Előadást letölteni
Az előadás letöltése folymat van. Kérjük, várjon

1
A génaktivitás szabályozása

2
Nem minden gén nyilvánul meg állandóan
Sejt típusa (egérben ~200), sejtciklus állapota Minden sejttípusban ugyanaz a genetikai állomány (kevés kivétellel) Egy bizonyos géntermék szintézisét génszabályozási mechanizmusok ellenőrzik A génaktivititást a transzkripció szinten Szignálok kívülről, vagy sejten belülről Ha kell ki/bekapcsolás
, sejtciklus állapota. Minden sejttípusban ugyanaz a genetikai állomány (kevés kivétellel) Egy bizonyos géntermék szintézisét génszabályozási mechanizmusok ellenőrzik. A génaktivititást a transzkripció szinten. Szignálok kívülről, vagy sejten belülről. Ha kell ki/bekapcsolás.")
3
A génexpresszió ellenőrzési pontjai
DNS átrendeződés – DNS szekvencia helye a genomban Transzkripciós szabályozás – RNS szintézis iniciáció/termináció RNS processzálás – splicing/alternatív splicing Transzlációs szabályozás mRNS stabilitás Poszttranszlációs szabályozás – enzimaktivitás, aktiválás, stabilitás, stb.

4
Prokarióták vs eukarióták
Prokarióták – maximális növekedés az adott körülmények között, kivéve, ha végzetes Transzkripció és transzláció egyszerre, egy helyen (splicing nem) Eukarióták – egyedfejlődés, osztódás és specializálódás Felnőttben – fenntartás, tápanyagok nem változnak olyan drasztikusan
 Eukarióták – egyedfejlődés, osztódás és specializálódás. Felnőttben – fenntartás, tápanyagok nem változnak olyan drasztikusan.")
5
Transzkripció szabályozása prokariótákban
Gyakran ki/be, csak akkor,ha a géntermék kell (A Ki leggyakrabban nagyon kicsi expressziót jelent (eukariótákban az egyedfejlődésnél)) Koordinált szabályozás – egy, vagy több metabolizmus útvonal génjei együtt, policisztronos mRNS (eukarióta monocisztronos)
) Koordinált szabályozás – egy, vagy több metabolizmus útvonal génjei együtt, policisztronos mRNS (eukarióta monocisztronos)")
6
Katabolizmus útvonalak (degradatív)
Indukálható, kis inducer molekula Anabolikus útvonalak (bioszintézis) Végtermék elég, represszálható enzimszintézis, kis korepresszor molekula
 Végtermék elég, represszálható enzimszintézis, kis korepresszor molekula.")
7
Két fő kategória: negatív, pozitív szabályozás

8
Indukálható rendszer:
Negatív szabályozás: a represszor fehérje megakadályozza a transzkripciót; az inducer antagonizálja a represszort; van transzkripció Represszálható rendszer: aporepresszor+korepresszor=funkcionális represszor nincs transzkripció Pozitív szabályozás: mRNS csak akkor, ha transzkripciós aktivátor kapcsolódik az aktiválandó gén megfelelő régiójához Autoreguláció: fehérje a saját génjét +/-

9
Laktóz metabolizmus és az operon
E. coli laktóz hasznosítás szabályozásának genetikai analízise Lac- mutánsok Két gén - β-galaktozidáz, laktóz permeáz – szükséges Néhány száz Lac- mutáns: kromoszómán és F’lac-on F’×F- mating = parciális diploidok; F’ lac- / lac+ és F’ lac+ / lac- = mind Lac+ F’ lac- × lac- → Lac+ fenotípusra szelektálva 2 komplementációs csoport, legalább két gén (lacZ, lacY), legalább két gén! (LacA laktóz transzacetiláz később, mert nem esszenciális a laktóz hasznosításhoz.)
, legalább két gén! (LacA laktóz transzacetiláz később, mert nem esszenciális a laktóz hasznosításhoz.)")
10
F’lacY- lacZ+/lacY+lacZ- és
F’lacY+lacZ-/lacY-lacZ+ → Lac+ fenotípus F’lacY-laZ+/lacY-lacZ+ és F’lacY+lacZ-/lacY+lacZ- → Lac- fenotípus lacZ → β-galaktozidáz lacY → laktóz permeáz lacZ és lacY nagyon közel, kotranszdukcióval térképezve

