Előadást letölteni
Az előadás letöltése folymat van. Kérjük, várjon

1
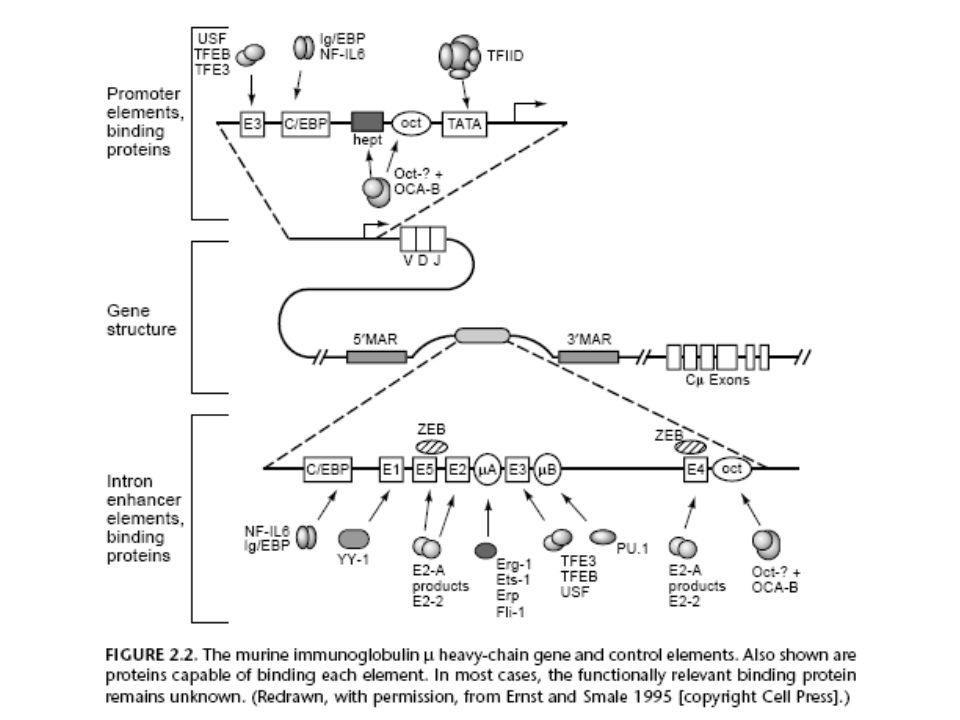
Genetikai szabályozó hálózat B. Subtilis-ban
A B. subtilis spórázás iniciációjának többé kevésbé teljes genetikai szabályozó hálózata Összetett: szignál transzdukció (phosporelay) szenzor/válasz regulátor; szigma kaszkád, szigma, antiszigma, antiantiszigma; represszió, aktiválás, autoaktiváció, stb. protein gene promoter kinA - + H KinA phospho- relay Spo0A˜P Spo0A A spo0A sinR sinI SinI SinR SinR/SinI sigF hpr (scoR) abrB Hpr AbrB spo0E sigH (spo0H) Spo0E F Signal The assembled data from many laboratories yield a qualitative scheme of the molecular interactions. A prediction of the global behavior of this system is no longer possible. We therefore have to develop conceptual and computer tools to estimate the behavior of such regulation networks.
 szenzor/válasz regulátor; szigma kaszkád, szigma, antiszigma, antiantiszigma; represszió, aktiválás, autoaktiváció, stb. protein. gene. promoter. kinA. - + H. KinA. phospho- relay. Spo0A˜P. Spo0A. A. spo0A. sinR. sinI. SinI. SinR. SinR/SinI. sigF. hpr (scoR) abrB. Hpr. AbrB. spo0E. sigH. (spo0H) Spo0E. F. Signal. The assembled data from many laboratories yield a qualitative scheme of the molecular interactions. A prediction of the global behavior of this system is no longer possible. We therefore have to develop conceptual and computer tools to estimate the behavior of such regulation networks.")
2
EUKARIÓTA RNS POLIMERÁZOK
VÁLTOZÓ 10 % kis RNS (tRNS & 5S rRNS) NUCLEOPLASMA POL III + 20-40 % mRNS POL II - 50-70 % rRNS NUCLEOLOUS POL I a AMANITIN ÉRZÉKENYSÉG RELATÍV AKTIVITÁS TERMÉK HELY TÍPUS
 NUCLEOPLASMA. POL III % mRNS. POL II % rRNS. NUCLEOLOUS. POL I. a AMANITIN ÉRZÉKENYSÉG. RELATÍV AKTIVITÁS. TERMÉK. HELY. TÍPUS.")
3
Prokarióta RNS polimeráz

