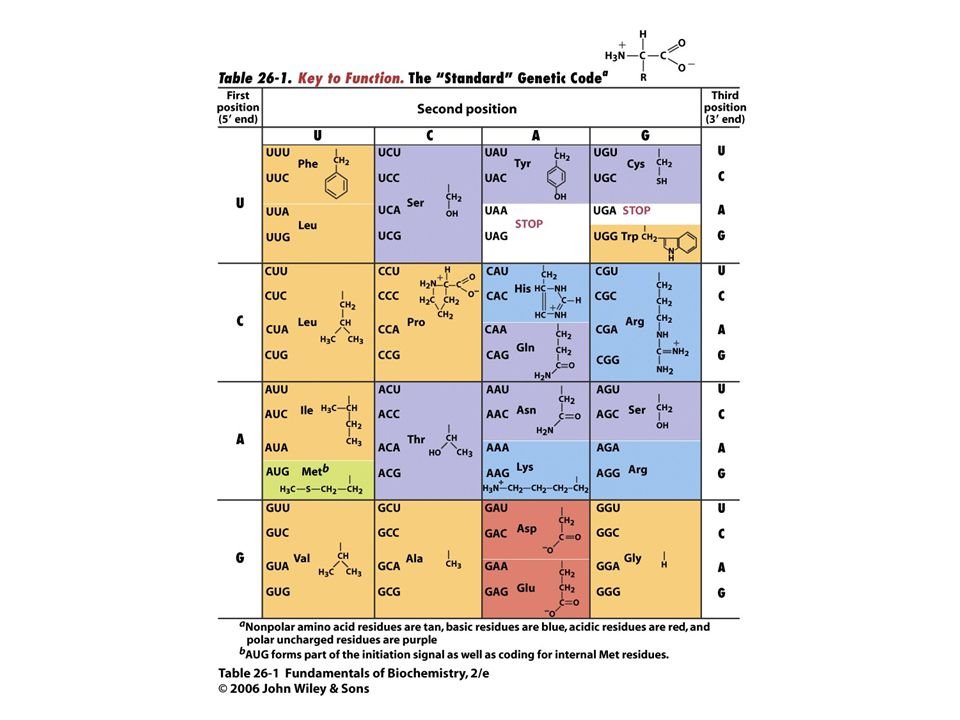
A DNS-ben 4 bázis (A,T,G,C) kódolja az információt.
Az RNS-ben szintén (A,U,G.C). Három bázis (kódon) felel meg egy aminosavnak a fehérjeszintézis során. Genetikai kód A fehérjéket 20-féle aminosav alkotja. A fehérjék elsődleges szerkezetében (szekvenciájában) kódolva van a háromdimenziós szerkezetük. A kód mibenléte nagyrészt ismeretlen.
. Három bázis (kódon) felel meg egy aminosavnak a fehérjeszintézis során. Genetikai kód. A fehérjéket 20-féle aminosav alkotja. A fehérjék elsődleges szerkezetében (szekvenciájában) kódolva van a háromdimenziós szerkezetük. A kód mibenléte nagyrészt ismeretlen.")
27
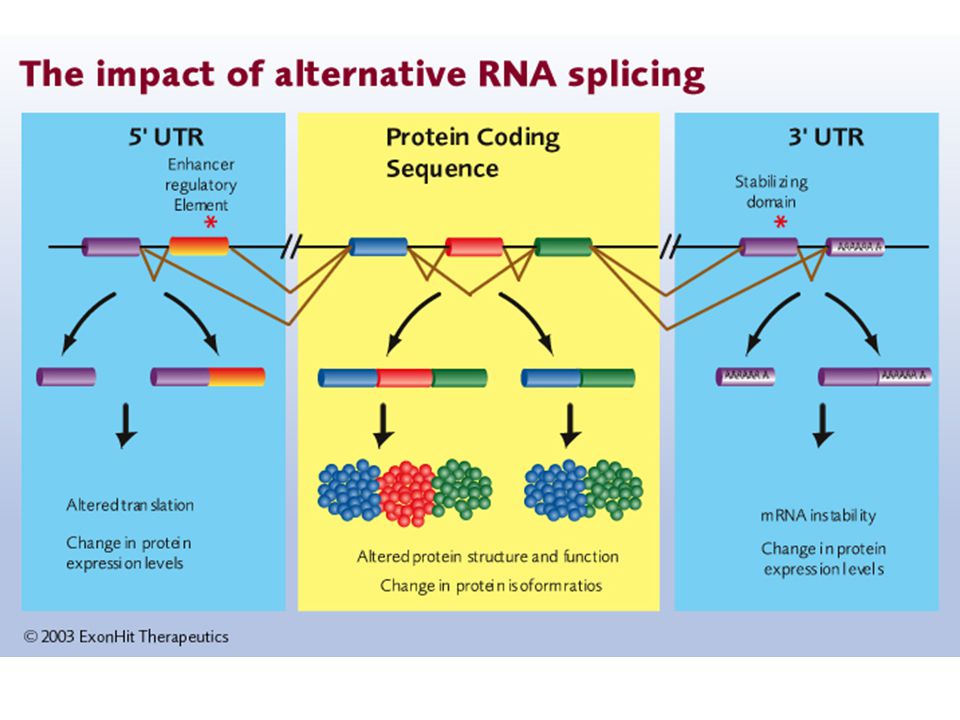
Az eukarióta génekben a fehérjét kódoló szakaszokat (exonok) nemkódoló DNS szakaszok (intronok) szakítják meg. Az intronok hossza többszöröse is lehet az exonok hosszának. Az intronok az mRNS érése során vágódnak ki. Ezt a folyamatot nevezzük „splicing”-nak. A splicing a biológiai diverzitás egyik forrása (alternatív splicing).
..")
28
Splicing a β globin gén kifejeződése során

