Mikrobasejtek ciklus alatti növekedése A tenyészet sejtszáma az idő függvényében N(t) = N 0 ·e ·t (ha a külső környezet és a sejtek fiziológiai állapota változatlan és a populáció aszinkron) A tenyészet sejttömege az idő függvényében M(t) = M 0 ·e ·t (a sejtek koreloszlása és ciklus alatti tömegnövekedése ezt nem befolyásolja) Az egyedi sejt tömege az idő függvényében m(t) = m 0 ·e ·t ??? m 0 a születéskori sejttömeg 0 < t < CT, ahol CT a sejt ciklusideje m(CT) = 2m 0 (tetszőleges monoton függvény elképzelhető) exponenciális-e a növekedés a sejtek életében ???
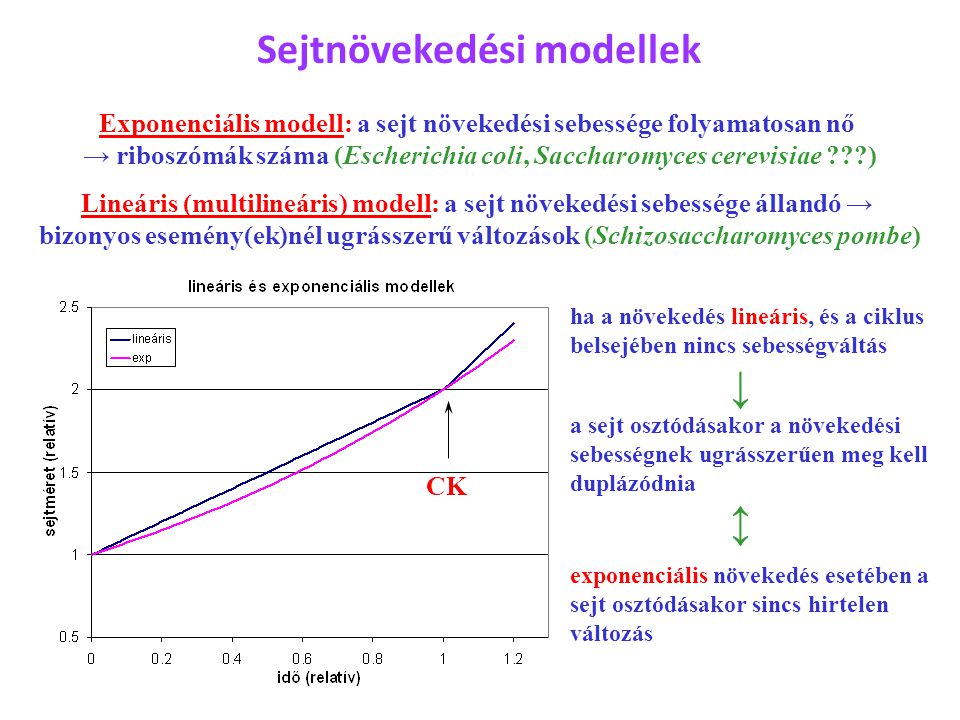
Sejtnövekedési modellek Exponenciális modell: a sejt növekedési sebessége folyamatosan nő → riboszómák száma (Escherichia coli, Saccharomyces cerevisiae ???) Lineáris (multilineáris) modell: a sejt növekedési sebessége állandó → bizonyos esemény(ek)nél ugrásszerű változások (Schizosaccharomyces pombe) CK ha a növekedés lineáris, és a ciklus belsejében nincs sebességváltás ↓ a sejt osztódásakor a növekedési sebességnek ugrásszerűen meg kell duplázódnia ↕ exponenciális növekedés esetében a sejt osztódásakor sincs hirtelen változás
A hasadó élesztőgomba, mint a sejtnövekedés modellje Mitózisos (ivartalan) sejtciklus Mitchison, 1957 csak hosszirányú sejtnövekedés → tömeg, térfogat és hossz ~ arányos a ciklus végén a hossznövekedés megszűnik Mitchison & Nurse, 1985 Sveiczer, Novák & Mitchison, 1996 a növekedés mintázata (bi)lineáris a ciklus közepe táján van egy sebességváltási pont (RCP)
A hasadó élesztőgomba mitózisos sejtciklusa
SG2MG1S RCP2 Idő (min) Sejthossz, L ( m) NETO új régi vég CK A Schizosaccharomyces pombe növekedése RCP3 RCP1
DL BL CT 5 min A mikrofotográfia módszere mért változó: a sejthossz (L) a születéstől (BL) az osztódásig (DL) az idő függvényében (0 < t < CT)
A mérési pontokra illesztett modellek Lineáris L(t) = a·t + b Exponenciális L(t) = c·e d·t Bilineáris L(t) = ·ln{exp[ ·(t- )/ ] + exp[ ·(t- )/ ]} + ahol az RCP pozíciója, pedig az átmenet élessége A legadekvátabb modell kiválasztási kritériumai Korrelációs koefficiens Reziduális standard deviációs = (SSE/df) 1/2 Akaike információs kritérium AIC = n obs · ln(SSE) + 2n par Schwarz Bayes információs kritérium SBIC = n obs · ln(SSE) + n par · ln(n obs )
RCP α 1 = μ m min -1 α 2 = μ m min -1 ε = 9.84 μ m τ = 57.6 min η = 0.01 μ m Horváth, A., Rácz-Mónus, A., Buchwald, P. & Sveiczer, A. (2013). Cell length growth in fission yeast: an analysis of its bilinear character and the nature of its rate change transition. FEMS Yeast Res. 13: Egy hasadó élesztő sejt növekedési mintázata (éles bilineáris eset)
RCP α 1 = μ m min -1 α 2 = μ m min -1 ε = 10.2 μ m τ = 59.7 min η = μ m Horváth, A., Rácz-Mónus, A., Buchwald, P. & Sveiczer, A. (2013). Cell length growth in fission yeast: an analysis of its bilinear character and the nature of its rate change transition. FEMS Yeast Res. 13: Egy hasadó élesztő sejt növekedési mintázata (sima bilineáris eset)
γ = μ m min -1 δ = 7.86 μ m Horváth, A., Rácz-Mónus, A., Buchwald, P. & Sveiczer, A. (2013). Cell length growth in fission yeast: an analysis of its bilinear character and the nature of its rate change transition. FEMS Yeast Res. 13: Egy hasadó élesztő sejt növekedési mintázata (lineáris eset)
α 1 = μ m min -1 α 2 = μ m min -1 ε = 10.8 μ m τ = 64.3 min η = μ m RCP Horváth, A., Rácz-Mónus, A., Buchwald, P. & Sveiczer, A. (2013). Cell length growth in fission yeast: an analysis of its bilinear character and the nature of its rate change transition. FEMS Yeast Res. 13: Hasadó élesztő sejtek „átlagos” növekedési mintázata (sima bilineáris)
A hasadó élesztő sejtnövekedése általában bilineáris, de gyakran nem éles
A méretkontroll igazolása hasadó élesztőben 1. WT cdc2 ts Fantes, P. A. (1977). Control of cell size and cycle time in Schizosaccharomyces pombe. J. Cell Sci. 24,
Sveiczer, A., Novak, B. & Mitchison, J. M. (1996). The size control of fission yeast revisited. J. Cell Sci. 109, A méretkontroll igazolása hasadó élesztőben 2.
Sveiczer, A., Novak, B. & Mitchison, J. M. (1996). The size control of fission yeast revisited. J. Cell Sci. 109, A méretkontroll a G2 fázis közepén hat → G2 = „sizer + timer” A méretkontroll pozícionálása hasadó élesztőben