11
Génexpresszió baktériumokban – néhány fontos megkülönböztetés:
Szabályozott gének Sejt növekedés és sejtosztódás szabályozása. Megnyilvánulásukat (expresszió) a sejt szükségletei és a környezet szabályozzák, szükség szerint, nem folyamatosan működnek. Konstitutív gének Folyamatosan kifejeződnek. Háztartási gének (housekeeping genes), mint pl. a fehérjeszintézishez és a glükóz metabolizmushoz szükséges gének. *Valamelyik szinten minden gén szabályozott!
 a sejt szükségletei és a környezet szabályozzák, szükség szerint, nem folyamatosan működnek. Konstitutív gének. Folyamatosan kifejeződnek. Háztartási gének (housekeeping genes), mint pl. a fehérjeszintézishez és a glükóz metabolizmushoz szükséges gének. *Valamelyik szinten minden gén szabályozott!")
12
Operon – mi az? Gének csoportja (cluster), amelyeknek expresszióját operátor-represszor fehérje kölcsönhatás szabályozza, operátor régió és a promóter. Egy operon részei: Promóter Represszor Operator (ellenőrző hely) Kódoló szekvenciák (gének, cisztronok, ORF-ek) Terminátor Egymás melleti kódoló szekvenciák (pl., baktériumok, mtDNS) együtt íródnak át poligénes (policisztronos) mRNS-sé.
, amelyeknek expresszióját operátor-represszor fehérje kölcsönhatás szabályozza, operátor régió és a promóter. Egy operon részei: Promóter. Represszor. Operator (ellenőrző hely) Kódoló szekvenciák (gének, cisztronok, ORF-ek) Terminátor. Egymás melleti kódoló szekvenciák (pl., baktériumok, mtDNS) együtt íródnak át poligénes (policisztronos) mRNS-sé.")
13
Inducer és indukció: Inducer = kémiai, vagy környezeti ágens, ami egy operon transzkripcióját iniciálja. Indukció = géntermék(ek) szintézise az inducerre adott válaszként.
 szintézise az inducerre adott válaszként.")
14
Operon szerveződése

15
E. coli lac operonja: Az E. coli a glükóz metabolizmus géneket folyamatosan kifejezi (konstitutív gének). Az alternatív cukorok (pl. laktóz) metabolizmusa specifikusan szabályozott. laktóz = diszaccharid (glükóz + galaktóz), energia forrás. A laktóz inducer (efektor molekula), három fehérje termelését az 1000-szeresére növeli: -galaktozidáz (lacZ) A laktózt glükózra + galaktózra hidrolizálja. A laktózt allolaktózzá alakítja, ez regulálja a lac operont. Laktóz permeáz (lacY) Laktóz transzport a citoplazma membránon. Transzacetiláz (lacA) A többi szacharidot acetilezi.
 metabolizmusa specifikusan szabályozott. laktóz = diszaccharid (glükóz + galaktóz), energia forrás. A laktóz inducer (efektor molekula), három fehérje termelését az 1000-szeresére növeli: -galaktozidáz (lacZ) A laktózt glükózra + galaktózra hidrolizálja. A laktózt allolaktózzá alakítja, ez regulálja a lac operont. Laktóz permeáz (lacY) Laktóz transzport a citoplazma membránon. Transzacetiláz (lacA) A többi szacharidot acetilezi.")
16
Az E. coli lac operon: Francois Jacob és Jacques Monod Két különböző mutáció típust vizsgáltak a lac operonban: Mutációk a fehérje-kódoló génszekvenciákban. Mutációk a szabályozó szekvenciákban.

