Előadást letölteni
Az előadás letöltése folymat van. Kérjük, várjon

1
Glutation és a kapcsolódó mechanizmusok Fitokelatinok, metallotioneinek

2
A nehézfémfelesleg hatásai
Más elemek gyökér általi felvételének befolyásolása (Cd, Cu, Zn) Szabad oxigéngyökök generálása Cu+ + O2 → Cu2+ + O2.- Cu+ +H2O2 → Cu2+ + OH- + OH. lipidperoxidáció iniciálása Cu+ + LOOH → Cu2+ + OH- + LO. Cu2+ + LOOH → Cu+ + H+ + LOO. Enzimek működésének gátlása (pl. NR-t az Pb) Klorofill bioszintézis gátlása (δ-aminolevulinsav DH, protoklorofillid reduktáz)
 Szabad oxigéngyökök generálása. Cu+ + O2 → Cu2+ + O2.- Cu+ +H2O2 → Cu2+ + OH- + OH. lipidperoxidáció iniciálása. Cu+ + LOOH → Cu2+ + OH- + LO. Cu2+ + LOOH → Cu+ + H+ + LOO. Enzimek működésének gátlása (pl. NR-t az Pb) Klorofill bioszintézis gátlása. (δ-aminolevulinsav DH, protoklorofillid reduktáz)")
3
A nehézfémfelesleg hatásai
A nehézfém-toxicitás általános tünetei gyökér- (hajtás-) növekedésének gátlása vízfelvétel- és belső vízmozgás gátlása Különösen magasabb koncentrációban a legtöbb nehézfém: gátolja sztómák nyitódását és a transzspirációt CO2-fixáció, a fotoszintézis egyes folyamatainak gátlása károsítja a plazmamembránt
 növekedésének gátlása. vízfelvétel- és belső vízmozgás gátlása. Különösen magasabb koncentrációban a legtöbb nehézfém: gátolja sztómák nyitódását és a transzspirációt. CO2-fixáció, a fotoszintézis egyes folyamatainak gátlása. károsítja a plazmamembránt.")
4
A nehézfémrezisztencia lehetséges mechanizmusai
Elkerülés (baktériumban P-típusú ATPázok kipumpálják) Detoxifikálás a sejten belül (kelatálás, transzport) - fehérjék indukciója hősokk (Heat-shock) proteinek, metallotioneinek, fitokelatinok, ferritin, transzferrin - antioxidáns enzimek indukciója és aktiválása szuperoxid dizmutáz, kataláz, glutation peroxidáz - szerves savak magasabb „turnover”-e malát, citrát, oxalát, fitát, szukcinát stb. A nehézfémtolerancia fontos a nehézfém hiperakkumuláláshoz !
 Detoxifikálás a sejten belül (kelatálás, transzport) - fehérjék indukciója. hősokk (Heat-shock) proteinek, metallotioneinek, fitokelatinok, ferritin, transzferrin. - antioxidáns enzimek indukciója és aktiválása. szuperoxid dizmutáz, kataláz, glutation peroxidáz. - szerves savak magasabb „turnover -e. malát, citrát, oxalát, fitát, szukcinát stb. A nehézfémtolerancia fontos a nehézfém hiperakkumuláláshoz !")
5
A Cd kelatálásában és vakuólumba juttatásában központi szerepe van a glutationnak

6
Antioxidáns védő mechanizmusok

7
Az aszkorbinsav-glutation ciklus szerepe a H2O2 eltávolítása

8
A glutation

9
A glutation szerepe a növényekben
Előfordulása: kloroplasztiszban,citoszólban, mitokondriumban mM-os nagyságrendben (500 µM - 5 mM) Redukált/oxidált glutation aránya: normál körülmények között: % stresszhatásra: 30-80%
 Redukált/oxidált glutation aránya: normál körülmények között: % stresszhatásra: 30-80%")
10
Redukált glutation – oxidált glutation
2 GSH → GSSG Oxidált glutation = glutation diszulfid

