Előadást letölteni
Az előadás letöltése folymat van. Kérjük, várjon

1
1.) Magas csoportátviteli potenciálú vegyületek egymásba általában szabadon átalakulnak, mert a termék és reaktáns koncentrációarány változhat úgy a sejtben, hogy a szabadentalpiaváltozás negatív legyen, a folyamat spontán végbemenjen. 2.) Alacsony csoportátviteli potenciálú vegyületek egymásba általában szabadon átalakulhatnak, mert a termék és reaktáns koncentrációarány szintén változhat eleget. 3. ) A magas csoportátviteli potenciálú vegyületek bomlása, amely exergonikus folyamat, fedezi az energiáját egy alacsony csoportátviteli potenciálú vegyület endergonikus szintézisének. Ezek a kapcsolt reakciók. 4.) Alacsony csoportátviteli potenciálú vegyületről csoport nem kerülhet át egy szubsztrátra, hogy magas csoportátviteli potenciálú vegyület keletkezzen. 5.) Természetesen a magas csoportátviteli potenciálú vegyületeknek is keletkezniük kell valahogy, ha léteznek. Kétféle folyamat létezik, amelyben létrejönnek: szubsztrátszintű foszforiláció: 2 lépés a glikolízisben és egy a citrátkörben oxidatív foszforiláció a mitokondriumban
 Alacsony csoportátviteli potenciálú vegyületek egymásba általában szabadon átalakulhatnak, mert a termék és reaktáns koncentrációarány szintén változhat eleget. 3. ) A magas csoportátviteli potenciálú vegyületek bomlása, amely exergonikus folyamat, fedezi az energiáját egy alacsony csoportátviteli potenciálú vegyület endergonikus szintézisének. Ezek a kapcsolt reakciók. 4.) Alacsony csoportátviteli potenciálú vegyületről csoport nem kerülhet át egy szubsztrátra, hogy magas csoportátviteli potenciálú vegyület keletkezzen. 5.) Természetesen a magas csoportátviteli potenciálú vegyületeknek is keletkezniük kell valahogy, ha léteznek. Kétféle folyamat létezik, amelyben létrejönnek: szubsztrátszintű foszforiláció: 2 lépés a glikolízisben és egy a citrátkörben oxidatív foszforiláció a mitokondriumban.")
2
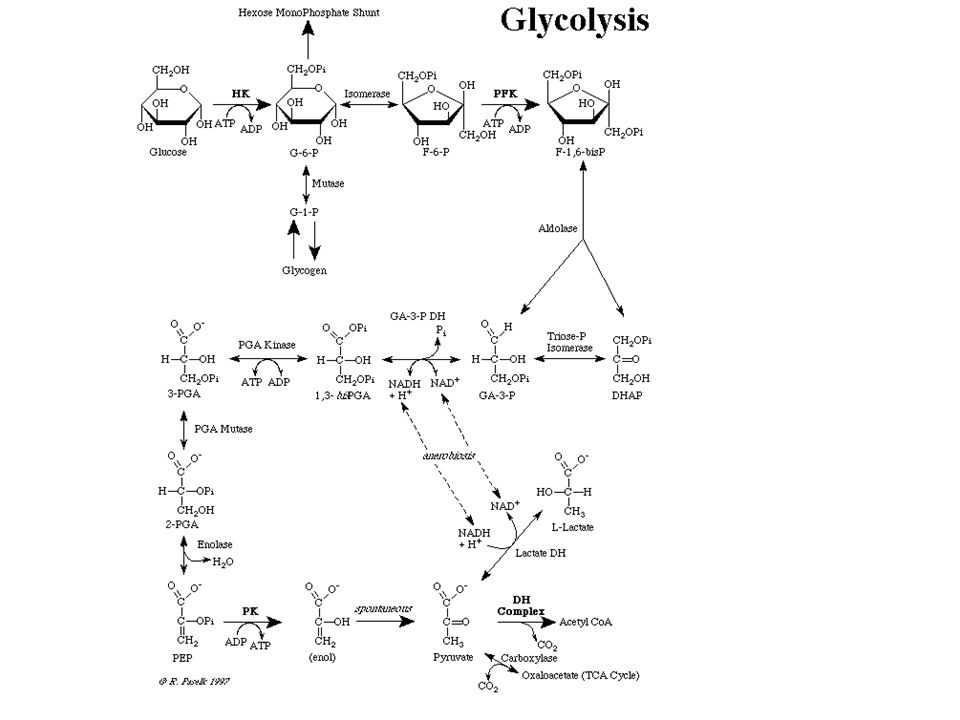
Szubsztrátszintű foszforiláció a glikolízisben