17
Fehérje kódoló gének szekvenciáiban mutánsokkal térképezték a géneket:
lacZ (-galaktozidáz) knock-out mutánsok (funkció vesztés) gátolják a laktóz permeáz (lacY) és a transzacetiláz (lacA) funkciókat. lacY (laktóz permeáz) knock-out mutánsok gátolják a transzacetiláz (lacA) funkciót, de nincsenek hatással a -galaktozidázra (lacZ). lacA (transzacetiláz) knock-out mutánsok sem a -galaktozidázt (lacZ) sem pedig a laktóz permeázt (lacY) nem befolyásolják. Következtetés: a 3 lac operon gén a következő sorrendben kapcsolt: lacZ lacY lacA
 knock-out mutánsok (funkció vesztés) gátolják a laktóz permeáz (lacY) és a transzacetiláz (lacA) funkciókat. lacY (laktóz permeáz) knock-out mutánsok gátolják a transzacetiláz (lacA) funkciót, de nincsenek hatással a -galaktozidázra (lacZ). lacA (transzacetiláz) knock-out mutánsok sem a -galaktozidázt (lacZ) sem pedig a laktóz permeázt (lacY) nem befolyásolják. Következtetés: a 3 lac operon gén a következő sorrendben kapcsolt: lacZ. lacY. lacA.")
18
A lac operon transzlációja vad típusú és mutáns E. coli-ban.