29
A splicing mechanizmusa

31
RNS szerkesztés / RNA editing
Apolipoprotein B 100 (513 kDa) Apolipoprotein B (250 kDa) Az mRNS közepén egy stop kódon keletkezik A transzláció félúton leáll. citozin dezamináz CAA UAA Gln Stop
 Apolipoprotein B 48 (250 kDa) Az mRNS közepén egy stop kódon keletkezik. A transzláció félúton leáll. citozin. dezamináz. CAA UAA. Gln. Stop.")
32
Genom Egy élőlény teljes genetikai állománya (össz. DNS tartalom)
Pl. ember: 23 (22+2) kromoszóma + mitokondriális DNS A különböző élőlények genomjai szerkezetükben és információtartalmukban jelentősen eltérhetnek egymástól. 1.) Méret (kbp=1000bp, Mbp=106bp) 2.) Gének száma (génsűrűség= gének száma/genom mérete) 3.) Génszerkezet (intron-exon) 4.) Topológia (lineáris vs. cirkuláris)
 kromoszóma + mitokondriális DNS. A különböző élőlények genomjai szerkezetükben és információtartalmukban jelentősen eltérhetnek egymástól. 1.) Méret (kbp=1000bp, Mbp=106bp) 2.) Gének száma (génsűrűség= gének száma/genom mérete) 3.) Génszerkezet (intron-exon) 4.) Topológia (lineáris vs. cirkuláris)")
33
Az információáramlás a makromolekulák között (különösen eukarióták esetén) nagyfokú diverzitás forrása Ember: kb gén genom transzkripció alternatív splicing mRNS RNS editing transzláció Fehérje poszttranszlációs módosítás Bonyolult anyagcsere hálózat Több mint egymillió különböző géntermék proteóma

