Előadást letölteni
Az előadás letöltése folymat van. Kérjük, várjon

1
Szabályozás Transzkripció, transzláció, enzimaktivitás, fehérje koncentráció, efektor molekulák

2
Szabályozó útvonalak prokariótákban

3
A szabályozás elsődlegesen a transzkripciónál
A prokarióta gének operonokba szerveződnek A gének megnyilvánulását promóterek vezérlik

4
A DNS függő RNS polimeráz (RNAP) felismeri a promótert és elkezdi a transzkripciót
Iniciáció RNS polimeráz (core enzim) + szigma faktor (sokféle; specifikusság)
 + szigma faktor (sokféle; specifikusság)")
5
Az mRNS transzkriptum szinézise (5’3’); komplementere a templát szálnak– szigma disszociál
Elongáció

6
Az elongáció addíg folytatódik, míg az RNAP el nem éri a terminátort és disszociál

7
A transzkripció iniciáció: a RNAP holoenzim a promoterhez kapcsolódik
holoenzim = RNAP core + σ faktor

8
A vegetatív (s70) promóter felépítése
-a core promótert ismeri fel a σ faktor T T G A C A T A T A A T 17 bp spacer (43) % gyakoriság A 47 -60 UP Element DSR -35 -10 +1 Core Promoter
 % gyakoriság. A UP Element. DSR Core Promoter.")
9
Az iniciációs komplextől az elongációsig
RNAP + RNAP RNAP RNAP Zárt komplex Nyitott komplex Elongációs komplex promóter

10
A transzkripció iniciáció mechanizmusa
NTP-ék k1 k2 k3 k4 R + P RPC RPO RPI RPE Ez az átmenet a “promoter clearance” a promóter felszabadulása - kIV k-1 k-2 k-3 A KI egyensúlyi konstanssal írható le KI = RPC/(R + P) abortív transzkriptumok A nyitott komplex képződés sebességét gyakran kII konstanssal R – RNAP P – Promóter RPC – zárt komplex RPO – nyitott komplex RPI – iniciációs komplex RPE- elongációs komplex
 abortív. transzkriptumok. A nyitott komplex képződés sebességét gyakran kII konstanssal. R – RNAP. P – Promóter. RPC – zárt komplex. RPO – nyitott komplex. RPI – iniciációs komplex. RPE- elongációs komplex.")
11
A transzkripciós ciklus
Körfolyamatnak tekinthető iniciáció elongáció termináció

12
A Thermus aquaticus RNAP core enzim 3D szerkezete (ribbon diagram)
")
13
Mitől függ a gén transzkripciós aktivitása?
Promóter erősség Mennyire hasonlítanak a promóter fő részei (-10,-35 boxok és a köztük levő távolság) a konszenzus szekvenciához? - általában minél inkább, annál aktívabb a promóter - de néhány hely sokkal fontosabb, mint a többi TTGACA ---17bp---TATAAT---A konszenzus TCGACA---17bp---TATTAT---A erős promóter TCAGTT---19bp---GATAAC---A gyenge promóter
 a konszenzus szekvenciához - általában minél inkább, annál aktívabb a promóter. - de néhány hely sokkal fontosabb, mint a többi. TTGACA ---17bp---TATAAT---A konszenzus. TCGACA---17bp---TATTAT---A erős promóter. TCAGTT---19bp---GATAAC---A gyenge promóter.")
14
A fő elemeken kívüli szekvenciák befolyásolják
a promóter erősségét Néhány promóternél hosszabb –10-es szekvenciák (cisz) 2. UP elemek más promótereknél a -35-ös boksz előtti AT gazdag szekvenciák növelik a transzkripció sebességét (cisz) 3. Downstream elemek közvetlenül a transzkripciós start hely utáni szekvenciák befolyásolhatják a transzkripció iniciációjának hatékonyságát (cisz) Szabályozó fehérjék, vagy más effektor molekulák (transz)
 2. UP elemek. más promótereknél a -35-ös boksz előtti AT gazdag szekvenciák növelik a transzkripció sebességét (cisz) 3. Downstream elemek. közvetlenül a transzkripciós start hely utáni szekvenciák befolyásolhatják a transzkripció iniciációjának hatékonyságát (cisz) Szabályozó fehérjék, vagy más effektor molekulák (transz)")
15
Génszabályozás I – Transzkripció szabályozása
NTP-k k1 k2 k3 k4 R + P RPC RPO RPI RPE Ez az átmenet a „promóter tisztulása,” “promoter clearance”, a kIV konstanssal jellemezhető k-1 k-2 k-3 abortív transzkriptumok A KI egyensúlyi konstanssal jellemezhető KI = RPC/(R + P) A nyitott komplex képződés sebessége kII
 A nyitott komplex képződés sebessége kII.")
16
Szabályozó fehérjék – általában DNS-kötő fehérjék
Két lépés a szabályozott gén-megnyilvánuláshoz 1. σ faktor választék – megszabja, hogy melyik promoter legyen be-, vagy kikapcsolva. Szabályozó fehérjék – általában DNS-kötő fehérjék a. Represszorok – gátolják a transzkripciót b. Aktivátorok – növelik a transzkripciót c. Kettős hatású szabályozók – mindkettőt – a körülményektől függően

17
A transzkripció inicációjának kezdete: RNAP holoenzim kötődik a promóterhez
holoenzim = RNAP core (α2ββ’) + σ
 + σ.")
18
A vegetatív (s70) promóter felépítése
-a core promótert ismeri fel a σ faktor T T G A C A T A T A A T 17 bp spacer (43) % gyakoriság A 47 -60 UP elemek DSR -35 -10 +1 Core promóter DSR = downstream regulátor elemek
 % gyakoriság. A UP elemek. DSR Core promóter. DSR = downstream regulátor elemek.")
19
Alternatív σ faktorok Különböző szerkezetű promótereket ismernek föl – gének különböző regulonjai