19
2. A gén-megnyilvánulást befolyásoló mutációk a regulátor szekvenciákban:
Jacob és Monod olyan mutánsokat is vizsgált, amelyek annak függvényében termelték a lac operon fehérjéit, hogy jelen volt-e az inducer laktóz: 2 típusú upstream lac regulátor mutánst jósoltak meg: Mutációk a lac operátorban (lacO) Mutációk a lac represszorban (lacI)
 Mutációk a lac represszorban (lacI)")
20
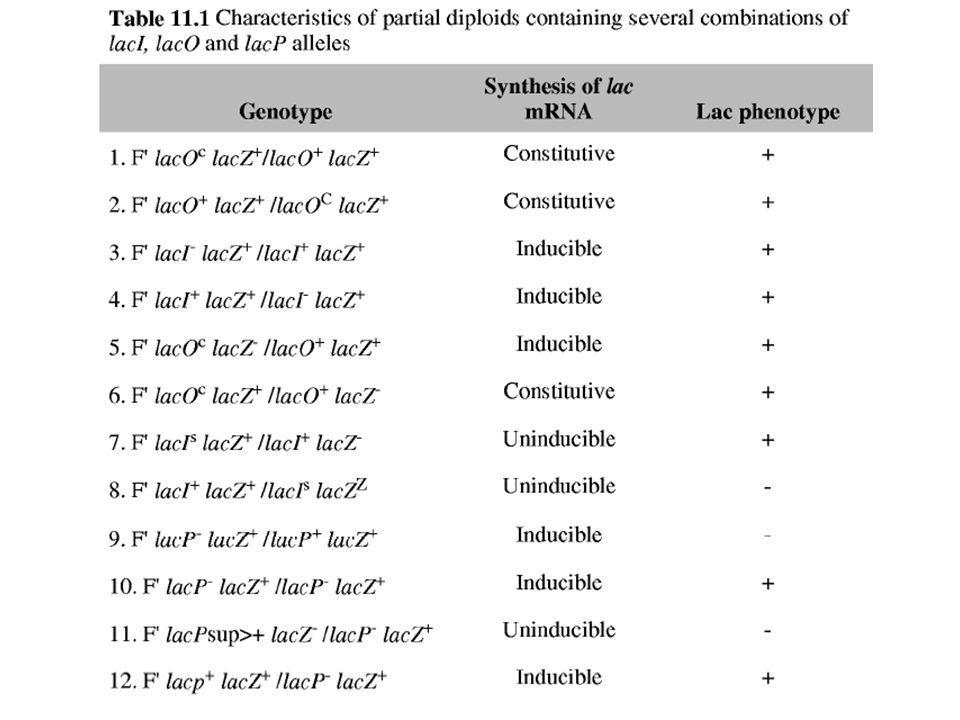
Plazmid F’ lacO+ lacZ- lacY+ permeáz (csak laktózzal)
Mutációk a lac operátorban (lacO): Parciális diploid E. coli törzseket használtak: lac operon gének normális promóterrel a plazmidon (F’) . Plazmid F’ lacO+ lacZ- lacY+ permeáz (csak laktózzal) Kromoszóma C lacOc lacZ+ lacY- -galaktozidáz (laktóz nélkül is) (folyamatosan megnyilvánul, konstitutív) Következtetések: A lacO a lacZ és a lacY előtt helyezkedik el és az utána következő fehérjék termelését befolyásolja ugyanazon a molekulán. A lacO egy szabályozó DNS szekvencia; nincs diffúzóra képes géntermék. Ha lenne diffúzióra képes géntermék, akkor laktóz nélkül is termelődne permeáz.
: Parciális diploid E. coli törzseket használtak: lac operon gének normális promóterrel a plazmidon (F’) . Plazmid F’ lacO+ lacZ- lacY+ permeáz (csak laktózzal) Kromoszóma C lacOc lacZ+ lacY- -galaktozidáz (laktóz nélkül is) (folyamatosan megnyilvánul, konstitutív) Következtetések: A lacO a lacZ és a lacY előtt helyezkedik el és az utána következő fehérjék termelését befolyásolja ugyanazon a molekulán. A lacO egy szabályozó DNS szekvencia; nincs diffúzóra képes géntermék. Ha lenne diffúzióra képes géntermék, akkor laktóz nélkül is termelődne permeáz.")
21
Mutációk a lac represszorban (lacI):
Szintén parciális diploid E. coli F’ törzsekkel; lac operon gének normális promóterrel és normális operátorral. F’ lac I+ lacO+ lacZ- lacY+ C lacI- lacO+ lacZ+ lacY- Laktóz nélkül, nincs -galaktozidáz és permeáz termelés. Laktózzal (inducer), -galaktozidáz és permeáz szintetizálódnak. Következtetés: lacI+ represszor fehérjét kódol (diffúzióra képes termék). Laktóz nélkül a represszor fehérje kötődik az operátorhoz és gátolja a downstream fehérjék termelődését. Laktóz jelenlétében a represszor fehérjét gátolja az allolaktóz és a fehérjék termelődnek.
, -galaktozidáz és permeáz szintetizálódnak. Következtetés: lacI+ represszor fehérjét kódol (diffúzióra képes termék). Laktóz nélkül a represszor fehérje kötődik az operátorhoz és gátolja a downstream fehérjék termelődését. Laktóz jelenlétében a represszor fehérjét gátolja az allolaktóz és a fehérjék termelődnek.")
22
Mutáció lehet a promóterben is (Plac):
Gátlódhat az RNS polimeráz kötődése és nincs fehérje szintézis a struktúrgénekről, ha van laktóz, ha nincs. Egyetlen mutáció mindhárom fehérje génjét befolyásolja, lacZ, lacY, és lacA.

23
A lac operon szerveződése a vad típusú E. coli-ban.
A szabályozó elemek és a gének sorrendje: lacI: promóter-lacI-terminátor operon: promóter-operátor-lacZ-lacY-lacA-terminátor