3
glykys = édes görögül lysis = bomlás görögül glikolízis = édes bomlása, vagyis egy cukor, a szőlőcukor degradációja Anyagcserefolyamatok tanulásakor felteendő kérdések: hol? = milyen élőlény mely sejtjének melyik organellumában? miért? = mi a célja a folyamatnak, mi termelődik, változik? mikor? = milyen fiziológiás vagy patológiás körülmények között zajlik? hogyan szabályozódik ennek megfelelően? hogyan? = mi az egyes reakciók mechanizmusa?

4
Lokalizáció: a glikolízis minden élőlény, tehát az ember minden élő sejtjének
citoplazmájában zajlik, semmilyen sejtorganellum nem kell hozzá. Energiatermelő folyamat, a szőlőcukor lebontásának egyetlen olyan, univerzális folyamata, amely ATP szintézisét eredményezi oxigén nélkül is. Történhet oxigén nélkül, anaerob körülmények között, ekkor a végtermék tejsav = laktát, és lezajlik oxigén jelenlétében, aerob körülmények között, ekkor piruvát a végtermék. Közvetlenül az oxigén nem vesz részt a folyamatban. Akkor zajlik a glikolízis, ha a sejtnek energiára van szüksége, vagy ezen kívül a terméket, a piruvátot, valaminek a szintézise érdekében tovább alakítja. Létezik sejt szintű energetikai szabályozás, gyors allosztérikus mechanizmussal. Ez a Pasteur-effektus. Az ember egyes szerveiben hormonális szabályozás biztosítja az összhangot a szervek között, a glukózhomeosztázis fenntartására.

5
Click on a reaction to show the enzymes mechanism
6
A glikolízis jellemzői
A kiindulási vegyület és a végtermékek nem foszforiláltak, minden köztitermék egy vagy két helyen foszforilált. 6 szénatomos hexózok és 3 szénatomos cukrok, valamint propionsavszármazékok vesznek részt benne. Résztvevő enzimek típusai: kinázok = molekulák közti foszfátcsoport átvivők , a 4 enzimből 3 irreverzibilis lépést katalizál, csak ezek a megfordíthatatlan lépések, és ezek a szabályozott enzimek izomerázok: aldo-keto cukorátalakítók, molekulán belüli csoportátvivő mutáz liázok: 6 szénatomos vegyületet ketté hasít az egyik, vizet eliminál a másik dehidrogenázok: 2 H-atomot von el az egyik és NAD-ra teszi, a másik az előbbi reakcióban keletkezett NADH-ról ad két H-atomot egy molekulának Két oxidációs lépésben az oxidáció energiáját felhasználva magas csoportátviteli potenciálú vegyület keletkezik, amelyekről a foszfát aztán ADP-re kerülve ATP szintetizálódhat. Ez a szubsztrátszintű foszforiláció.