34
Az eukarióta genom felépítése
1.) Gének és szabályozó elemek: exonok és intronok transzkripciós szabályozó elemek (promóter, enhancer, terminátor, stb.) replikációt szabályozó elemek (replikációs kezdőpont) transzlációt szabályozó elemek (start, stop kodon) rekombinációs szekvenciák 2.) Ismétlődő (repetitív szekvenciák): highly repetitive sequences simple sequence DNA centroméra, teloméra satellite DNA Az egér kromoszóma 10%-a. Kevesebb mint 10 bp ismétlődik több milliószor. moderately repetitive transzpozonok (Alu repeat) Az egér kromoszóma 20%-a. Néhány száz bázispár, néhány ezerszer ismétlődik.
 Gének és szabályozó elemek: exonok és intronok. transzkripciós szabályozó elemek (promóter, enhancer, terminátor, stb.) replikációt szabályozó elemek (replikációs kezdőpont) transzlációt szabályozó elemek (start, stop kodon) rekombinációs szekvenciák. 2.) Ismétlődő (repetitív szekvenciák): highly repetitive sequences. simple sequence DNA centroméra, teloméra. satellite DNA. Az egér kromoszóma 10%-a. Kevesebb mint 10 bp ismétlődik több milliószor. moderately repetitive transzpozonok (Alu repeat) Az egér kromoszóma 20%-a. Néhány száz bázispár, néhány ezerszer ismétlődik.")
36
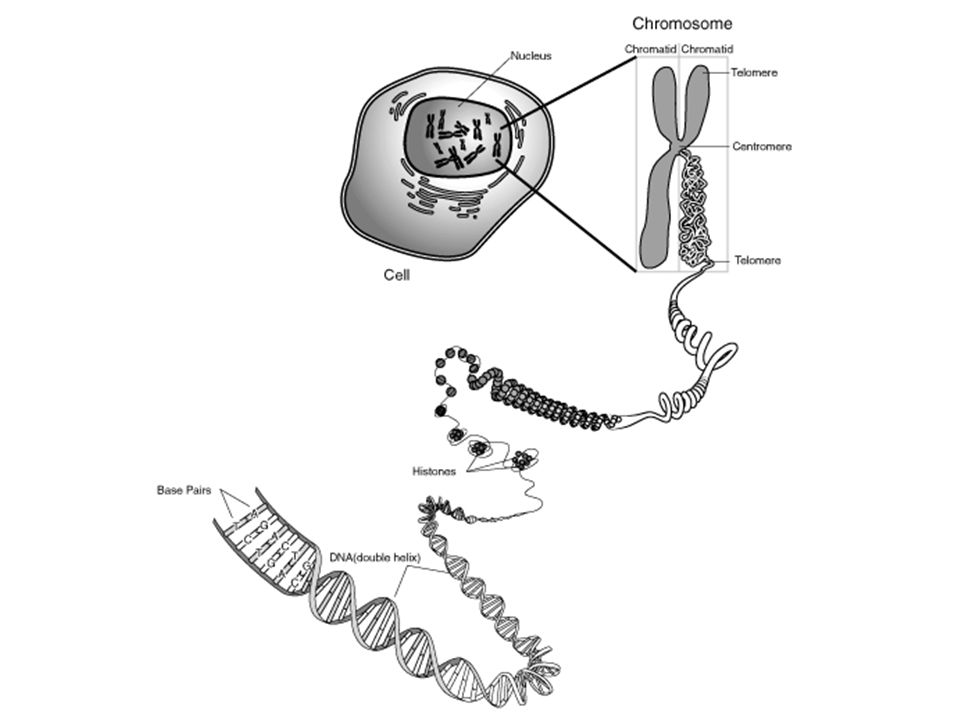
Az eukarióta sejtek kromoszómáiban a DNS molekulák bázikus fehérjék, a hisztonok köré vannak feltekeredve (nukleoszóma).
.")
37
A hisztonok köré tekeredett DNS molekulák (nukleoszómák) magasabb szervezettségi fokú srtuktúrákat (pl. szolenoid) alkotnak. A DNS molekulák ily módon szervezetten, „tömörítve” helyezkednek el a sejtmagon belül a kromoszómákban. Ez a tömörítés teszi lehetővé, hogy pl. egy emberi sejtben lévő 2m hosszú DNS a kromoszómákon mindössze csak 200 µm helyet foglaljon el.