24
Az E. coli lac operon funkcionális állapota laktóz nélkül:

25
Az E. coli lac operon funkcionális állapota laktózon növesztve :

26
A lac represszor modellje tetramer (4 polipetid) fehérje.
 fehérje.")
27
A Jacob-Monod féle E. coli lac operon modell:
Az operon egy génklaszter; a gének megnyilvánulásást opero-represszor kölcsönhatás szabályozza. A lac I génnek van saját gyenge promótere és terminátora; lacI represszor fehérje mindig van kis mennyiségben. A represszor fehérje tetramer (4 polipeptid). A represszor kötődik az operátorhoz (lacO) és akadályozza az RNS polimerázt a transzkripció iniciációjában. A kötődés reverzibilis, így mindig van kevés LacZ, LacY, és LacA fehérje. Amint nagy mennyiségű laktóz kerül a tápoldatba a lac operon azonnal bekapcsol. A laktózon növekvő vad típusú E. coli-ban a -galaktozidáz a laktózt átalakítja allolaktózzá. Az allolaktóz kapcsolódik a represszorhoz, ezért a represszor fehérje alakja megváltozik és nem tud kötődni az operátorhoz. Az allolaktóz indukálja a lac operon expresszióját.
. A represszor kötődik az operátorhoz (lacO) és akadályozza az RNS polimerázt a transzkripció iniciációjában. A kötődés reverzibilis, így mindig van kevés LacZ, LacY, és LacA fehérje. Amint nagy mennyiségű laktóz kerül a tápoldatba a lac operon azonnal bekapcsol. A laktózon növekvő vad típusú E. coli-ban a -galaktozidáz a laktózt átalakítja allolaktózzá. Az allolaktóz kapcsolódik a represszorhoz, ezért a represszor fehérje alakja megváltozik és nem tud kötődni az operátorhoz. Az allolaktóz indukálja a lac operon expresszióját.")
28
Az RNS polimeráz egyetlen poligénes mRNS szintézisét iniciálja, a lacZ, lacY, és lacA gének mRNS-ét.
mRNS egyetlen molekulaként transzálódik riboszóma füzérrel. A lac operon úgynevezett negatív kontroll alatt áll (a lacI blokkolja az RNS polimerázt, ha nincs indukáló ágens) Különböző típusú mutációk lehetnek a lacO, alacI, és a promóter régiókban: lacO -megváltozik a represszor kötőhelye (represszor nem tud kötődni) -állandó, konstitutív exszpresszió lacI -a represszor konformáció változik meg (nem tud kötődni az operátorhoz) -konstitutív -szuper-represszor köti az operátort, de az allolaktózt nem -a laktóz nem tudja indukálni az operont, nincs expresszió promóter -megváltozik affinitása az RNS polimerázhoz -növeli, vagy csökkenti a transzkripció mértékét
 Különböző típusú mutációk lehetnek a lacO, alacI, és a promóter régiókban: lacO -megváltozik a represszor kötőhelye (represszor nem tud kötődni) -állandó, konstitutív exszpresszió. lacI -a represszor konformáció változik meg (nem tud kötődni az operátorhoz) -konstitutív. -szuper-represszor köti az operátort, de az allolaktózt nem. -a laktóz nem tudja indukálni az operont, nincs expresszió. promóter -megváltozik affinitása az RNS polimerázhoz. -növeli, vagy csökkenti a transzkripció mértékét.")
30
A lac operon pozitív szabályozás alatt is áll:
Pozitív a lac operon szabályozása, ha az E. coli egyedüli C-forrása a laktóz (de nem, ha glükóz is van a tápoldatban). A katabolit aktívátor fehérjéhez (catabolite activator protein) (CAP) kötődik a cAMP, aktíválódik, és kapcsolódik a promóter előtti CAP felismerő helyhez (cAMP koncentráció nagyon kicsi glükóz jelenlétében). CAP megváltoztatja a DNS konformációját, megkönnyítve az RNS polimeráz kötődését és a transzkripciót. Ha glükóz és laktóz egyszerre vannak jelen, a glükózt hasznosítja először a kevés cAMP miatt nem aktíválódik elegendő CAP. (diauxiás növekedés) Ha feleslegben adunk cAMP-ot, akkor lesz transzkripció a lac operonról, még glükóz jelenlétében is. A LacI lokális, helyi szabályozó. A CAP globális, máshol is ható szabályozó.
. A katabolit aktívátor fehérjéhez (catabolite activator protein) (CAP) kötődik a cAMP, aktíválódik, és kapcsolódik a promóter előtti CAP felismerő helyhez (cAMP koncentráció nagyon kicsi glükóz jelenlétében). CAP megváltoztatja a DNS konformációját, megkönnyítve az RNS polimeráz kötődését és a transzkripciót. Ha glükóz és laktóz egyszerre vannak jelen, a glükózt hasznosítja először a kevés cAMP miatt nem aktíválódik elegendő CAP. (diauxiás növekedés) Ha feleslegben adunk cAMP-ot, akkor lesz transzkripció a lac operonról, még glükóz jelenlétében is. A LacI lokális, helyi szabályozó. A CAP globális, máshol is ható szabályozó.")
31
A lac operon pozitív szabályozása CAP-val