4
RNS POLIMERÁZOK ÖSSZEHASONLÍTÁSA
E. coli a2bb’ EUKARIÓTA I II III L’ L L’ L L’ L b b’ ~ b és b’ CTD a a’ ~ a és a’ KÖZÖS

5
A génexpresszió szabályozása eukariótákban:
Rövid távú, hosszú távú – génszabályozás: fejlődés/differenciálódás A Drosophila egyedfejlődés szabályozása

6
Az eukarióta gének is fehérje-kódoló egységekként és a szomszédos szabályozó régiókkal szabályozódnak, de nem operonok. Az eukarióta gének szabályozása komplexebb, mert van sejtmagjuk. (a transzkripció és a transzláció elkülönülten). Az eukarióta gének szabályozásának két kategóriája: Rövid távú – gének gyors ki/bekapcsolása a környezetre és a sejt igényeire adott válaszként. Hosszú távú – az egyedfejlődés és a differenciálódás génjei.
. Az eukarióta gének szabályozásának két kategóriája: Rövid távú – gének gyors ki/bekapcsolása a környezetre és a sejt igényeire adott válaszként. Hosszú távú – az egyedfejlődés és a differenciálódás génjei.")
7
Az eukarióta gének hat szinten szabályozódnak:
transzkripció RNS processzálás mRNS transzport mRNS transzláció mRNS degradáció protein degradáció

8
A transzkripció szabályozása:
Promóterekkel A transzkripciós starthely előtt. Néhány azt határozza meg, hol kezdődjön a transzkripció (pl. TATA), míg mások, hogy megkezdődjön-e. A promóterek aktivációját specifikus transzkripciós faktor (TF) proteinek (specifikus TF-ek kapcsolódnak specifikus promóterekhez). Egy adott génnek egy, vagy több promótere (mindegyik specifikus TF fehérjékkel) lehet. A promóterek lehetnek pozitívan és negatívan szabályozottak.
, míg mások, hogy megkezdődjön-e. A promóterek aktivációját specifikus transzkripciós faktor (TF) proteinek (specifikus TF-ek kapcsolódnak specifikus promóterekhez). Egy adott génnek egy, vagy több promótere (mindegyik specifikus TF fehérjékkel) lehet. A promóterek lehetnek pozitívan és negatívan szabályozottak.")
9
Enhancerekkel A transzkripciós starthely előtt, vagy után vannak. A szabályozó fehérjék specifikus enhancer szekvenciákhoz kötődnek; a kötödést a DNS szekvencia határozza meg. A TF-ekhez kötődő DNS hurkot képez és így teremti meg a kapcsolatot az enhancer elemekkel. A szabályozó fehérjék közötti kölcsönhatások határozzák meg, hogy a transzkripció aktiválódik,vagy represszálódik, azaz pozitívan, vagy negatívan szabályozott lesz-e. Például

10
Még a promóterekről és az enhancerekről:
Néhány szabályozó fehérje minden sejttípusban van, mások specifikusak. Minden promóternek és enhancernek az expressziót meghatározó specifikus fehérje készlete van. A génmegnyilvánulás mértékét a pozitívan és a negatívan szabályozó fehérjék közötti kölcsönhatások alakítják. Kombinatorikus génszabályozás; enhancerek és promóterek sok hasonló szabályozó fehérjét tudnak kötni, számtalan kölcsönhatási lehetőséggel.