7
Energetics of glycolysis:
1 glucose + 2 ADP + 2 P + 2 NAD = 2 pyruvate + 2 ATP + 2 NADH + 2 H+ if oxygen is present = aerobic condition NADH is oxidized back by shuttles/by respiratory chain of mitochondria. 1 glucose + 2 ADP + 2 P = 2 lactate + 2 ATP in anaeric condition, without O2 NADH is oxidized back by lactate dehydrogenase in the cytosol to produce lactate from pyruvate and NAD needs for glyceraldehyde-3P oxidation

8
A glikolízis szabályozása
hexokináz I, II, III izoenzimek: mindenhol van valamelyik, kivéve, ahol glukokináz van allosztérikus gátló: glukóz-6P termék glukokináz = hexokináz IV izoenzim: májban, pancreasban fruktóz-6P-ot kötött reguláló fehérje gátolja PFK-1 = foszfofruktokináz-1 szabályozása májban, szívben, izomban, vesekéregben: allosztérikus gátlók: ATP, citrát,P-kreatin, (2,3-BPG, PEP) ― mindegyik csak energetikailag jól feltöltött sejtben magas koncentrációjú, hosszú szénláncú zsírsavak, ketontestek ― alternatív tápanyag H+, K+, Mg2+ ― pathológiás állapotokban allosztérikus aktiválók: AMP, ADP,P, ― energiahiány esetén magas koncentrációjúak (fruktóz-6P, fruktóz-1,6P2) ― glukóz van a sejtben fruktóz-2,6-biszfoszfát ― hormonális szabályozás alatt áll a szintje
 ― mindegyik csak energetikailag jól feltöltött sejtben magas koncentrációjú, hosszú szénláncú zsírsavak, ketontestek ― alternatív tápanyag. H+, K+, Mg2+ ― pathológiás állapotokban. allosztérikus aktiválók: AMP, ADP,P, ― energiahiány esetén magas koncentrációjúak. (fruktóz-6P, fruktóz-1,6P2) ― glukóz van a sejtben. fruktóz-2,6-biszfoszfát ― hormonális szabályozás alatt áll a szintje.")
9
Pasteur-effektus Louis Pasteur 1860-ban felfedezte, hogy az élesztőgombák anaerob körülmények között sokkal több szőlőcukrot bontanak le, mint oxigén jelenlétében, és oxigén nélkül sok tejsav = laktát keletkezik, míg aerob körülmények között alig. A glukózlebontás sebességmeghatározó lépése a PFK-1 által katalizált reakció. Oxigén nélkül, az ősi körülmények között csak a glikolízis termeli az energiát, vagyis 2 ATP-t glukózonként. A sok AMP serkenti az enzimet. 2 ADP ↔ ATP + AMP jobbra tolódik Aerob körülmények között, (a megjelenő légkörben) a glikolízis után folytatódik a szőlőcukor lebomlása egészen CO2-ig és H2O-ig, további, összesen max ATP-t eredményezve. A sok ATP gátolja az enzimet, AMP ekkor szinte nincs jelen.
 a glikolízis után folytatódik a szőlőcukor lebomlása egészen CO2-ig és H2O-ig, további, összesen max ATP-t eredményezve. A sok ATP gátolja az enzimet, AMP ekkor szinte nincs jelen.")
10
Pasteur-effektus 2. 1. ) Aerob körülmények között a glikolízisben a végtermék piruvát, a keletkező NADH visszaoxidálódik a citoplazmában egy „inga” segítségével, H-je bejut a mitokondriumba, és ott ATP szintézisét teszi lehetővé. 2.) Anaerob körülmények között a glukózlebontás mitokondriális folyamatai nem működnek, a NADH H-je nem ingázik, ezért kell lennie a citoplazmában egy reakciónak, amely visszaoxidálja, hogy a glicerinaldehid-3P-deh. folyamatosan tudjon dolgozni. Ez a laktát-dehidrogenáz által katalizált reakció, így a végtermék a piruvát továbbalakulásával keletkező laktát.
 Aerob körülmények között a glikolízisben a végtermék piruvát, a keletkező NADH visszaoxidálódik a citoplazmában egy „inga segítségével, H-je bejut a mitokondriumba, és ott ATP szintézisét teszi lehetővé. 2.) Anaerob körülmények között a glukózlebontás mitokondriális folyamatai nem működnek, a NADH H-je nem ingázik, ezért kell lennie a citoplazmában. egy reakciónak, amely visszaoxidálja, hogy a glicerinaldehid-3P-deh. folyamatosan tudjon dolgozni. Ez a laktát-dehidrogenáz által katalizált reakció, így a végtermék a piruvát továbbalakulásával keletkező laktát.")
11
Piruvátkináz szabályozása májban:
allosztérikus gátlók: ATP, alanin allosztérikus serkentő: fruktóz-1,6-biszfoszfát glukagon szignálra foszforilált enzim: inaktív inzulin szignálra defoszforilált enzim: aktív Hosszú távú szabályozás gén szinten: magas vércukorszint → inzulin → HK, PFK-1, PK indukálódnak éhezés = alacsony vércukorszint → glukagon → GK, PFK-1, PK represszálódnak