11
A glutation szerepe a növényekben
Oxidatív stressz elleni védelem (GR, GSPX) Szubsztrátja a glutation S-transzferáznak Redox puffer, redox szabályozás (fehérjék -SH csoportja, sejtciklus) Génexpresszió szabályozása Fitokelatinok prekurzora Fő raktározott formája a redukált kénnek A szervek közötti kénmegoszlás szabályozása
 Szubsztrátja a glutation S-transzferáznak. Redox puffer, redox szabályozás (fehérjék -SH csoportja, sejtciklus) Génexpresszió szabályozása. Fitokelatinok prekurzora. Fő raktározott formája a redukált kénnek. A szervek közötti kénmegoszlás szabályozása.")
12
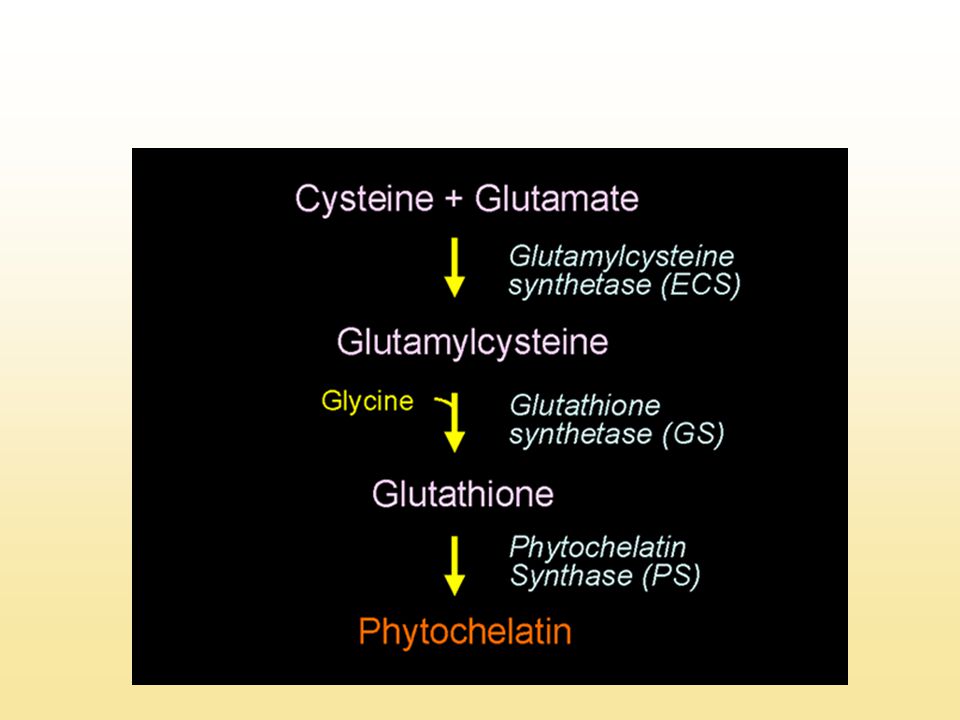
A glutation bioszintézise

13
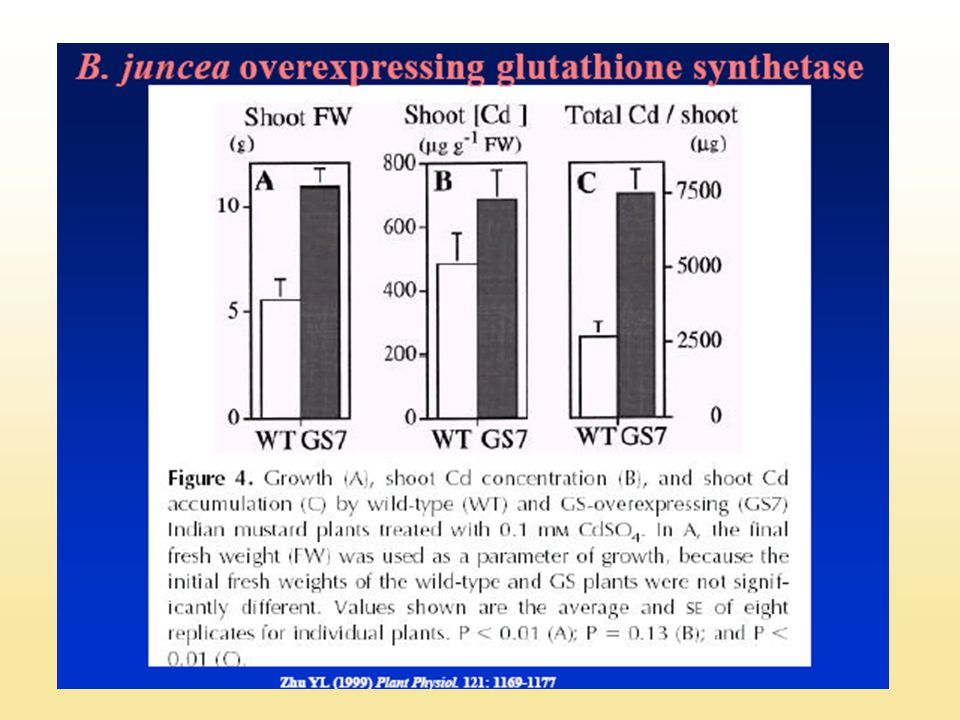
A glutation szintetáz, GS (GSase)
Transzgénikus nyárfa Citoszolban, 100-szoros GS-növekedés, GSH tartalom nem nőtt Transzgénikus Brassica j. : 2-3-szoros növekedés a Cd-toleranciában