20
Az különböző σ faktorral szabályozott promótereknek teljesen eltérő konszenzus szekvenciája lehet
s70 TTGACA – 17 bp – TATAATN3-6-A s CTTGAAA – 16 bp – CCCCATNTN3-10-T/A s GG – N12 – GC/T – 12bp – A

21
Egy adott pillanatban a legtöbb RNAP s70 –es, de néhány százalékban más σ faktor is van

22
A legtöbb s faktor a s70 –es családhoz tartozik és ezért a σ70 –hez hasonlóan működik
A s54 eltérő – a kötödés után egy aktívátor fehérjének kell aktiválni – kII–őt befolyásolja – RPO képződik

23
Hogyan tudja egy anti-σ faktor a diferenciális génszabályozást biztosítani?
Példa: egy anti-σ faktor az FlgM (FlgM – s28)
")
24
Transzkripciós faktorok
Általában DNS-kötő fehérjék, amelyek asszociálnak a szabályozott promóterrel és csökkentik, vagy növelik a transzkripció gyakoriságát. Represszorok és aktívátorok –a szabályozó fehérjék jó része mindkettőt tudja a körülményektől függően

25
A represszorok eltérő szabályozási mintázatokat juttathatnak érvényre
Arginin bioszintézis Laktóz degradáció Represszió Indukció

26
A represszió, vagy korepresszió mechanizmusa
Példa: arginin, a 20 aminosav egyike A baktériumok el tudják készíteni maguknak, de ha arginint adunk a táptalajhoz, nem működtetik a bioszintézis géneket (nem nyilvánulnak meg) (A fene se dolgozik, ha van kaja bőven!) Nincs arginin Kell a sejtnek A represszor nem kötődik Az arginin gének megnyilvánulnak Sok az arginin Elegendő a sejtnek Represszor + Arg kötődik Az arginin gének csendesek Arginin Represszor, korepresszor, effektor, operátor
 (A fene se dolgozik, ha van kaja bőven!) Nincs arginin. Kell a sejtnek. A represszor nem kötődik. Az arginin gének megnyilvánulnak. Sok az arginin. Elegendő a sejtnek. Represszor + Arg kötődik. Az arginin gének csendesek. Arginin. Represszor, korepresszor, effektor, operátor.")
27
Egy indukálható repressziós mechanizmus
Példa: laktóz, cukor szénforrás a baktériumok hasznosítani fogják a laktózt, ha az van a tápközegben, de nem működtetik az ehhez szükséges géneket, ha nincs (Miért gyártsuk le az enzimeket, ha nincs is laktóz?) Nincs laktóz A sejt nem hasznosíthatja A represszor kötődik a lac operátorhoz Lac enzimek nem képződnek Van laktóz Tápanyagként hasznosíthatják [Represszor + allolaktóz] leválik a DNS-ről A Lac enzimek gyártása elkezdődhet Lactose
 Nincs laktóz. A sejt nem hasznosíthatja. A represszor kötődik a lac operátorhoz. Lac enzimek nem képződnek. Van laktóz. Tápanyagként hasznosíthatják. [Represszor + allolaktóz] leválik a DNS-ről. A Lac enzimek gyártása elkezdődhet. Lactose.")
28
A transzkripciót aktíváló mechanizmus
Példa: maltóz, cukor szénforrás A baktériumok hasznosítják a maltózt, ha van, de a gének nem nyilvánulnak meg, ha nincs (Miért készítsük el a maltóz hasznosításához szükséges enzimeket, ha nincs a környezetünkben? – a lac-hoz hasonló logika) Van maltóz Tápanyagként hasznosíthatják a sejtek [Aktívátor+maltóz] kötődhet a DNS-hez Készülnek a Mal enzimek Nincs maltóz A sejtek nem Az aktívátor nem kötődhet a DNS-hez Nincs Mal enzim szintézis Gyenge promóter maltóz Aktívátor fehérje, iducer, aktívátor-kötő hely
 Van maltóz. Tápanyagként hasznosíthatják a sejtek. [Aktívátor+maltóz] kötődhet a DNS-hez. Készülnek a Mal enzimek. Nincs maltóz. A sejtek nem. Az aktívátor nem kötődhet a DNS-hez. Nincs Mal enzim szintézis. Gyenge promóter. maltóz. Aktívátor fehérje, iducer, aktívátor-kötő hely.")
29
Hova kötődnek a szabályozó fehérjék a promótereknél?
Az aktívátorok általában -30-as hely előtt (upstream) kötődnek; míg sok represszor utána (downstream), valamint a -30-as boksz előtt is kapcsolódhat a DNS-hez
 kötődnek; míg sok represszor utána (downstream), valamint a -30-as boksz előtt is kapcsolódhat a DNS-hez.")
30
Represszió, gátló mechanizmusok
Térbeli gátlás (steric hindrance) – a kötőhely átfed a promóterrel és és a represszor kötődés affinitása nagyobb, mint a RNAP-é (KI) Fehérje-fehérje kölcsönhatások – a represszor megakadályozza a RNAP kötődése utáni lépéseket (kII és kIV) RNS polimeráz bezárása – a represszor megváltoztatja a lokális DNS szerkezetet és így limitálja a kötődött RNAP produktivitását (kII és kIV) Összetett (multipartite) promóterek és DNS kihurkolás (DNA looping) – összetett represszorok kötődnek különböző helyeknél, megváltoztatva a DNS konformációját és befolyásolják a RNAP kötődését (KI)
 – a kötőhely átfed a promóterrel és és a represszor kötődés affinitása nagyobb, mint a RNAP-é (KI) Fehérje-fehérje kölcsönhatások – a represszor megakadályozza a RNAP kötődése utáni lépéseket (kII és kIV) RNS polimeráz bezárása – a represszor megváltoztatja a lokális DNS szerkezetet és így limitálja a kötődött RNAP produktivitását (kII és kIV) Összetett (multipartite) promóterek és DNS kihurkolás (DNA looping) – összetett represszorok kötődnek különböző helyeknél, megváltoztatva a DNS konformációját és befolyásolják a RNAP kötődését (KI)")
31
A l represszor térbeli gátlással akadályozza PR aktivitását
Lítikus funkciók -10 -35 -35 -10 Lizogén funkciók OR3 OR2 OR1 PRM PR