11
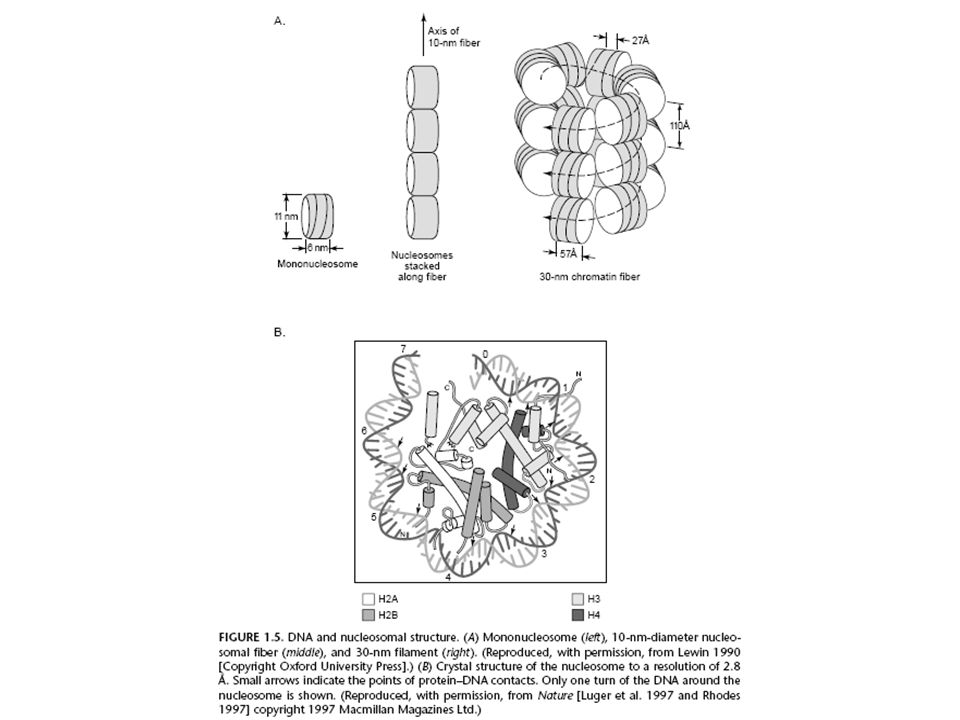
Kromoszómaszerkezet, az eukarióta kromoszóma hisztonokkal van csomagolva:
A prokariótáknak nincsenek hisztonjaik és más struktúr fehérjéik, így a így a DNS közvetlenül elérhető. Az eukariótáknak vannak hisztonjaik és a hisztonok represszálják a transzkripiót, mert interferálnak a DNS-kötő fehérjékkel. Igazolás DNáz I érzékenységi kísérletekkel: A DNáz I lebontja a transzkripciósan aktív DNS-t. A nem-átíródó DNS-t a hiszton pajzs megvédi a DNáztól és nem tudja olyan könnyedén lebontani. Ha kísérletesen adunk hisztonokat és és promóter-kötő fehérjéket, a hisztonok kompetitíven kapcsolódnak a promóterhez és gátolják a transzkripciót. Megoldás: a transzkripciósan aktív géneknek lazább a kromoszóma szerkezete, mint az inaktív géneké. A hisztonok acetilezése és foszforilezése megváltoztatja DNS-kötő képességüket. Az enhancer-kötő fehérjék kompetitíven blokkolják a hisztonokat, ha kísérletesen együtt adjuk hisztonokkal és és promóter-kötő TF-ekkel. Az RNS polimeráz és a TF-ek körülfogják a hiszton/nukleoszómát és lesz transzkripció.

12
DNS metiláció és a transzkripció:
Az újonnan szintetizálódott DNS kis mennyisége (~3% emlősökben) metilezéssel kémiailag módosított. A metiláció leggyakrabban szimmetrikus CG szekvenciáknál. A transzkripciósan aktív gének szignifikánsan kisebb mértékben metilezettek, mint az inaktív gének. A metiláció génje esszenciális az egér egyedfejlődésénél (egy gén kikapcsolása szintén lehet fontos). A törékeny X szindróma emberi betegséget is metilezés eredményezi; az FMR-1 gént metilezés csendesíti.
 metilezéssel kémiailag módosított. A metiláció leggyakrabban szimmetrikus CG szekvenciáknál. A transzkripciósan aktív gének szignifikánsan kisebb mértékben metilezettek, mint az inaktív gének. A metiláció génje esszenciális az egér egyedfejlődésénél (egy gén kikapcsolása szintén lehet fontos). A törékeny X szindróma emberi betegséget is metilezés eredményezi; az FMR-1 gént metilezés csendesíti.")
13
Rövid távú transzkripció szabályozás – a galaktóz hasznosítás génjei élesztőben:
3 gén (GAL1, GAL7, & GAL 10) kódolja a galaktóz metabolizmus útvonalban működő enzimeket. GAL1 galaktokináz GAL7 galaktóz transzferáz GAL10 galaktóz epimeráz Az útvonal végén D-glükóz 6-foszfát képződik, ami belép a glikolízis útvolalba és folyamatosan átíródó gének metabolizálják. Galaktóz nélkül, a GAL gének csendesek. A GAL gének glükóz hiányában és galaktóz jelenlétében gyorsan indukálódnak. Analógia: az E. coli lac operon repressziója glükózzal.
 kódolja a galaktóz metabolizmus útvonalban működő enzimeket. GAL1 galaktokináz. GAL7 galaktóz transzferáz. GAL10 galaktóz epimeráz. Az útvonal végén D-glükóz 6-foszfát képződik, ami belép a glikolízis útvolalba és folyamatosan átíródó gének metabolizálják. Galaktóz nélkül, a GAL gének csendesek. A GAL gének glükóz hiányában és galaktóz jelenlétében gyorsan indukálódnak. Analógia: az E. coli lac operon repressziója glükózzal.")
14
A galaktóz metabolizmus útvonal élesztőben.