15
g-glutamil cisztein szintetáz, g-ECS
Szerepe bizonyított a GSH szint emelkedésben stresszhatásra Transzgénikus nyárfa: citoszolban: enzim mennyiségben 5-10-szeres növekedés, 2-3-szoros GSH- és GSSG- szint növekedés (a redox állapot nem különbözött a kontrolltól !) Kloroplasztiszban: hasonló eredmény Transzgénikus Brassica j. : 2-3-szoros növekedés a Cd-toleranciában Transzgénikus dohány: nekrózis, csökkent növekedés
 Kloroplasztiszban: hasonló eredmény. Transzgénikus Brassica j. : 2-3-szoros növekedés a Cd-toleranciában. Transzgénikus dohány: nekrózis, csökkent növekedés.")
16
A bioszintézis génjének túltermeltetésével a glutation mennyisége megemelhető növényekben

17
Glutation reduktáz, GR Transzgénikus nyárfa
kloroplasztiszban: 1000-szeres GR-növekedés 2-szeres növekedés a glutationban (aszkorbát mennyisége is nőtt stresszhatásra) citoszolban:5-10- szeres GR növekedés glutation mennyisége nem nőtt GR overexpresszió a kloroplasztiszban növeli a GSH/GSSG arányt és a totál glutation mennyiséget is Transzgénikus dohány: 1-3,5-szörös GR növekedés
 citoszolban:5-10- szeres GR növekedés. glutation mennyisége nem nőtt. GR overexpresszió a kloroplasztiszban növeli a. GSH/GSSG arányt és a totál glutation mennyiséget is. Transzgénikus dohány: 1-3,5-szörös GR növekedés.")
18
Glutation bioszintézis és homeosztázis összetett szabályozása

19
A glutation szintézise, felhasználása és lebontása a növényi sejtben
GSH: redukált glutation, Glu: glutaminsav g-ECS: g-glutamil cisztein szintetáz, GSase: glutation szintetáz, GSSG: glutation diszulfid, GRase: glutation reduktáz, GST: glutation S-transzferáz, PCS: fitokelatin szintetáz, CPase: karboxipeptidáz, g-GTase: g-glutamil transzpeptidáz, DPase: dipeptidáz