32
A lac operon szekvencia volt az első jól jellemzett génszabályozási modell :
A lac operon promóter -84 bp-ra kezdődik, közvetlenül a lacI stop kodon után és -8 bp-ra végződik a transzkripciós starthelytől. CAP-cAMP kötőhelyek -54 →-58 és -65 → -69. RNS polimeráz kötőhely -47 → -8. Operátor a promóter mellett -3 → +21. mRNS transzkriptum +1 bp-nál kezdődik, az operátorban. A -galaktozidáz génnek a start kodon előtt van egy vezető szekvenciája (riboszóma kötőhely/Shine-Delgarno szekvencia) A -galaktozidáz start kodonodon (AUG) +39 → +41
 A -galaktozidáz start kodonodon (AUG) +39 → +41.")
33
Lac operon szabályozó szekvenciái

34
Az E. coli triptofán operon:
Ha van aminosav a tápoldatban, akkor „importálja” az aminosavakat, mielőtt szintetizálna. Az aminosav szintézis gének represszáltak, represszálható operonok. Ha nincs aminosav a táptalajban, a gének „bekapcsolnak” (megnyilvánulnak) és lesz aminosav szintézis. Az E. coli triptofán operonja az egyik legjobban tanulmányozott represszálható operon.
 és lesz aminosav szintézis. Az E. coli triptofán operonja az egyik legjobban tanulmányozott represszálható operon.")
35
Az E. coli Trp operon; (Charles Yanofsky és mtsai.):
A Trp operon ~7kb méretű és 5, a triptofán szintézishez szükséges génterméket kódol , trpA-E. A promóter és az operátor a trpE előtt van. A vezető régió (trpL) a trpA-E struktúrgének és az operátor között helyezkedik el. A trpL-ben van egy attenuátor régió (att). TrpR (represszor fehérje gén) a promóter előtt van.
 a trpA-E struktúrgének és az operátor között helyezkedik el. A trpL-ben van egy attenuátor régió (att). TrpR (represszor fehérje gén) a promóter előtt van.")
36
Az E. coli Trp operon szerveződése:

37
A trp operon szabályozása:
A trp operont két mechanizmus szabályozza: Represszor/operátor kölcsönhatás A megkezdett transzkripció attenuációja (a transzkripció megakad)
")
38
1. Represszor/operátor kölcsönhatás
Ha van triptofán, akkor kötődik a trpR gén termékéhezt. trpR fehérje kapcsolódik a trp operátorhoz és megakadályozza a transzkripciót. A represszió ~70-edére csökkenti a trp operon transzkripciójának mértékét.

39
2. A megkezdett transzkriptum terminációja
A transzkripciót attenuáció is szabályozza, egy rövid, nem komplett fehérje transzlálódik. Ha a sejtek „éheznek” triptofánra, akkor a trp gének maximálisan kifejeződnek. Kevésbé súlyos éhezés esetén, csak a maximálisnál kisebb mértékben nyilvánulnak meg. Az attenuáció a transzkripció mértékét 8-10-szeres faktorral szabályozza; a repressziós mechanizmussal kombinálódva szoros mértékben csökkentheti.