15
A GAL gének egymáshoz közel, de nem egy operonban vannak.
Egy másik nem-kapcsolt gén, a GAL4, egy represszor fehérjét kódol, ami egy promóter elemhez kötődik: upstream aktivátor szekvencia (UASG). Az UASG a GAL1 és a GAL10 között helyezkedik el. Transzkripció az UASG-től mindkét irányba. Ha nincs galaktóz, a GAL4 terméke (GAL4p) és egy másik géntermék (GAL80p) kötődik az UASG szekvenciához; nem lesz transzkripció. Ha galaktózt adunk, a galaktóz metabolit kapcsolódik a GAL80p-vel és a GAL4p aminosavak foszforileződnek. A galaktóz inducerként működik, konformáció változást okoz a GAL4p/GAL80p-ben.
. Az UASG a GAL1 és a GAL10 között helyezkedik el. Transzkripció az UASG-től mindkét irányba. Ha nincs galaktóz, a GAL4 terméke (GAL4p) és egy másik géntermék (GAL80p) kötődik az UASG szekvenciához; nem lesz transzkripció. Ha galaktózt adunk, a galaktóz metabolit kapcsolódik a GAL80p-vel és a GAL4p aminosavak foszforileződnek. A galaktóz inducerként működik, konformáció változást okoz a GAL4p/GAL80p-ben.")
16
A GAL gének aktivációs modellje élesztőben.

17
Hormonális szabályozás – egy másik jó példa a rövid távú transzkripció-szabályozásra:
A magasabb rendű eukarióták sejtjei specializálódottak és általában jól védettek a külső környezet gyors változásaitól. A hormon szignálok az egyik mechanizmus, amivel a környezetre reagálva szabályozzák a transzkripciót. Az egyik sejt által termelt hormonok inducerként hatnak és fiziológiai választ váltanak ki egy másik sejtben. A hormonok csak a hormon-specifikus receptorokkal rendelkező célsejtekre hatnak és a hormonszintet feedback útvonalak állítják be. A hormonok két módon szolgáltatnak jelet: A szteroid hormonok átmennek a membránon és a citoplazmás receptorokhoz kapcsolódnak, együtt kötődnek a DNS-hez és szabályozzák a génexpressziót. A polipeptid hormonok a sejtfelszíni receptorokhoz kapcsolódnak és aktiválják a transzmembrán enzimeket, hogy másodlagos hírvivőket szintetizáljanak (pl. cAMP), amelyek a aktiválják a transzkripciót.
, amelyek a aktiválják a transzkripciót.")
18
Emlős szteroid hormonok.
Növény hormonok

19
Hormonális szabályozás:
A szteroid hormonokkal regulált gének szekvenciáiban van kötő régió: szteroid hormone response elements (HREs). HRE-k gyakran több kópiában is az enhancer szekvencia régióban. Ha nincs szteroid: a receptor beborítva és dajka fehérjék (chaperone) „vigyázzák”; nincs transzkripció Ha van szteroid: A szteroid lecseréli a dajka fehérjéket, kötődik a receptorhoz, majd a HRE szekvenciához; megkezdődik a transzkripció.
. HRE-k gyakran több kópiában is az enhancer szekvencia régióban. Ha nincs szteroid: a receptor beborítva és dajka fehérjék (chaperone) „vigyázzák ; nincs transzkripció. Ha van szteroid: A szteroid lecseréli a dajka fehérjéket, kötődik a receptorhoz, majd a HRE szekvenciához; megkezdődik a transzkripció.")
20
A glükokortikoid szteroid hormonszabályozás modellje.