20
A glutation S-transferáz (GST) szerepe
Különböző elektrofil komponensek konjugálása GSH-val (endogén metabolitok, gyomirtószerek detoxifikálása ) GSH-PX funkció→ lipid peroxidok detoxifikálása GST/GS-PX gént túltermelő transzgénikus dohány növények megemelkedett stressz toleranciával rendelkeznek izomerizációs reakciók katalizátora (pl. aromás aminosavak lebontásában) nem-enzimatikus szállító (ligandin-funkció): endogén molekulák kötése nem-katalitikus helyen (kötés és transzport) - auxin- és citokinin-kötő fehérjék - cianidin-3-glukozid vakuolumba történő transzportja - intracelluláris hormon transzport? Apoptózis szabályozása - véd a letális hatástól („Bax-inhibitor GST”)
 GSH-PX funkció→ lipid peroxidok detoxifikálása. GST/GS-PX gént túltermelő transzgénikus dohány növények megemelkedett stressz toleranciával rendelkeznek. izomerizációs reakciók katalizátora (pl. aromás aminosavak lebontásában) nem-enzimatikus szállító (ligandin-funkció): endogén molekulák kötése nem-katalitikus helyen (kötés és transzport) - auxin- és citokinin-kötő fehérjék. - cianidin-3-glukozid vakuolumba történő transzportja. - intracelluláris hormon transzport Apoptózis szabályozása - véd a letális hatástól. („Bax-inhibitor GST )")
21
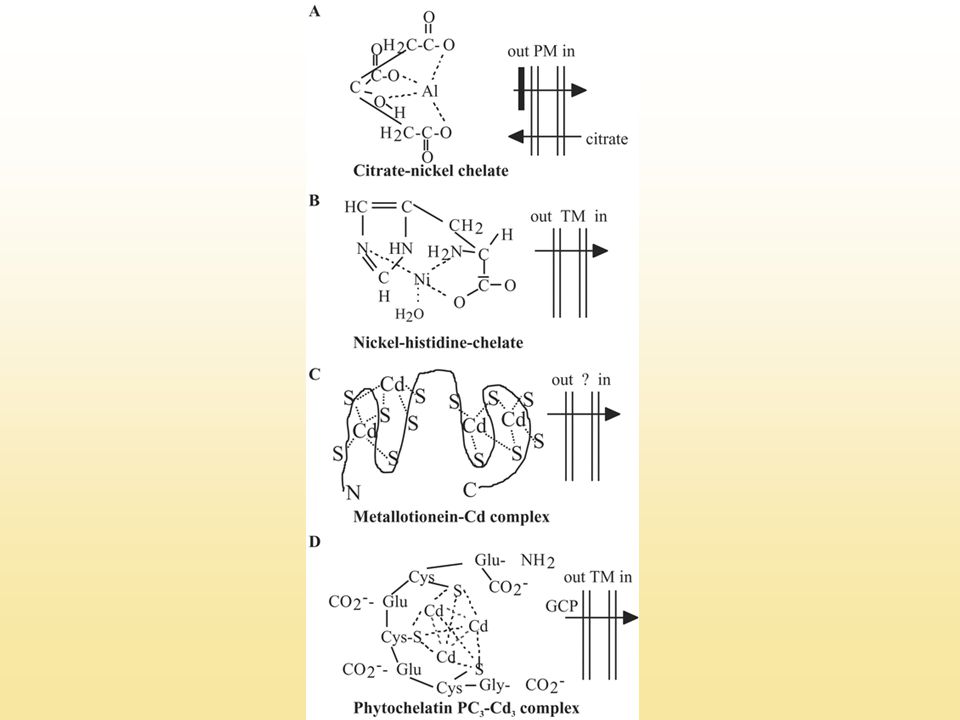
A fitokelatinok Fémkötő polipeptidek (γ-Glu-Cys)2-11-Gly a felesleges, toxikus fémionok megkötése, kizárása, elkülönítése