32
A legtöbb represszor sokkal komplikáltabb – a LacI represszor is
Az összetett operátorok és a DNS looping is gyakori További fehérjék (mint pl. a CAP és a CytR) is gyakran szerepelnek
 is gyakran szerepelnek.")
33
A transzkripció aktíválásának néhány mechanizmusa
Szinte mindig szükséges a RNAP-al való kontaktus Nagyon jó példa a katabolit represszió – a katabolit aktivátor fehérje (catabolite activator protein) (CAP) Ez „globális” szabályozó – több mint 100 promótert szabályoz
 (CAP) Ez „globális szabályozó – több mint 100 promótert szabályoz.")
34
II. – átfeda –35–ös régióval
A CAP a cAMP-vel kapcsolódva aktíválódik (cAMP intracelluláris koncentrációja nő, ha a glükóz elfogy) A fehérje dimerizálódik és különböző módon kötődhet a promóterekhez – és nagy mértékben meghajlítja a DNS-t (DNA bending) I. – a -35 box előtt II. – átfeda –35–ös régióval A RNAP-pal érintkezve stimulálja a transzkripciót Class I Class II
 A fehérje dimerizálódik és különböző módon kötődhet a promóterekhez – és nagy mértékben meghajlítja a DNS-t (DNA bending) I. – a -35 box előtt. II. – átfeda –35–ös régióval. A RNAP-pal érintkezve stimulálja a transzkripciót. Class I. Class II.")
35
A promóter aktíválás I. és a II csoportjának modelljei
Az I. csoportban a CAP kötőhelyek -62-től -103-ig terjedhetnek. A CAP a RNAP α-alegységének C-terminális (αCTD) részével lép kölcsönhatásba A II. csoportban a CAP kötőhelye általában átfed a -35-ös régióval. A CAP kölcsönhat az aCTD-vel, aNTD-velés a s faktorral is
 részével lép kölcsönhatásba. A II. csoportban a CAP kötőhelye általában átfed a -35-ös régióval. A CAP kölcsönhat az aCTD-vel, aNTD-velés a s faktorral is.")
36
Az AraC szükséges az ara gének aktíválásához
További adatok szerint az AraC kapcsolódik egy a PBAD promótere (az araBAD gének promótere) előtti távoli araO2 operátorhoz és ekkor gátolja a PBAD aktivitását.
 előtti távoli araO2 operátorhoz és ekkor gátolja a PBAD aktivitását.")
37
Regulátorok aktívátor és represszor aktivitással
Az arabinóz katabolizmus regulátora az AraC jó példa erre Sok gén vesz részt a felvételében és a katabolizmusában

38
Az AraC-től függő szabályozás
Összetett repressziós loop Transzkripciós aktívátor I. csoport araC-PBAD kazetta kereskedelmi forgalomban Szigorú represszió arabinóz nélkül (és glükóz jelenlétében) 2. Arabinóz hozzáadásával erős aktíválódás
 2. Arabinóz hozzáadásával erős aktíválódás.")
39
A s54 –től függő aktíválás mechanizmusa
( a jó öreg σ) Az NtrC transzkripciós aktívátorral való kölcsönhatással – egy enhancer típusú fehérje NtrC stimulálja a s54 RPO képződést – így befolyásolja kII-őt
 Az NtrC transzkripciós aktívátorral való kölcsönhatással – egy enhancer típusú fehérje. NtrC stimulálja a s54 RPO képződést – így befolyásolja kII-őt.")
40
A regulátorok a RNAP különböző komponenseivel lépnek kölcsönhatásba

41
Szabályozási útvonalak prokariótákban

42
A transzkripció terminációja a szabályozás fontos célpontja lehet

43
A transzkripció sok gén esetében a kódoló szekvenciák után fejeződik be

44
A trp mRNS upstream regiójának több lehetséges másodlagos szerkezete lehet, az egyik transzkripiós terminátor

45
A triptofán bioszintézise többszörösen szabályozott
- Egyszerű korepressziós mechanizmus a triptofánra adott válasz – a regulátor a TrpR triptofán + TrpR TrpR Genotípus trpEDCBA expresszió - Trp Trp WT +++++ -/+ trpR mutáns +++ A trp operon expressziója – trpEDCBA A trpR null mutánsban a trp gének nem működnek konstitutívan. Van valamennyi triptofán represszió.