21
Szabályozás RNS processzálódással:
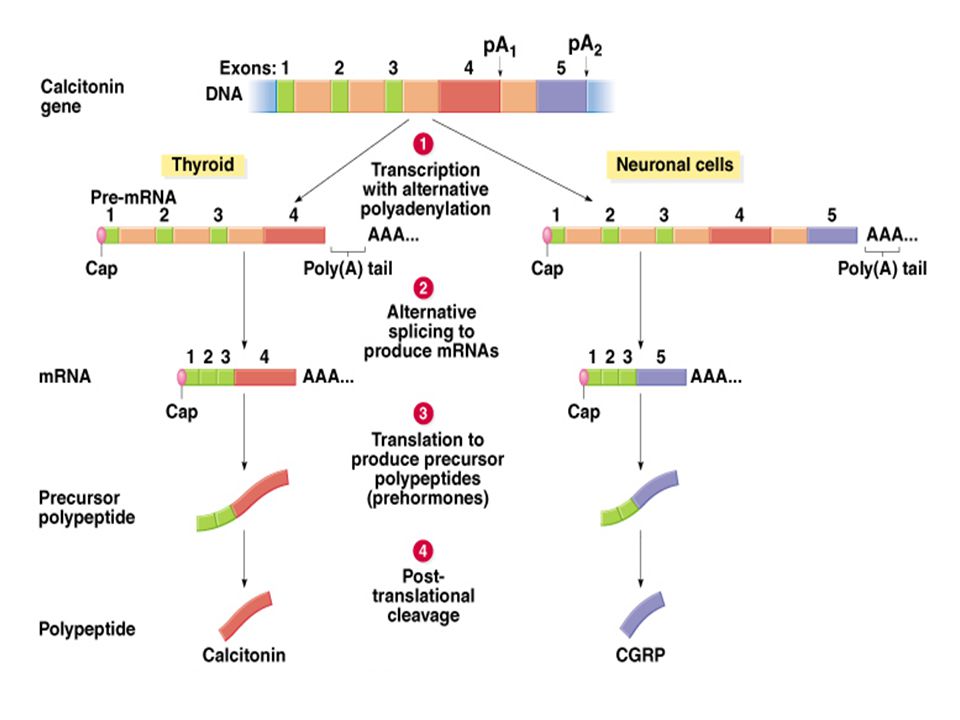
Az RNS processzálódás szabályozza az mRNS képződését a prekurzor RNS-ekből. Két független szabályozó mechanizmus van: Alternatív poladeniláció = amikor a polyA farok szintetizálódik Alternatív splicing = amikor az exonok vágódnak ki Az alternatív pliadeniláció és splicing együtt is előfordulhatnak. példák: Humán kalcitonin (CALC) gén a thyroid (pajzsmirigy) és a neuron sejtekben Ivar meghatározás Drosophila-ában
 gén a thyroid (pajzsmirigy) és a neuron sejtekben. Ivar meghatározás Drosophila-ában.")
23
Az ivar meghatározása Drosophila-ban alternatív splicing-gal.
Az ivart az X:A arány határozza meg. (X=kromoszóma; A=allél) Sxl (sex lethal) gén határozza meg az utat a hímeknek és a nőstényeknek. Ha X:A = 1, az összes intron és az exon 3 (ami stop kodont tartalmaz) is kivágódik. Ha X:A = 0.5, nem képződik funkcionális fehérje. Átíródik, jó splicing Átíródik, rossz splicing Jó Sxlp Rossz fehérje Jó tra splicing Rossz alternatív Trap- Trap+ Trap+Tra-2p női dsx splicing Dsx fiú splicing Fiú diff gátolt Leány diff.
 Sxl (sex lethal) gén határozza meg az utat a hímeknek és a nőstényeknek. Ha X:A = 1, az összes intron és az exon 3 (ami stop kodont tartalmaz) is kivágódik. Ha X:A = 0.5, nem képződik funkcionális fehérje. Átíródik, jó splicing. Átíródik, rossz splicing. Jó Sxlp. Rossz fehérje. Jó tra splicing. Rossz alternatív. Trap- Trap+ Trap+Tra-2p női dsx splicing. Dsx fiú splicing. Fiú diff. gátolt Leány diff.")
24
mRNS transzport szabályozás:
Az eukarióta mRNS transzport szabályozott. Néhány kísérlet azt mutatta, hogy az elsődleges transzkriptimok ~1/2-e soha nem hagyja el a sejtmagot és lebontódik. Az érett mRNS-ek a nukleáris pórusokon keresztül távoznak.