22
A fitokelatinok

24
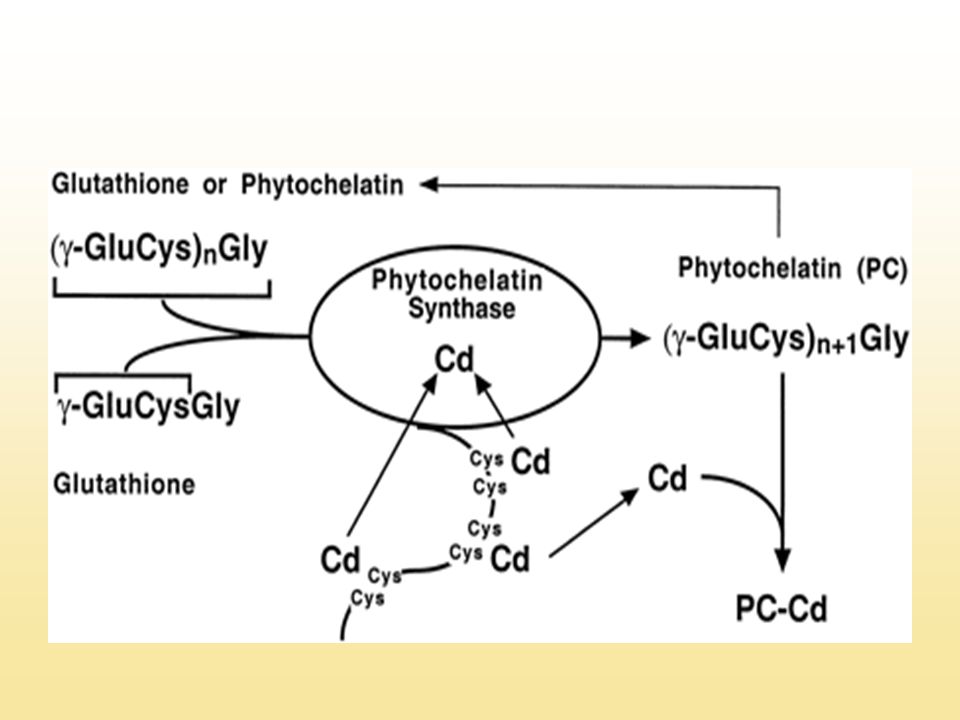
A fitokelatin szintáz (PCS)
Nehézfémek és tiolátjaik aktiválják (Cd2+, Zn2+ ,Cd.GS2, Zn.GS2) C-terminális részen Cys-csoportok fontosak az aktiválásban cad1 mutánsok (Arabidopsis, S. pombe) mutáció a PC szintázban - Cd2+ -ra, Cu2+-ra hiperszenzitívek - fitokelatin-deficiensek PCS-expresszáló transzgénikus sejtek Cd2+-toleranciája és -akkumulációja, nagyobb, mint a kontrollé (S. pombe, S. cerevisiae, dohány, Arabidopsis)
 C-terminális részen Cys-csoportok fontosak az aktiválásban. cad1 mutánsok (Arabidopsis, S. pombe) mutáció a PC szintázban. - Cd2+ -ra, Cu2+-ra hiperszenzitívek. - fitokelatin-deficiensek. PCS-expresszáló transzgénikus sejtek Cd2+-toleranciája és -akkumulációja, nagyobb, mint a kontrollé. (S. pombe, S. cerevisiae, dohány, Arabidopsis)")
26
Cd2+ kapcsolódása két fitokelatin molekulával
A fitokelatinok Cd2+ kapcsolódása két fitokelatin molekulával

27
A fitokelatinok

29
A fitokelatinok A Cd2+-ot jól toleráló sejtekben a Cd2+ min. 90%-a Cd-fitokelatin komplex formájában található A Cd-fitokelatin komplexek általában a növényi sejtek vakuólumában találhatóak Transzporterük: ABC-transzporter (HMT1: heavy metal tolerance factor 1)
")
30
A vakuólum a növényi sejtek 90%-át is kitöltheti

31
Fitokelatin-közvetített Cd-transzport és Cd-S komplexek kialakulásának modellje

33
A kén csökkenti a Cd toxicitását

34
A metallotioneinek Kis molekulasúlyú, ciszteinben gazdag fémkötő proteinek (8-14 kDa, aminosav, >30% Cys) „fitokelatin aggregátumok” ? Fitokelatinokhoz hasonló szerep a fémek komplexálásában (nagyobb affinitás a Cu-hez) Antioxidáns funkciójuk is lehet (membránok védelme) Génexpressziójukat befolyásolják pl. nehézfémek (Cu, Zn, Cd), sebzés, öregedés, patogén fertőzés.
 Antioxidáns funkciójuk is lehet (membránok védelme) Génexpressziójukat befolyásolják pl. nehézfémek (Cu, Zn, Cd), sebzés, öregedés, patogén fertőzés.")
35
S. cerevisie metallotionein
A metallotionein molekula szerkezete, kiemelve a Cys csoportok S atomjait (világosabb gömbök ) és a kötött Cu atomokat (sötétebb gömbök)
 és a kötött Cu atomokat (sötétebb gömbök)")
36
A metallotioneinek csoportosítása aminosav sorrend és összetétel alapján
I. típus: 20 Cys-t tartalmaznak (6 Cys-X-Cys két fő doménban, köztük kb. 40 aminosav) II. típus: N- és C-terminális részen más, konzervált elrendezésben vannak az aminosavak (pl. Cys-Cys, Cys-Glu-Glu-Cys, MSCCGGNCGCS) tipikusan növényi MT III. típus: N-terminális részen csak 4 Cys, C-terminálison 6 enzimatikusan, nem transzlációsan szintetizálódnak! IV. típus: 3 Cys-gazdag domént tartalmaznak (5-6 Cys), köztük aminosav)
 II. típus: N- és C-terminális részen más, konzervált elrendezésben vannak az aminosavak (pl. Cys-Cys, Cys-Glu-Glu-Cys, MSCCGGNCGCS) tipikusan növényi MT. III. típus: N-terminális részen csak 4 Cys, C-terminálison 6. enzimatikusan, nem transzlációsan szintetizálódnak! IV. típus: 3 Cys-gazdag domént tartalmaznak (5-6 Cys), köztük aminosav)")
37
Egy metallotionein feltételezett szerkezete
Az N- ás C-terminális részen lévő Cd- (kék gömbök) kötő fém-centrumokat hosszabb fehérjelánc köti össze.
 kötő fém-centrumokat. hosszabb fehérjelánc köti össze.")
38
A metallotioneinek Génexpressziójuk különbözhet szövettől függően is:
I. típus gyökérben jobban expresszálódik, mint hajtásban, II. típus ellenkezőleg III. típus expressziója magas levelekben és éretlen termésekben IV. típus fejlődő magvakban expresszálódik. Nyárfában 6 MT expresszióját vizsgálták, eltérő mintázat