46
A gén szekvenciájának elemzése érdekes tulajdonságot fedett fel
Ennek a szekvenciának a deléciója megszüntette a TrpR-től független repressziót

47
A transzkripció attenuációjának mechanizmusa
RNAP megpihen az 1:2 hajtű szintézise után 2a. A riboszóma elkezdi a leader transzlációját – áthalad a Trp kodonokon (sok Trp) 2b. A riboszóma elkezdi a leader transzlációját – a Trp kodonoknál megáll (kevés a Trp) 3a. A riboszóma befejezi a leader peptidet – a 3:4 hajtű kialakul 3b. Az álló riboszóma megakadályozza 1:2 kialakulását – helyette 2:3 képződik 4a. A RNAP terminációját a 3:4 hajtű kialakulása okozza 4b. A 3:4 hajtű nem tud idejében kialakulni, a RNAP átírja a lehetséges terminátor helyet
 2b. A riboszóma elkezdi a leader transzlációját – a Trp kodonoknál megáll (kevés a Trp) 3a. A riboszóma befejezi a leader peptidet – a 3:4 hajtű kialakul. 3b. Az álló riboszóma megakadályozza 1:2 kialakulását – helyette 2:3 képződik. 4a. A RNAP terminációját a 3:4 hajtű kialakulása okozza. 4b. A 3:4 hajtű nem tud idejében kialakulni, a RNAP átírja a lehetséges terminátor helyet.")
48
A transzláció attenuációja
Gyakran az antibiotikum rezisztencia géneknél – néhány antibiotikum célpontja a riboszóma. Egy egyedi mRNS szerkezeten és egy leader peptiden alapul

49
mRNS másodlagos szerkezete lefedi a riboszóma-kötő helyet
A transzláció várakozása a vezető-peptidnél megtöri a másodlagos szerkezetet és lehetővé teszi a transzlációt

50
A fehérje funkció szabályozása a stabilitásával
Proteázok befolyásolhatják a fehérje aktivitásának szabályozását

51
Példa: A sS faktor stabilitásának szabályozása
Az RssB fehérje versenyez a sS –ért és kijelöli lebontásra

52
Adaptáció környezeti változásra adott válaszként
Külső jelek Transzmembrán receptor Indirekt hatás Belső jelek Membránon áthatoló jelek Fiziológiai változás Génszabályozás

53
A környezeti jelzés egyszerű paradigmája – a két komponensű rendszer
> 30 ilyen rendszer az E. coli-ban – növényekben és gombákban is, nem csak baktériumokban 1. Környezetből jel (signal) 2. Érzékelő kináz (sensor kinase) 3. Válasz szabályozó (response regulator)
 2. Érzékelő kináz (sensor kinase) 3. Válasz szabályozó (response regulator)")
54
A két komponensű jelátvivő rendszer egyszerűsített modellje (two component-regulatory system)
Az értékelő hisztidin kináz (sensor histidine kinase) (HK) – általában transzmembrán fehérje – foszforilezi saját magát (autofoszforiláció) A válasz szabályozó (response regulator) (RR) – gyakran, de nem mindig, hat a génexpresszióra – a HK foszforilezi (phosphorelay)
 (HK) – általában transzmembrán fehérje – foszforilezi saját magát (autofoszforiláció) A válasz szabályozó (response regulator) (RR) – gyakran, de nem mindig, hat a génexpresszióra – a HK foszforilezi (phosphorelay)")
55
A foszfotranszfer reakciók alaplépései
Autofoszforiláció: HK-His + ATP HK-His~P +ADP Foszfotranszfer: HK-His~P + RR-Asp HK-HIs + RR-Asp~P Defoszforiláció: RR-Asp~P + H20RR-Asp + Pi Alternatív mechanizmusok 1. HK foszfatázok: HK-His~P + PPHK-His + Pi 2. RR foszfatázok: RR-Asp~P + PP RR-Asp + Pi

56
Változatos fizikai és kémiai stimulusokra válasz lehetőség
Az alap téma változatos variációi léteznek és minél többet vizsgáljuk, annál több permutációját figyelhetjük meg.

57
A kétkomponensű RR-ek szerkezetének vizsgálata szerint jól konzerválódott modul rendszerű felépítés
Che B metilészteráz NarL RR CheA hisztidin kináz

58
Hogyan reagálnak a megfelelő szignálra?
E tekintetben mind különböző A szenzor azon része, ami lehetővé teszi a szignálra adott választ nagyon különbözik az egyes fehérjékben

59
Egy jól ismert szignál transzdukciós rendszer
(N2-kötés) L A Rhizobium FixL-FixJ rendszer szabályozza a gének válaszát a molekuláris oxigénre A FixL észleli a hem csoportjához kötődő oxigént a sejt belsejében A FixJ~P aktíválja a célgéneket, ha kicsi az O2 koncentráció O2 Hem P His J J P Asp
 L. A Rhizobium FixL-FixJ rendszer szabályozza a gének válaszát a molekuláris oxigénre. A FixL észleli a hem csoportjához kötődő oxigént a sejt belsejében. A FixJ~P aktíválja a. célgéneket, ha kicsi az O2. koncentráció. O2. Hem. P. His. J. J. P. Asp.")
60
Az RssB is egy RR típusú fehérje
HK, ha van egyáltalán, nincs azonosítva. A célfunkció a proteolízis.

61
A két komponens nem elég – phosphorelay, foszfát továbbítás

63
Phosphorelay szabályozza a B. subtilis spórázását

64
Egy kettős kináz, phosphorelay szabályozza a Vibrio harveyi fénytermelését

65
Eukarióta kétkomponensű rendszerek

66
Mikrobiális differenciálódás
Definíciók Két sejt különbözik egymástól abban a tekintetben, hogy bár genomjuk azonos, a szintetizálódó fehérjék különböznek. (Jacob and Monod, 1963) A differenciálódás a sejtek alakjában és funkciójában végbemenő stabil, vagy metastabil változás, amikor is a változás a sejt normális életciklusának a része. (Marty Dworkin, 1985) A prokarióta differenciálódáshoz olyan, az alaki és funkcióbeli változások szükségesek, amelyek kiemelkedő szerepet játszanak az organizmus életciklusában. (Brun and Shimkets, 2000)
 A differenciálódás a sejtek alakjában és funkciójában végbemenő stabil, vagy metastabil változás, amikor is a változás a sejt normális életciklusának a része. (Marty Dworkin, 1985) A prokarióta differenciálódáshoz olyan, az alaki és funkcióbeli változások szükségesek, amelyek kiemelkedő szerepet játszanak az organizmus életciklusában. (Brun and Shimkets, 2000)")
67
Myxobacterium-ok: a fruiting body (termő-test) képződés komplexitása.
 képződés komplexitása.")
68
A Myxobacterium-ok termő-testjei baktérium sejtek ezreinek aggregátumából állnak
érett aggregátum fiatal aggregátum