25
mRNS transzlációs szabályozás:
Például a megtermékenyítetlen petében az mRNS-ek (a petében tárolt /nincs új mRNS szintézis) transzlációja a megtermékenyítés után azonnal megindul. A tárolt mRNS-eket védő fehérjék akadályozzák a transzlációt. Poli(A) farok segíti a transzlációt. A tárolt mRNS-eknek általában rövid poli(A) farka van. (15-90 A vs A). Specifikus mRNS-ek kijelölődnek a dezadenilációra (“tail-chopping”) a betárolás előtt az AU-gazdag 3’-UTR szekvenciáik alapján. Akkor aktiválódnak, ha egy enzim felismeri az AU-gazdag szekvenciát és még hozzáad ~150 A-t és a ploi(A) farok eléri teljes hosszúságát.
 transzlációja a megtermékenyítés után azonnal megindul. A tárolt mRNS-eket védő fehérjék akadályozzák a transzlációt. Poli(A) farok segíti a transzlációt. A tárolt mRNS-eknek általában rövid poli(A) farka van. (15-90 A vs A). Specifikus mRNS-ek kijelölődnek a dezadenilációra ( tail-chopping ) a betárolás előtt az AU-gazdag 3’-UTR szekvenciáik alapján. Akkor aktiválódnak, ha egy enzim felismeri az AU-gazdag szekvenciát és még hozzáad ~150 A-t és a ploi(A) farok eléri teljes hosszúságát.")
26
mRNS degradációs szabályozás:
A citoplazmában minden mRNS ki van téve degradációnak. A tRNS-ek és az rRNS-ek általában nagyon stabilak; mRNS-ek stabilitása változó (percek, hónapok). A stabilitás a szabályozó szignálok hatására változhat és úgy vélik, hogy ez a legfontosabb szabályozó pont. Különböző szekvenciák és folyamatok befolyásolják az mRNS félélet-idejét: AU-gazdag elemek Másodlagos szerkezet Dezadenilációs enzimek 5’ de-capping Vágás az mRNS belsejében és a fragmentumok degradációja.
. A stabilitás a szabályozó szignálok hatására változhat és úgy vélik, hogy ez a legfontosabb szabályozó pont. Különböző szekvenciák és folyamatok befolyásolják az mRNS félélet-idejét: AU-gazdag elemek. Másodlagos szerkezet. Dezadenilációs enzimek. 5’ de-capping. Vágás az mRNS belsejében és a fragmentumok degradációja.")
27
Poszt-transzlációs szabályozás - protein degradáció:
A fehérjék lehetnek rövid-(pl. szteroid receptorok) vagy hosszú-életűek (pl. a szemlencse fehérjéi). A fehérjék degradációjához az eukariótákban szükséges az ubiquitin kofaktor. Az ubiquitin kötődik a fehérjékhez és megjelöli azokat a proteolítikus enzimeknek. Az N-terminális aminosavtól függ a fehérje stabilitása és meghatározza az ubiquitin kapcsolódás mértékét. Arg, Lys, Phe, Leu, Trp /2 élet ≤3 perc Cys, Ala, Ser, Thr, Gly, Val, Pro, Met 1/2 élet ≥ 20 óra
 vagy hosszú-életűek (pl. a szemlencse fehérjéi). A fehérjék degradációjához az eukariótákban szükséges az ubiquitin kofaktor. Az ubiquitin kötődik a fehérjékhez és megjelöli azokat a proteolítikus enzimeknek. Az N-terminális aminosavtól függ a fehérje stabilitása és meghatározza az ubiquitin kapcsolódás mértékét. Arg, Lys, Phe, Leu, Trp 1/2 élet ≤3 perc. Cys, Ala, Ser, Thr, Gly, Val, Pro, Met 1/2 élet ≥ 20 óra.")
28
Hosszú távú génszabályozás - fejlődés és differenciálódás:
Egy adott genotípusú, egyetlen egy sejtből számos specializált szövet és szerv képződik. Fejlődés A fejlődés a genom kölcsönhatása a citoplazmával és a külső környezettel, hogy egy programozott lépéssorral jellemző irreverzibilis események jöjjenek létre. Differenciálódás: Differenciálódással sejttípusok, szövetek és szervek jönnek létre specifikus génszabályozással. A fejlődés és a differenciálódás több szinten tanulmányozható: Morfológia Biokémia Aktivációs-repressziós mintázat

29
Genom aktivitás Az eukarióta DNS nagy része nem kódol fehérjét. Tengeri sün, az egyedi szekvenciák ~6%-a aktív egyszerre egy időben. Az érett szövetekben az expresszió ~0,8%-ra csökken. A nem-átíródó DNS legnagyobb részének funkciója nem ismert.