 alkotnak. A DNS molekulák ily módon szervezetten, „tömörítve helyezkednek el a sejtmagon belül a kromoszómákban. Ez a tömörítés teszi lehetővé, hogy pl. egy emberi sejtben lévő 2m hosszú DNS a kromoszómákon mindössze csak 200 µm helyet foglaljon el..")
38
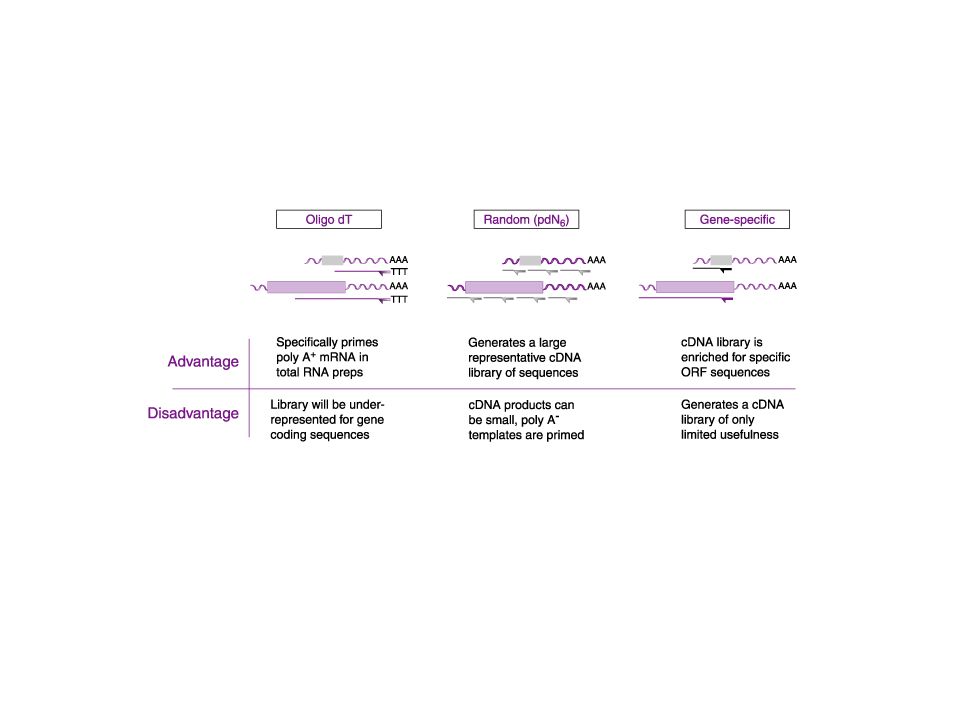
A cDNS fogalma Mivel az eukarióta mRNS már nem tartalmazza az intronokat, szekvenciája közvetlenül megfeleltethető az általa kódolt fehérje szekvenciájával. Az RNS azonban instabil, bomlékony, klónozási célokra nem alkalmazható. Megoldás: Az RNS molekulából készítsünk DNS-t reverz transzkriptáz enzim segítségével. Az így készült DNS kópiát nevezzük c(opy)DNS-nek. A cDNS alkalmas baktériumsejtekben való rekombináns fehérjetermeltetésre is. Reverz transzkriptáz: Virális eredetű enzim → retrovírusok Emberben is van reverz transzkripció → telomeráz
DNS-nek. A cDNS alkalmas baktériumsejtekben való rekombináns fehérjetermeltetésre is. Reverz transzkriptáz: Virális eredetű enzim → retrovírusok. Emberben is van reverz transzkripció → telomeráz.")
41
Probléma: Hogyan tudunk megkeresni, azonosítani és vizsgálni egy DNS szakaszt (gént) a rendkívül nagy (pl. humán 3X109 bp) és komplex genomban? (Tű a szénakazalban.) Megoldás: DNS klónozás, vagyis egy kiválasztott DNS szakasz izolálása és megsokszorozása akár több milliárd példányban. Módszer: Rekombináns DNS technika ( génsebészet, genetic engineering). A biokémia, molekuláris genetika (molekuláris biológia) legnagyobb hatású felfedezése a 20. században. Rekombináns DNS nélkül ma már elképzelhetetlen a kutatás és a biotechnológiai ipar.