69
Caulobacter crescentus, mint a sejtdifferenciálódás modellje
Kirajzó (swarmer) sejt: szóródás - Nincs DNS replikáció - Nincs sejtosztódás - Mozgékony (motilis) Kocsányos (stalked) sejt: reprodukció - DNS replikáció - Sejtosztódás - Felülethez kapcsolva
 sejt: szóródás. - Nincs DNS replikáció. - Nincs sejtosztódás. - Mozgékony (motilis) Kocsányos (stalked) sejt: reprodukció. - DNS replikáció. - Sejtosztódás. - Felülethez kapcsolva.")
70
A Caulobacter életciklusa
kirajzás horgony szintézis Undergoes cellular differentiation--predivisional cell has two types of cell poles The holdfast appears during the swarmer to stalked cell differentiation The holdfast is made at a specific time in a specific place, but it’s unclear how this spatial and temporal regulation is achieved. composed in part of complex polysaccharides, namely N-acetylglactosamine and N-acetylglucosamine (may also have acidic components but proteolic enzymes don’t seem to have an affect the holdfast mediates attachment to substrates in the environment allowing Caulobacter’s stalked cells to live a sessile life lehorgonyzás

71
Az úszó-sejt differenciálódás komplex programja

72
Az endospóra képződés mint fejlődési modell
Spóraképző baktériumok (pl. Bacillus, Clostridium, Myxococcus) A baktérium sejtben képződik, ezért „endo”-spóra A baktérium sejtciklus nyugvó fázisa Képződése rendszerint valamilyen környezeti hatásra, mint az éhezés, indukálódik Nagyon ellenálló - meleg, hideg, kiszáradás, UV sugárzás spórák
 A baktérium sejtben képződik, ezért „endo -spóra. A baktérium sejtciklus nyugvó fázisa. Képződése rendszerint valamilyen környezeti hatásra, mint az éhezés, indukálódik. Nagyon ellenálló. - meleg, hideg, kiszáradás, UV sugárzás. spórák.")
73
Spórázás

74
A Bacillus endospóra modell rendszer
DNS-SASP komplex dipikolinsav SASP: small acid soluble proteins

75
Az endospóra képződésének diszkrét fejlődési állomásai

76
Előspóra képződés az anyasejtben

77
Végül a spóra kiszabadul, az anyasejt lizál

78
Spóra csírázás Kedvező környezetben a spóra kicsírázik és visszatér a vegetatív szaporodáshoz. vegetatív sejt spóra

79
A spórázáshoz szükséges gének
Összességében több mint 100 gén spo gének - > 50 különböző gén – spórázáskor nyilvánulnak meg ger gének – a csírázáskor megnyilvánuló specifikus gének más gének – számos gén szükséges még spórázáskor/csírázáskor, ezek természetesen máskor is szükségesek

80
A spo géneket aszerint nevezték el, hogy a spórázás melyik fejlődési szakaszában nyilvánult meg a hibájuk

81
A B. subtilis különböző szempotok figyelembevételével dönt a spórázásról
Végül mind a SpoOA fehérje foszforilezében nyilvánul meg

82
A SpoOA~P különböző géneket aktívál és represszál, amelyek a spórázás iniciációjához kellenek
Az elsődleges célpontok közül kettő σ faktor

83
A SpoOA~P s factor kaszkádot indít el
- Számos szabályozási célpont, a σ-ák is, kompartment-specifikusak sA – vegetatív, sH – korai spórázás, sF előspóra, sE – anyasejt, sG, elkülönült spóra, sK – pusztuló anyasejt

84
A sF és a sE mindkét kompartmentben anyasejt specifikus szigma faktorok
sF aktíváláshoz kellenek: anti-s faktor és anti-anti-s faktor

85
A sE aktíválásához a pre-protein proteolitikus hasítása kell
sE érését az előspórában lévő sF aktíválja

86
A kompartmentek közötti criss-cross kommunikáció spórázáskor
Az anyasejt-specifikus s faktorok pre-proteinként képződnek, aktíválásukhoz utómunkálatok szükségesek. Az előspóra-specifikus s faktorokat anti/anti-anti s faktor rendszerek szabályozzák

87
A kromoszóma DNS partíciója alapvetően különbözik a vegetatív szaporodási mechanizmustól

88
A DNS és a fehérjék elhelyezkedése spórázáskor
SpoIIE A konjugációs transzfer rendszerekhez hasonló

89
B. subtilis spórázás: szabályozó kölcsönhatások
A gének, fehérjék és kis molekulák különböző kölcsönhatása kell a spórázás iniciációjához. SinR~SinI SinI inaktíváljaSinR-t AbrB - AbrB represszálja sin operont sinR A H sinI SinR SinI sin operon Spo0A˜P + Spo0A~P aktíválja sin operont

90
Spórázás iniciáció: szabályozó hálózat B. subtilis-ban
protein gene promoter kinA - + H KinA phospho- relay Spo0A˜P Spo0A A spo0A sinR sinI SinI SinR SinR/SinI sigF hpr (scoR) abrB Hpr AbrB spo0E sigH (spo0H) Spo0E F Signal
 abrB. Hpr. AbrB. spo0E. sigH. (spo0H) Spo0E. F. Signal.")
91
EUKARIÓTA GÉNEXPRESSZIÓ