30
A DNS állandósága a fejlődés alatt
A felnőtt sejtjeinek ugyanolyan a genomja, mint a zigótának? Frederick Steward (1950) sikeresen növesztett új sárgarépát a növény szárszövetéből. Ian Wilmut és mtsi (1997) birkát klónoztak. 1997 óta egeret, marhát, majmot és humán embriósejtet klónoztak. A differenciálódott szomatikus sejt nukleusza totipotens; mindazzal a genetikai információval rendelkezik, ami a fejlődéshez szükséges.
 sikeresen növesztett új sárgarépát a növény szárszövetéből. Ian Wilmut és mtsi (1997) birkát klónoztak óta egeret, marhát, majmot és humán embriósejtet klónoztak. A differenciálódott szomatikus sejt nukleusza totipotens; mindazzal a genetikai információval rendelkezik, ami a fejlődéshez szükséges.")
31
A fejlődés és a differenciálódás modell szervezetei:
Drosophila melanogaster (ecet muslica) Régóta vizsgált. Sok, a fejlődést befolyásoló mutáció azonosítottak. Caenorhabditis elegans (nematóda, fonalféreg) Kis genom (97 Mb) A testi sejtek száma és a leszármazása ismert. A genetikai keresztezés és az öntermékenyítés könnyű. A test áttetsző. Brachydanio rerio (zebradánió) Az embriók áttetszőek. Nagyszámú hal tenyészthető. Screening technikák jól kidolgozottak. Arabidopsis thaliana (növény, libatop) Kicsi, könnyű keresztezni és elemezni nagyszámú utódot. Sok fejlődési mutánst azonosítottak.
 Régóta vizsgált. Sok, a fejlődést befolyásoló mutáció azonosítottak. Caenorhabditis elegans (nematóda, fonalféreg) Kis genom (97 Mb) A testi sejtek száma és a leszármazása ismert. A genetikai keresztezés és az öntermékenyítés könnyű. A test áttetsző. Brachydanio rerio (zebradánió) Az embriók áttetszőek. Nagyszámú hal tenyészthető. Screening technikák jól kidolgozottak. Arabidopsis thaliana (növény, libatop) Kicsi, könnyű keresztezni és elemezni nagyszámú utódot. Sok fejlődési mutánst azonosítottak.")
32
A Drosophila fejlődés genetikai szabályozása:
A Drosophila fejlődés stádiumai: (10-12 nap) Tojás Lárva (3 instars) Báb Felnőtt
 Tojás. Lárva (3 instars) Báb. Felnőtt.")
33
A Drosophila embrió fejlődése:
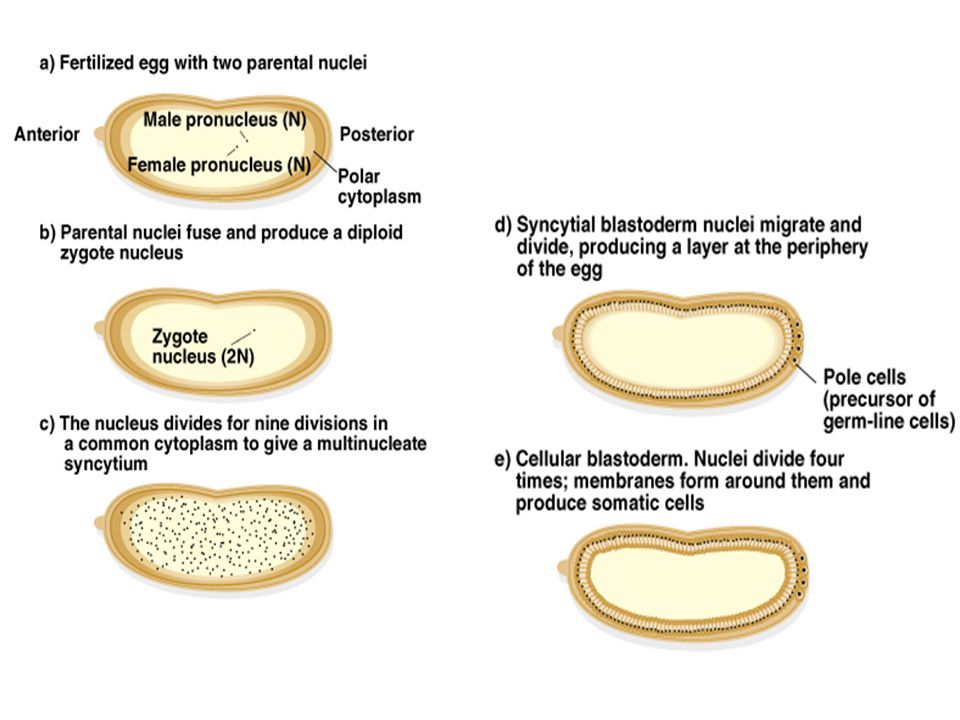
A fejlődés a megtermékenyítéssel kezdődik. A megtermékenyítés előtt, a petében molekuláris gradiens van. Poláris citoplazma alakul ki a posterior végén. A 2 nukleusz fuzionál a megtermékenyítés után és zigóta képződik. 9 mitotikus osztódás lesz sejtosztódás nélkül és 7 osztódás után néhány nukleusz a poláris citoplazmához vándorol (posterior) és kialakítja a csíra-vonal prekurzorokat. Más nukleuszok a sejtfelülethez vándorolnak és kialakul a blastoderma prekurzor. 4 további mitotikus osztódás az összes sejtmag elkülönül egymástól sejtmembránnal.
 és kialakítja a csíra-vonal prekurzorokat. Más nukleuszok a sejtfelülethez vándorolnak és kialakul a blastoderma prekurzor. 4 további mitotikus osztódás az összes sejtmag elkülönül egymástól sejtmembránnal.")
35
A fejlődés egymást követő lépései két folyamattól függenek:
Az anterior-posterior és a dorsal-ventral molekuláris gradiensektől . A (1) paraszegmentumok és az (2) embrionális szegmentumok képződése, amelyek végül a (3) felnőtt szegmentumok képződéséhez vezetnek. A felnőtt szelvényezettsége az embrióénak felel meg.
 paraszegmentumok és az (2) embrionális szegmentumok képződése, amelyek végül a (3) felnőtt szegmentumok képződéséhez vezetnek. A felnőtt szelvényezettsége. az embrióénak felel meg.")
36
A gének három fő csoportja szabályozza a fejlődést és a differenciálódást
*Letális és abnormális szerkezetek a fejlődés közben. Anyai hatású gének Szegmentáció gének Homeotikus gének