 Megoldás: DNS klónozás, vagyis egy kiválasztott DNS szakasz izolálása és megsokszorozása akár több milliárd példányban. Módszer: Rekombináns DNS technika ( génsebészet, genetic engineering). A biokémia, molekuláris genetika (molekuláris biológia) legnagyobb hatású felfedezése a 20. században. Rekombináns DNS nélkül ma már elképzelhetetlen a kutatás és a biotechnológiai ipar.")
42
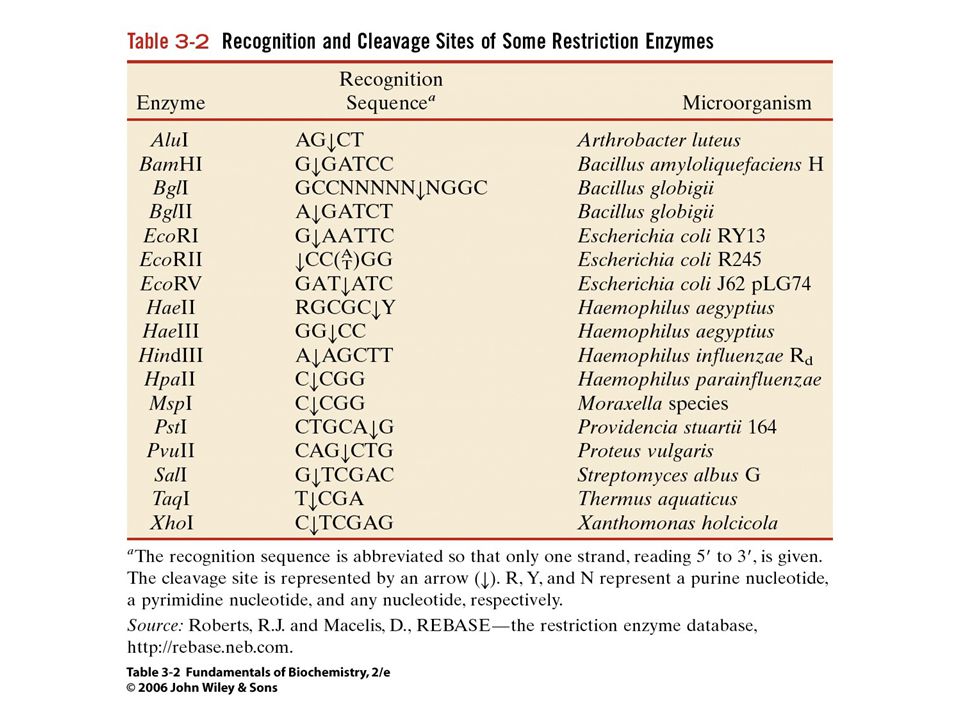
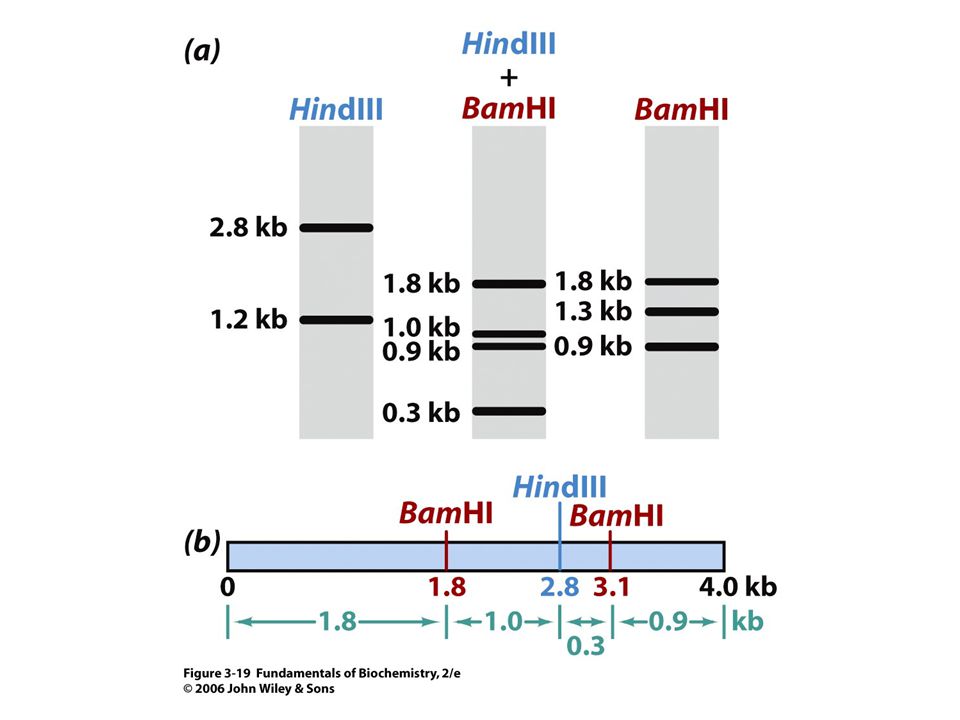
A DNS klónozása Génsebészet (genetic engineering): a DNS manipulálása (vágás/illesztés) speciális enzimekkel. Restrikciós endonukleázok: Bakteriális eredetű enzimek. Egy adott DNS szekvenciát – ált. 4-6 bázis hosszúságú – ismernek fel a kettősszálú DNS molekulán belül és elhasítják azt. Több ezer különböző restrikciós endonukleázt ismerünk, amelyek több mint száz DNS szekvenciát ismernek fel. Elkészíthetjük a DNS restrikciós (fizikai) térképét. A restrikciós fragmentumokat mesterséges hordozó (vektor) DNS-be ültetjük (ligáz enzim) Rekombináns DNS A rekombináns DNS-t megfelelő gazdaszervezetben (pl. E. coli baktérium) több millió kópiában megsokszorozhatjuk Klón A rekombináns klón elegendő mennyiségű anyagot szolgáltat a DNS analízisére (pl. szekvenálás).
 térképét. A restrikciós fragmentumokat mesterséges hordozó (vektor) DNS-be ültetjük (ligáz enzim). Rekombináns DNS. A rekombináns DNS-t megfelelő gazdaszervezetben (pl. E. coli baktérium) több millió kópiában megsokszorozhatjuk. Klón. A rekombináns klón elegendő mennyiségű anyagot szolgáltat a DNS analízisére (pl. szekvenálás).")
43
Restrikciós endonukleázok

