Előadást letölteni
Az előadás letöltése folymat van. Kérjük, várjon

1
Az elektrontranszportlánc működése
Fotoszintézis II. Az elektrontranszportlánc működése

2
Energiaszint változások a fotoszintézis folyamán

3
14-47 Energiaszint változások a fotoszintézis folyamán
Minél nagyobb az elektron energiatartalma annál negatívabb a redox potenciálja Az első fényreakció a PS II-ben történik (680 nm) Az elektron energiája csökken az elektrontranszport során Újragerjesztés a PS I-ben (700 nm). Energia tárolás ATP és NADPH formájában.
 Az elektron energiája csökken az elektrontranszport során. Újragerjesztés a PS I-ben (700 nm). Energia tárolás ATP és NADPH formájában.")
4
14-47 Az elektrontranszportlánc két formája
A nem ciklusos elektrontranszport (Z séma) A ciklikus variációban a ferredoxin a citokróm b6/f komplexnek adja át az elektront. A fény ily módon a PSI egyedüli részvételével „körbe hajtja” az elektronokat és így hoz létre H+ koncentráció-különbséget a sztróma és a luminális tér között. Ez ATP szintézisre használódik fel.
 A ciklikus variációban a ferredoxin a citokróm b6/f komplexnek adja át az elektront. A fény ily módon a PSI egyedüli részvételével „körbe hajtja az elektronokat és így hoz létre H+ koncentráció-különbséget a sztróma és a luminális tér között. Ez ATP szintézisre használódik fel.")
5
A tilakoidmembránok makromolekuláris elemeinek elhelyezkedése

6
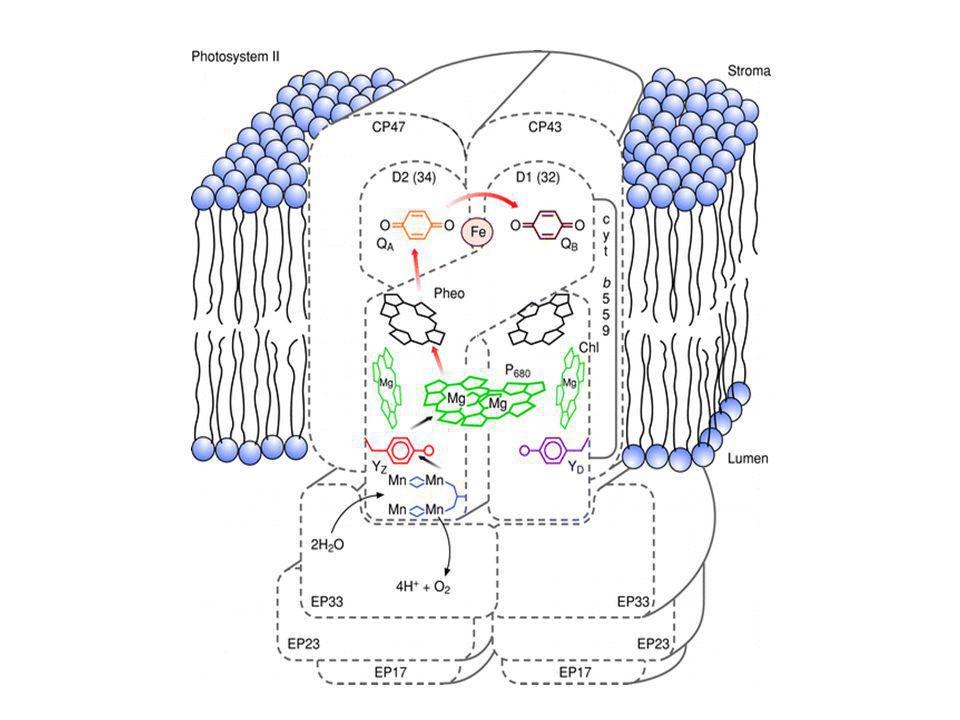
6. A II. fotokémiai rendszer működése. Az e- útja a víztől a PQ pool-ig.
A PS II felépítése 1.) A reakciócentrum: D1 és D2 fehérjék, P680, 3-4 kla, 1 β-karotin, 2 feofitin QA és QB (elsődleges és másodlagos akceptorok) D aminosava, tirozin; a Z elsődleges e- donor 2 db citokróm b559, fotoprotektív szerep Mn atomok - vízbontás ("M" enzim) klorid, kalcium további 7 fehérje
 A reakciócentrum: D1 és D2 fehérjék, P680, 3-4 kla, 1 β-karotin, 2 feofitin. QA és QB (elsődleges és másodlagos akceptorok) D aminosava, tirozin; a Z elsődleges e- donor. 2 db citokróm b559, fotoprotektív szerep. Mn atomok - vízbontás ( M enzim) klorid, kalcium. további 7 fehérje.")
7
A PS II felépítése