37
1. Anyai hatású gének (az anya készítette elő)
bicoid gén Az anterior szerkezet kialakulását szabályozza (a mutánsoknak mindkét végükön posterior szerkezet van). A gén a peteképződés közben íródik át és a megtermékenyítés után nyilvánul meg. A bicoid géntermék fehérje gradienst alakít ki. nanos gén A potroh képződést szabályozza (mRNS-ek a pete posterior részén gyűlnek össze). A nanos gén szintén fehérje gradienst alakít ki. torso gén Transzkripció és transzláció a peteképződés közben. A petében mindenhol van, de csak a plólusokon aktív.
. A gén a peteképződés közben íródik át és a megtermékenyítés után nyilvánul meg. A bicoid géntermék fehérje gradienst alakít ki. nanos gén. A potroh képződést szabályozza (mRNS-ek a pete posterior részén gyűlnek össze). A nanos gén szintén fehérje gradienst alakít ki. torso gén. Transzkripció és transzláció a peteképződés közben. A petében mindenhol van, de csak a plólusokon aktív.")
38
~25 szegmentációs gént azonosítottak és osztottak 3 csoportba:
2. Szelvényezettség gének (az embriót régiókra osztja): ~25 szegmentációs gént azonosítottak és osztottak 3 csoportba: Gap gének – a mutációjuk eredményeként néhány szomszédos szelvény deléciója. Pair rule gének – mutációjuk minden szegmensben a mintázat ugyanazon részének delécióját okozza. Szegmens polaritás gének – mutánsoknál a szegmens egy része kicserélődik a kapcsolódó fél-szegmens tükörképével.
: ~25 szegmentációs gént azonosítottak és osztottak 3 csoportba: Gap gének – a mutációjuk eredményeként néhány szomszédos szelvény deléciója. Pair rule gének – mutációjuk minden szegmensben a mintázat ugyanazon részének delécióját okozza. Szegmens polaritás gének – mutánsoknál a szegmens egy része kicserélődik a kapcsolódó fél-szegmens tükörképével.")
39
A szelvényezettség gének szerepe mutánsokkal meghatározva

40
3. Homeotikus gének (a szerkezetet határozzák meg):
A homeotikus gének minden egyes szelvénynél meghatározzák a testrész fejlődését. A felnőtt testrészei nem-differenciálódott lárva szövetekből, az imágó diszkekből fejlődnek ki. A homeotikus mutánsok más testrészt növesztenek, mint a normális testrész egy bizonyos szelvényen. A különböző homeotikus géncsoportoknak van azonos szekvenciája ~180 bp, a homeoboxok, amik fehérjét kódolnak. A homeoboxok szabályozzák a fejlődést és a termelt fehérjék a gén egységek upstream régiójához kötődnek. A homeotikus génkomplexeket Hox-nak rövidítjük. Hox gének gerincesekben és növényekben is.

41
A homológ imágó diszkek elhelyezkedése a lárvában és felnőttben.

42
Homeotikus mutánsok bithorax homeotikus mutáns antennapedia 4-szem

43
A bithorax homeotikus gének szerveződése Drosophila-ában, ~300 kb.

Hasonló előadás





 DNS-ből,>")


 daganatkeltő.>")
 megváltozHAT Ebből visszakövetkeztethetünk a mutációt szenvedett gén funkciójára.>")


 prokariótában>")




>")
 Bihari Péter.>")
 Csernetics Árpád Bioinformatika SZIT 2005. ápr. 18.>")