39
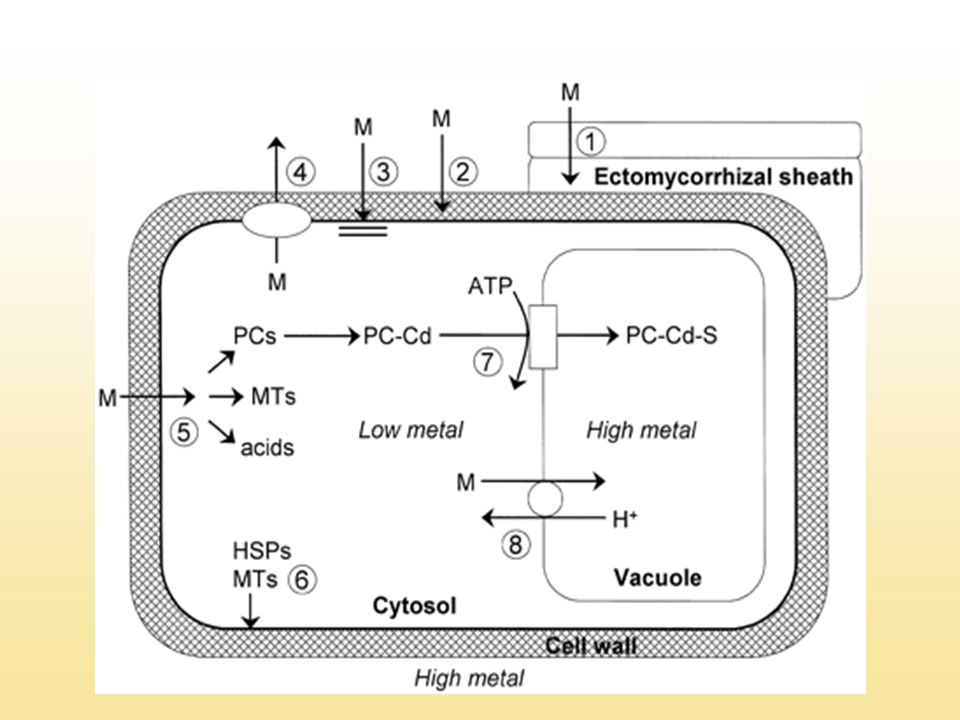
A nehézfémrezisztencia lehetséges mechanizmusai
Fémionok immobilizálása a sejtfalban (pektinek) PM permeabilitás változása, kompartmentizáció a vakuólumban Aktív kijuttatás Kelátképzés a citoplazmában, vakuoláris transzport
 PM permeabilitás változása, kompartmentizáció a vakuólumban. Aktív kijuttatás. Kelátképzés a citoplazmában, vakuoláris transzport.")
40
Lehetőségek a növények nehézfém rezisztenciájának, akkumulációjának, transzportjának növelésére

Hasonló előadás





 megváltozHAT Ebből visszakövetkeztethetünk a mutációt szenvedett gén funkciójára.>")