44
A restrikciós endonukleázok biológiai funkciója
A baktériumok „immunrendszere”. Minden endonukleáznak megvan a maga metiláz enzim párja. A metilálás megvédi a DNS-t az endonukleáz hasításától. Csak a II-es típusú endonukleáz-metiláz rendszerben válik szét két külön fehérjére a két funkció. Ezért csak a II-es típusú enzimeket használják a génsebészetben.

47
A DNS-ligáz enzim 5’foszfát és 3’OH végeket kapcsol össze ATP hasítása közben. DNS ligázzal bármilyen eredetű DNS darabok összekapcsolhatók. Rekombináns DNS: különböző biológiai eredetű DNS darabok összekapcsolásával keletkezett új DNS molekula.

48
A DNS klónozás sémája A rekombináns DNS-t gazdaszervezetbe juttatva megsokszorozzuk.

49
Escherichia coli K12-es törzs, a leggyakrabban használt gazdaszervezet a génsebészetben
Gram-negatív baktérium, molekuláris szinten a legjobban jellemzett élőlény. Kiválóan alkalmas rekombináns DNS molekulák megsokszorozására. A leggyakrabban alkalmazott vektorok: Plazmidok, bakteriofágok.

50
Plazmidok: Extrakromoszómális genetikai elemek a baktériumban
Plazmidok: Extrakromoszómális genetikai elemek a baktériumban. Kis méretű (1-200kb), cirkulárisan zárt, dupla szálú DNS molekulák, amelyek általában szuperhelikális konformációban vannak. A plazmidok önálló, a kromoszómától független replikációra képesek. A replikációs origó határozza meg a kópiaszámot, ami néhánytól néhány százig terjedhet. A plazmidok a legrégebbóta használt klónozó vektorok.
, cirkulárisan zárt, dupla szálú DNS molekulák, amelyek általában szuperhelikális konformációban vannak. A plazmidok önálló, a kromoszómától független replikációra képesek. A replikációs origó határozza meg a kópiaszámot, ami néhánytól néhány százig terjedhet. A plazmidok a legrégebbóta használt klónozó vektorok.")
51
A pBR322-es plazmid, az egyik legelső E. coli vektor