92
A TRANSZKRIPCIÓ SZABÁLYOZÁSA

93
EUKARIÓTA RNS POLIMERÁZOK
VÁLTOZÓ 10 % kis RNS (tRNS & 5S rRNS) NUCLEOPLASMA POL III + 20-40 % mRNS POL II - 50-70 % rRNS NUCLEOLOUS POL I a AMANITIN ÉRZÉKENYSÉG RELATÍV AKTIVITÁS TERMÉK HELY TÍPUS
 NUCLEOPLASMA. POL III % mRNS. POL II % rRNS. NUCLEOLOUS. POL I. a AMANITIN ÉRZÉKENYSÉG. RELATÍV AKTIVITÁS. TERMÉK. HELY. TÍPUS.")
94
RNS POLIMERÁZOK ÖSSZEHASONLÍTÁSA
E. coli a2bb’ EUKARIÓTA I II III L’ L L’ L L’ L b b’ ~ b és b’ CTD a a’ ~ a és a’ KÖZÖS

95
POL II transzkripció iniciáció
POL II-őn kívül még + 7 GTF (GENERAL TRANSCRIPTION FACTORS) A GTF-ek: TFIIA, IIB, IID, IIE, IIF, IIH & IIJ
 A GTF-ek: TFIIA, IIB, IID, IIE, IIF, IIH & IIJ.")
96
Transzkripció INICIÁCIÓ - RNS polimeráz kötödése
- DNS lokális kitekerése ELONGÁCIÓ - Komplementer bázispárosítás - foszfodiészter kötések kialakítása - 5’ → 3’ transzlokáció TERMINÁCIÓ - nem specifikus és rosszul meghatározott

97
POL II RNS szintézis DNS templátról
Az iniciáció és az elongáció során egy sor általános és specifikus transzkripciós faktorral lép kölcsönhatásba 8-12 alegységből épül fel Méreteik változóak 240→16 kDa

98
POL II alegységek

99
POL II nagy 240KD-os alegysége
E coli b’-höz hasonló Nagyon konzerválódott – ember/egér 95% C terminális ismétlődő CTD TYR SER PRO THR SER PRO SER 52X egérben 26X élesztőben CTD esszenciális az életképességhez

100
140 KD-os alegység E coli b - hoz hasonlít
A H1hisztonnal keresztreagál

101
SOKLÉPCSŐS MODELL PRE-INICIÁCIÓS KOMPLEX
H J E POL II IIF CTD IIA IIB IID INR TATA UPE -20 +1

102
SOKLÉPCSŐS MODELL INICIÁCIÓS KOMPLEX CTD foszforiláció
ATP IID IIA IIB H J E POL II IIF CTD INR TATA UPE -20 +1 P P P P

103
SOKLÉPCSŐS MODELL PROMÓTER CLEARANCE
RIBONUCLEOTIDES pre-mRNA H J E POL II IIA IIB IID INR TATA UPE -20 +1 P P P P ELONGATION FACTORS CTD

104
INICIÁCIÓ

105
PROMÓTER CLEARANCE

106
ELONGÁCIÓ

107
HOLOENZIM

108
HOLOENZYME

109
HOLOENZIM KOMPLEX ELŐSZERELT
POL II IIF CTD H J IIB E IIA IID UPE TATA INR -20 +1

110
HOLOENZIM KOMPLEX ELŐSZERELT
POL II IIF CTD H J IIB IIA IID UPE TATA INR -20 +1

111
VÁLTOZÁSOK AZ EXPRESSZÁLÓDÓ CHROMATINBAN
DNAase I ÉRZÉKENSÉG DNAase I HIPERÉRZÉKENYSÉG (kibomlik) DEMETILÁLÓDÁS (RE érzékenység: MspI (Met), HpaII) aktív gének gyengén metiláltak HISZTONOK ACETILEZÉSE (C-terminális vég lizinben gazdag, acetilezés semlegesíti, csökkenő kötődés)
 DEMETILÁLÓDÁS (RE érzékenység: MspI (Met), HpaII) aktív gének gyengén metiláltak. HISZTONOK ACETILEZÉSE (C-terminális vég lizinben gazdag, acetilezés semlegesíti, csökkenő kötődés)")
112
CHROMATIN ALAPEGYSÉG = NUKLEOSZÓMA 200 bp DNS H2A H2A H1 H2B H2B H1 H3

113
BIZONYÍTÉK A NUKLOSZÓMÁRA
CHROMATIN-t Micricoccus nukleázzal emésztve 200 bp-os „létrát” kapunk 2OO 6OO 4OO 8OO 10OO

114
EUKARIÓTA PROMÓTER TATA Inr -100 -30 Py2CAPy5
UPSTREAM PROMOTER ELEMENTS (UPEs) TATA Inr -100 -30 Py2CAPy5 Iniciáció általában egy A -nál
 TATA. Inr Py2CAPy5. Iniciáció általában egy A -nál.")
115
POL I, POL II ÉS POL III PROMÓTEREK
AdML UPE -60 TATA -30 Inr +1 POL III tRNA (internal) A +20 B +60 tRNA PSE -60 TATA -30 POL I rRNA UCE -90 CORE
 A +20. B +60. tRNA. PSE -60. TATA -30. POL I. rRNA. UCE -90. CORE.")
116
POL I, POL II ÉS POL III PROMÓTEREK
Minden promóterhez kell a TFIID TATA kötő helye