40
Az attenuáció molekuláris modellje:
A vezető régió (trpL) az operator és a trpE szekvenciák között van. E vezető szekvenciában van az attenuációs régió (att). Az att szekvenciában start kodon, 2 Trp kodon, egy stop kodon, és négy olyan szekvencia régió, amelyek három alternatív másodlagos szerkezet kialakítására képesek. Másoglagos szerkezet Szignál H-híd kötések 1-2 régió szünet H-híd kötések 2-3 régió antitermináció H-híd kötések 3-4 régió termináció
 az operator és a trpE szekvenciák között van. E vezető szekvenciában van az attenuációs régió (att). Az att szekvenciában start kodon, 2 Trp kodon, egy stop kodon, és négy olyan szekvencia régió, amelyek három alternatív másodlagos szerkezet kialakítására képesek. Másoglagos szerkezet Szignál. H-híd kötések 1-2 régió szünet. H-híd kötések 2-3 régió antitermináció. H-híd kötések 3-4 régió termináció.")
41
A vezető /attenuátor trp operon szekvencia:

42
Emlékezzünk arra, hogy prokariótákban a transzkripció és a transzláció egyszerre, egy helyen.
Az mRNS 1 és 2 régiói között kialakuló H-híd kötések (hajtű, stem loop) megakasztja az RNS polimerázt, amint ez a rész szintetizálódott. A szünet éppen elég hosszú ahhoz, hogy a riboszóma kapcsolódjon az mRNS-hez és elkezdje a transzlációt közvetlenül az RNS polimeráz mögött.
 megakasztja az RNS polimerázt, amint ez a rész szintetizálódott. A szünet éppen elég hosszú ahhoz, hogy a riboszóma kapcsolódjon az mRNS-hez és elkezdje a transzlációt közvetlenül az RNS polimeráz mögött.")
43
A riboszóma helyzetének fontos szerepe van az attenuációban:
Ha a Trp kevés, vagy nincs utánpótlás (és szükséges): Trp-tRNS-ek nincsenek, a riboszóma megáll a Trp kodonoknál és lefedi az 1-es attenuátor régiót. Az 1-es és 2-es régiók között nem tud kialakulni a hajtű szerkezet, ehelyett a 2-es régió a 3-assal alakít ki hajtűt, amint az elkészült. A 3-as régió (most a 2-essel párban) a 4-essel nem tud kapcsolódni a szintézis után. Az RNS polimeráz folytathatja a 4-es régió után a teljes trp mRNS szintézisét.
: Trp-tRNS-ek nincsenek, a riboszóma megáll a Trp kodonoknál és lefedi az 1-es attenuátor régiót. Az 1-es és 2-es régiók között nem tud kialakulni a hajtű szerkezet, ehelyett a 2-es régió a 3-assal alakít ki hajtűt, amint az elkészült. A 3-as régió (most a 2-essel párban) a 4-essel nem tud kapcsolódni a szintézis után. Az RNS polimeráz folytathatja a 4-es régió után a teljes trp mRNS szintézisét.")
44
A riboszóma helyzete fontos az attenuációnál:
Ha a Trp elegendő (és nem szükséges): A riboszóma nem áll meg a Trp kodonoknál és folytatja a vezető polipeptid szintézisét, befejezve a 2-es régiónál. A 2-es és a 3-as régiók nem képezhetnek hajtűt, ezért az a 3-as és a 4-es régió között alakul ki. A 3-as és a 4-es régió közötti hajtű az „attenuátor” szekvencia és terminációs szignálként szolgál. A transzkripció azelőtt fejeződik be, hogy a triptofán struktúrgéneket elérte volna az RNS polimeráz.
: A riboszóma nem áll meg a Trp kodonoknál és folytatja a vezető polipeptid szintézisét, befejezve a 2-es régiónál. A 2-es és a 3-as régiók nem képezhetnek hajtűt, ezért az a 3-as és a 4-es régió között alakul ki. A 3-as és a 4-es régió közötti hajtű az „attenuátor szekvencia és terminációs szignálként szolgál. A transzkripció azelőtt fejeződik be, hogy a triptofán struktúrgéneket elérte volna az RNS polimeráz.")
45
Az attenuációs modell a Trp éheztetett sejtekben.