52
A modern klónozó plazmidok számos praktikus „kényelmi” elemet tartalmaznak.

53
Bakteriofágok: a baktériumok vírusai.
Mivel önállóan képesek replikálódni a sejten belül és tolerálják az idegen DNS-t, alkalmasak klónozó vektoroknak. Előnyük a plazmidokhoz képest: 1.)Természetes úton képesek bejutni a sejtbe, ezért a transzformáció sokkal hatékonyabb. 2.) Lényegesen nagyobb DNS inszertek befogadására képes (8-24 kb/λ-fág), ezért alkalmasabb klóntárak készítésére. Hátrány: Körülményesebb, nehézkesebb a kezelésük a laboratóriumban. A két leggyakrabban használt klónozó fág: λ-fág és M13 fág
Természetes úton képesek bejutni a sejtbe, ezért a transzformáció sokkal hatékonyabb. 2.) Lényegesen nagyobb DNS inszertek befogadására képes (8-24 kb/λ-fág), ezért alkalmasabb klóntárak készítésére. Hátrány: Körülményesebb, nehézkesebb a kezelésük a laboratóriumban. A két leggyakrabban használt klónozó fág: λ-fág és M13 fág.")
54
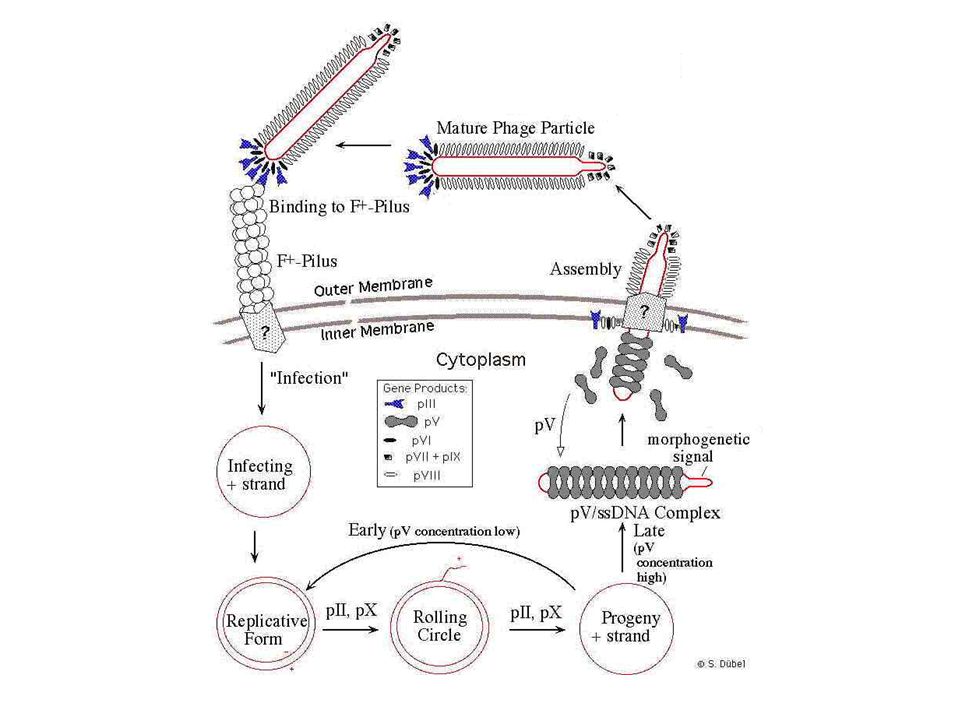
A λ-fág életciklusa

55
Klónozás λ-fág vektorba

56
M13 fág Kétféle célból használják a génsebészetben:
1.) Egyes szálú (ssDNS) előállítására. 2.) Fág bemutatásra (phage display). Kezelése egyszerűbb, mint a λ-fágé.
 Egyes szálú (ssDNS) előállítására. 2.) Fág bemutatásra (phage display). Kezelése egyszerűbb, mint a λ-fágé.")
58
Egyéb vektorok: pl. kozmidok, mesterséges kromoszómák
Ezek nagyméretű DNS darabok klónozására alkalmasak. Kozmid kb BAC (bacterial artificial chromosome) kb YAC (yeast artificial chromosome) kb
 kb. YAC (yeast artificial chromosome) kb.")
59
Egy adott szekvenciát (gént) hordozó klón kiválasztása kolónia/plakk hibridizációval
Ez a módszer volt az egyedüli a gének klónozására a nyolcvanas évek közepéig, amikor felfedezték a polimeráz-láncreakciót (PCR).
.")
60
Polimeráz láncreakció (PCR)
Vektor és gazdatörzs (baktérium) használata nélkül megsokszorozhatjuk (amplifikálhatjuk) a DNS-t. A sejtmagban végbemenő DNS replikáció in vitro imitálása. Tetszés szerinti DNS szakasz megsokszorozható. A megsokszorozandó DNS-t határoló rövid szakaszok szekvenciáját ismerni kell.
 használata nélkül megsokszorozhatjuk (amplifikálhatjuk) a DNS-t. A sejtmagban végbemenő DNS replikáció in vitro imitálása. Tetszés szerinti DNS szakasz megsokszorozható. A megsokszorozandó DNS-t határoló rövid szakaszok szekvenciáját ismerni kell.")
61
Polimeráz láncreakció

62
A PCR hőmérséklet profilja

63
A PCR-hez termostabil DNS-polimeráz enzim szükséges

64
Primer tervezés: az egyik legkritikusabb lépés a PCR során.
Szempontok: hossz, olvadáspont, kereszthibridizáció, másodlagos szerkezeti elemek a DNS-ben, stb. Ma már számos számítógép program áll rendelkezésre primer tervezéshez. PCR egyetlen igazi hátránya: ismernünk kell a DNS szekvenciáját, amit amplifikálni akarunk.



 DNS-ből,>")












 kialakulása Genetikai, Sejt- és Immunbiológiai Intézet Falus András.>")


.>")
>")