117
Hogyan vizsgáljuk a promótereket?
Klónozzuk a gén 5’ végét és keressük a konzerválódott motívumokat Megkeressük a fehérjéhez kötödő DNS szekvenciákat: band shifts, footprinting Funkcionális vizsgálatok: helyspecifikus mutagenezis, riporter gén

118
Upstream elemek, enhancerek, silencerek
Tipikus pol II: Eukarióta promóterek:

119
Transzkripciós aktivátorok
HTH motívum HLH motívum Zinc-finger

120
Transzkripciós aktivátorok
Leucin-zipper:

121
Példa: zinc finger, élesztő GAL4, GAL80
Galaktóz indukál

122
Transzkripció után: RNS splicing

123
RNS érés mechanizmusa Splicing szignálok:
5' - AG|GUAAGU – intron –YNCURAC – YnNAG|G - 3'

124
Splicosome

125
Alternatív splicing

126
Self-splicing intronok: I., II. csoport
I. csoport rRNS prekurzorok:

127
Self-splicing intronok: I., II. csoport
II. csoprt mitokondrium és kloroplaszt génekben:

128
5’sapka és 3’poliA farok

129
5’ sapka: RNS stabilitás

130
3’ polA farok: RNS stabilitás
Poliadenilációs szignál : AAUAAA

131
Rövidítés Teljes név Funkció CPSF cleavage and polyadenylation specificity factor hasításhoz, adenilációhoz is kell, köt a AAUAAA-hoz CF-I cleavage factor I RNS kötő fehérje, csak a hasításhoz kell CF-II cleavage factor II csak a hasításhoz kell CstF cleavage stimulation factor csak hasításhoz kell, a GU/U régióhoz köt PAP poly [A] polymerase poli [A] szintézist katalizálja és a hasításhoz is PAB II pol [A] binding protein II poliadenilációt stimulálja, segíti poli [A] farok növekedését