12
Click on a reaction to show the enzymes mechanism
14
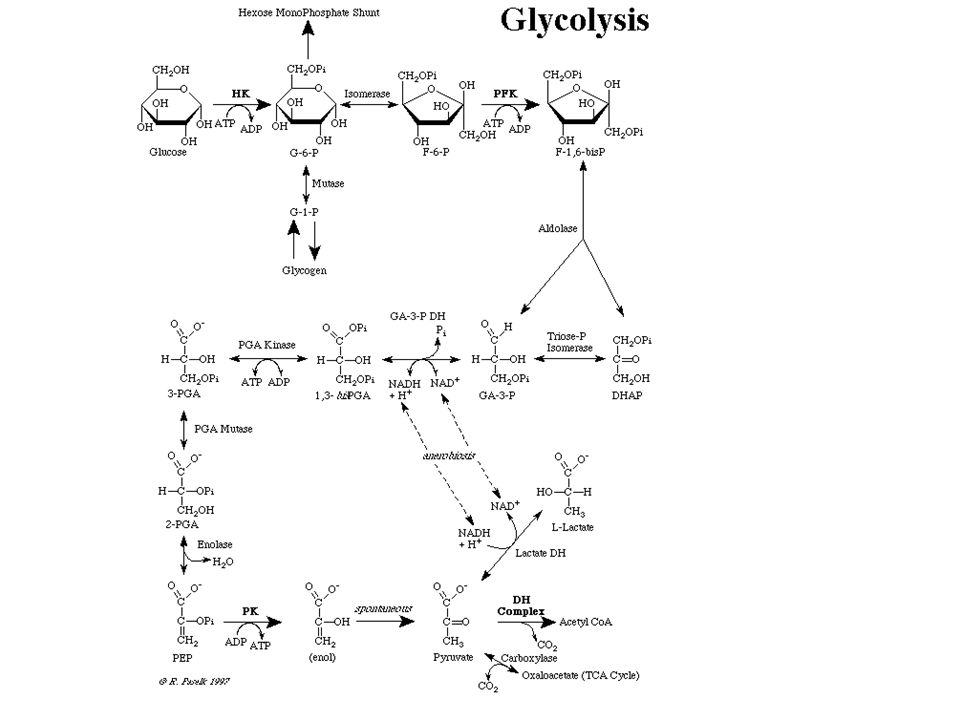
Foszfoglicerát-mutáz = phosphoglycerate mutase

15
enoláz = enolase

16
piruvátkináz = pyruvate kinase
logikátlan az enzim neve, mert a piruvát a termék, és sosem a szubsztrátja az enzimnek, lévén a reakció irreverzibilis az enol-oxo tautomerizáció spontán zajlik, az oxigén nagyon nagy elektronegativitása miatt, és 30 kJ/mol körüli energiafelszabadulással jár

17
laktát-dehidrogenáz = lactate dehydrogenase

Hasonló előadás




