46
Attenuáció modellje elegendő Trp jelenlétében.

47
Phe, His, Leu, Thr, és Ile operonok feltételezett attenuátorainak aminosav szekvenciái E. coli-ban.

48
E. coli RNS ploimeráz σ kaszkád
Gén Mw KDa Alk. -35 Szekv. Szepar. -10 Szekv. rpoA 40 a subunit - rpoB 155 b subnit rpoC 160 b' subunit rpoD 70 s70 Fő TTGACA 16-18 bp TATAAT rpoN 54 s54 Nitrogén CTGGNA 6 bp TTGCA rpoS 38 s38 Stacioner nem ismert rpoH 32 s32 hősokk CCCTTGAA 13-15 bp CCCGATNT fliA 28 s28 Flagellum CTAAA 15 bp GCCGATAA rpoE 24 s24 Magas hőm.

49
A Bacillus subtilis életciklusa
B. subtilis spórázik, ha a környezeti feltételek kedvezőtlenné válnak ? Osztódási ciklus Spórázás-csírázás ciklus Metabolit és környezeti jelek Start with a schematic overview of the life cycle of B. subtilis. Use this slide to draw attention to the question mark. This is the important developmental decision.

50
Szabályozási kölcsönhatások
Különböző típusú kölcsönhatások : gének, fehérjék és kis molekulák között a B. subtilis spórázásának szabályozásánál SinR~SinI SinI inaktiválja SinR-t AbrB - AbrB represszálja sin operont sinR A H sinI SinR SinI sin operon Spo0A˜P + Spo0A~P aktiválja sin operont

51
Genetikai szabályozó hálózat B. Subtilis-ban
A B. subtilis spórázás iniciációjának többé kevésbé teljes genetikai szabályozó hálózata Nagy és komplex protein gene promoter kinA - + H KinA phospho- relay Spo0A˜P Spo0A A spo0A sinR sinI SinI SinR SinR/SinI sigF hpr (scoR) abrB Hpr AbrB spo0E sigH (spo0H) Spo0E F Signal The assembled data from many laboratories yield a qualitative scheme of the molecular interactions. A prediction of the global behavior of this system is no longer possible. We therefore have to develop conceptual and computer tools to estimate the behavior of such regulation networks.
 abrB. Hpr. AbrB. spo0E. sigH. (spo0H) Spo0E. F. Signal. The assembled data from many laboratories yield a qualitative scheme of the molecular interactions. A prediction of the global behavior of this system is no longer possible. We therefore have to develop conceptual and computer tools to estimate the behavior of such regulation networks.")
52
A spórázás szimulációja B. subtilis-ban
kinA - + H KinA phospho- relay Spo0A˜P Spo0A A spo0A sinR sinI SinI SinR SinR/SinI sigF hpr (scoR) abrB Hpr AbrB spo0E sigH (spo0H) Spo0E F Signal
 abrB. Hpr. AbrB. spo0E. sigH. (spo0H) Spo0E. F. Signal.")
53
A szimuláció eredményeinek elemzése
A spórázás iniciációja a versengő pozitív és negatív szabályozó feedback hurkok eredményeként felhalmozódó Spo0A~P A spo mutánsok nem képesek visszacsatolásokra KinA kinA H + + + phospho- relay Spo0A˜P + Using only the known interactions leads to an inconsistency. The concentration of Spo0E has to be kept low in order to maintain a stable sporulation state. We therefore need an additional interaction that negatively regulates Spo0E after sporulation has been initiated. Spo0A - + F H sigF Spo0E spo0E A

54
REGULON több operon közös szabályozás alatt Pl. σB regulon

Hasonló előadás
 DNS-ből,>")



 megváltozHAT Ebből visszakövetkeztethetünk a mutációt szenvedett gén funkciójára.>")





 prokariótában>")



>")


 Bihari Péter.>")

 Csernetics Árpád Bioinformatika SZIT 2005. ápr. 18.>")