132
CPSF kék, CF I és II barna, CstF szürke
PAP piros PAB II sárga

133
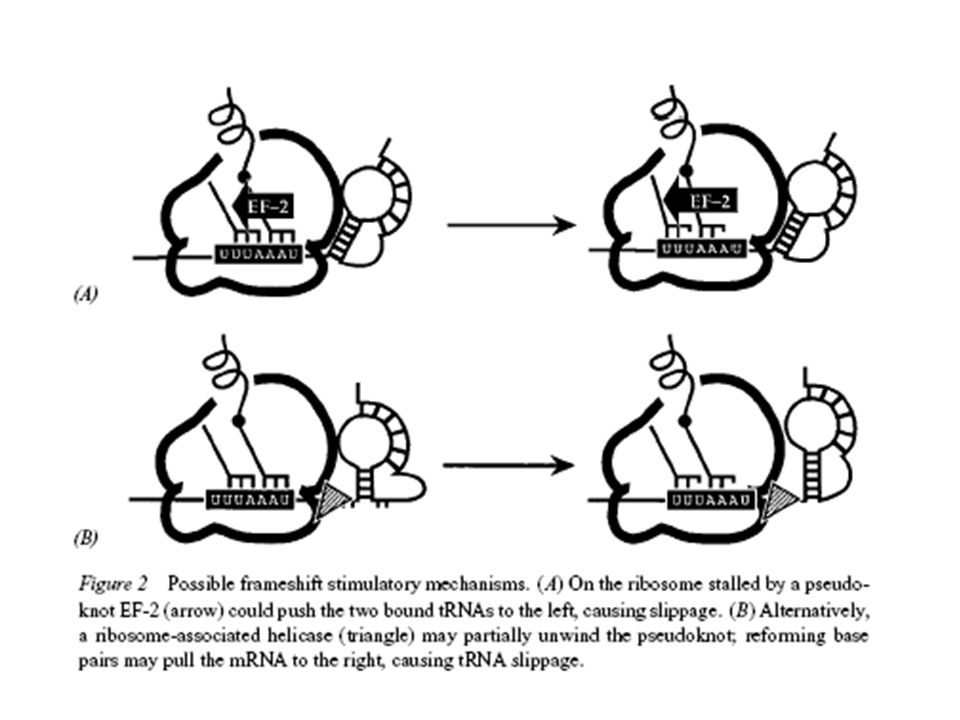
MÁS: Transzlációs frameshifting

135
Transzlációs bypass: T4 gene 60 (topoizomeráz)
")
136
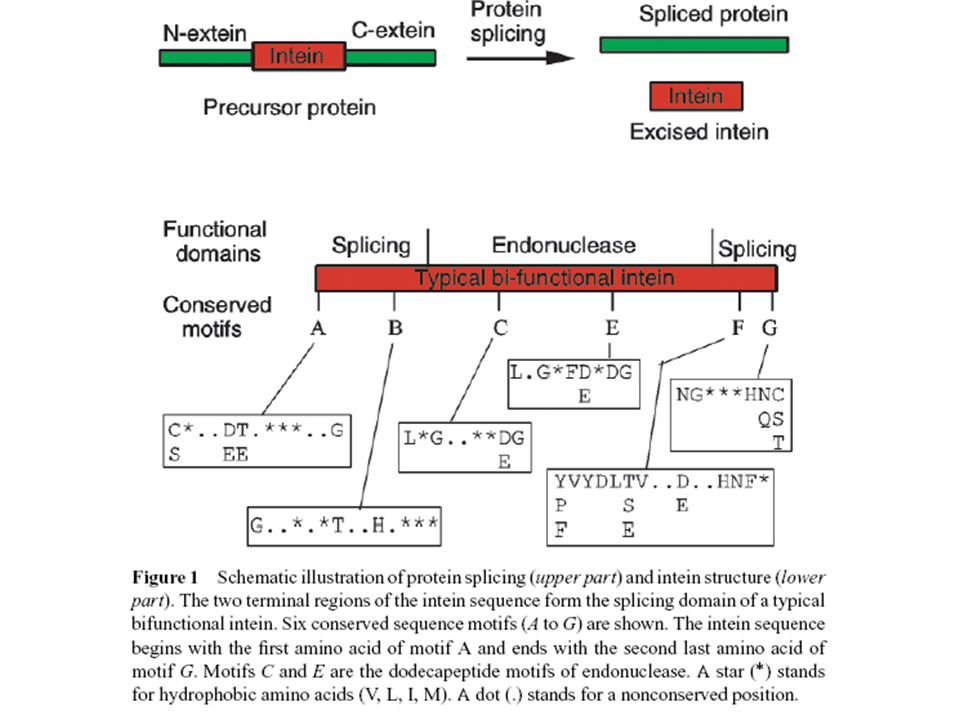
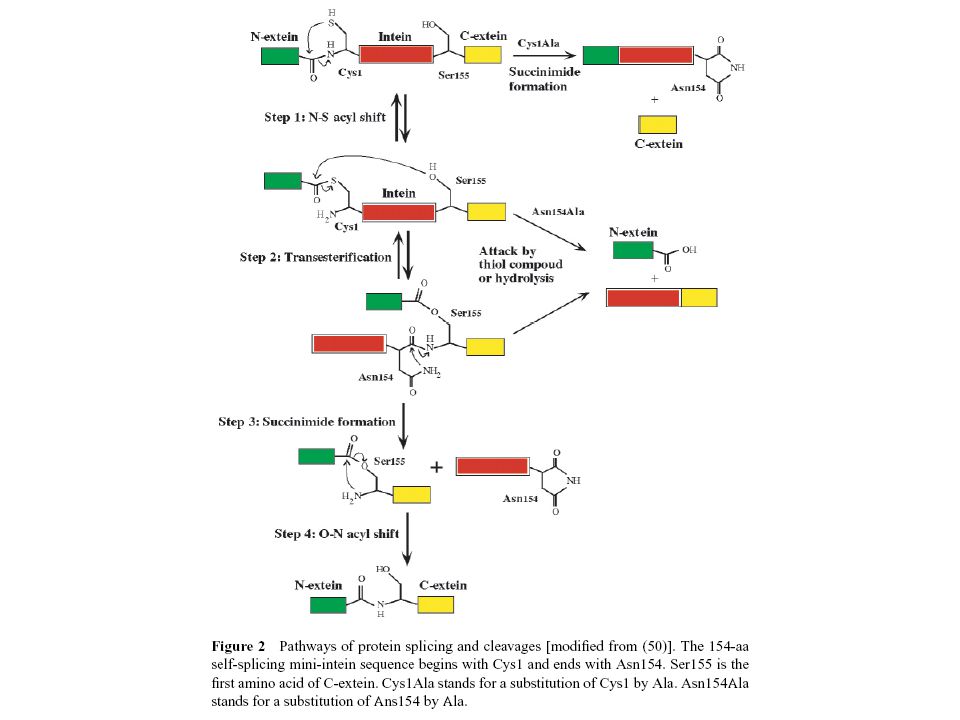
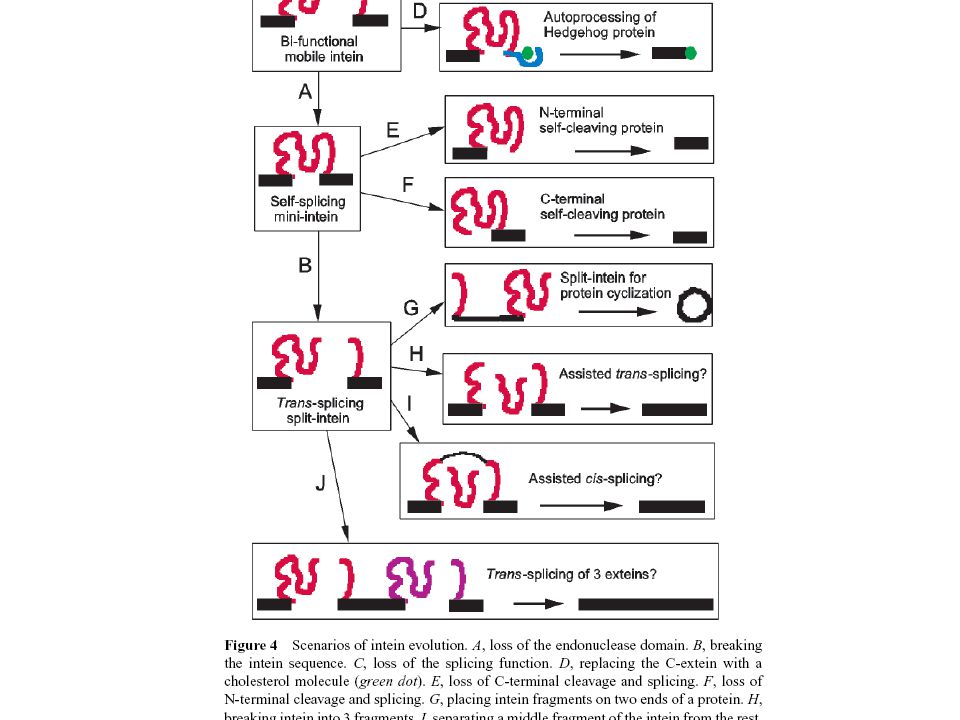
INTEIN, EXTEIN

Hasonló előadás

 DNS-ből,>")



 megváltozHAT Ebből visszakövetkeztethetünk a mutációt szenvedett gén funkciójára.>")




 prokariótában>")





>")

 Bihari Péter.>")
 Csernetics Árpád Bioinformatika SZIT 2005. ápr. 18.>")