9
A PS II belső elektrontranszportja

10
A PS II energia szint változásai és kinetikai paraméterei

11
A II. fotokémiai rendszer működése. Az e- útja a víztől a PQ pool-ig
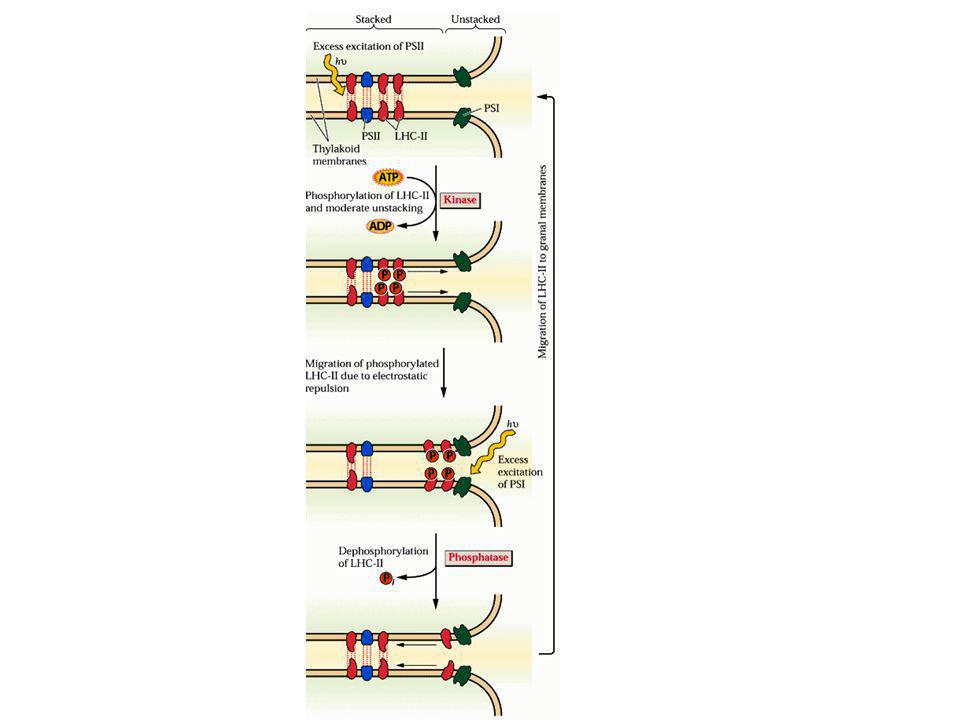
2.) A reguláló sapka 33, 23, 16 kDa proteinek a lumen felőli oldalon 3.) A proximális antenna CP47, CP43 kl-protein komplexek (25 kla és 5 β-karotin) 4.) A disztális antenna LHC-II és járulékos kl-proteinek Kla/b = 1.2, 200 db; xantofill LHC-II szerepe a fényenergia megosztásában a PSII/PSI között foszforilálódás/defoszforilálódás)
 A reguláló sapka. 33, 23, 16 kDa proteinek a lumen felőli oldalon. 3.) A proximális antenna. CP47, CP43 kl-protein komplexek (25 kla és 5 β-karotin) 4.) A disztális antenna. LHC-II és járulékos kl-proteinek. Kla/b = 1.2, 200 db; xantofill. LHC-II szerepe a fényenergia megosztásában a PSII/PSI között foszforilálódás/defoszforilálódás)")
12
Az LHC II szabályozza az energiamegoszlást a PS II és PS I között

14
A II. fotokémiai rendszer működése. Az e- útja a víztől a PQ pool-ig
A primér töltésszeparáció (3 ps, P680) feofitin -- QA (300 ps) QA -- QB (1.: 0.2 ms; 2.: 0.5 ms) -- PQH2 gyors és lassú fluoreszcencia indukciós görbék (fluoreszcencia hozam változások)
 feofitin -- QA (300 ps) QA -- QB (1.: 0.2 ms; 2.: 0.5 ms) -- PQH2. gyors és lassú fluoreszcencia indukciós görbék. (fluoreszcencia hozam változások)")
15
Kautsky effektus I. fluoreszcencia indukció, indukált fluoreszcencia

16
Kautsky effektus II. A fluoreszcencia indukció és kioltás szakaszai
F0 = minimális fluoreszcencia Fm = maximális fluoreszcencia Fv = változó fluoreszcencia Fv = Fm - F0 ; Fv/Fm max. 0,84 D → P; PSII → PSI e- átmenet Fluoreszcencia kioltás: fotokémiai és nem-fotokémiai P → M → T szakaszok

17
Kautsky effektus III. (fluoreszcencia indukció, indukált fluoreszcencia)
")
18
Kautsky effektus IV. A gyors fluoreszcencia indukció: az O-J-I-P tranziens
30 ms 2 ms

19
A DCMU a QA és QB között gátol: minden QA redukálódik

20
Kautsky effektus IV. A gyors fluoreszcencia indukció: az O-J-I-P tranziens

21
Fluoreszcencia analízis Kioltási (quenching) analízis
 analízis")
22
Fluoreszcencia analízis

23
Fluoreszcencia analízis
F0 = a sötét-adaptált minta minimális fluoreszcenciája Fm = a sötét-adaptált minta maximális fluoreszcenciája ugyanezek , - vel jelölve a fényadaptált mintára nézve Ft = a fluoreszcencia egy adott „t” időpontban Y(ield) = (Fm, - Ft) : Fm, ; a PSII átlagos kvantumhatásfoka ETR = Y x PAR x 0,5 x 0,84 ; az elektrontranszport látszólagos sebessége qP = (Fm, - Ft) : (Fm, - F0,) fotokémiai kioltási koefficiens qN = (Fm – Fm,) : (Fm - F0,) nem-fotokémiai kioltási koeff. NPQ = (Fm – Fm,) : Fm, nem-fotokémiai kioltás
 = (Fm, - Ft) : Fm, ; a PSII átlagos kvantumhatásfoka. ETR = Y x PAR x 0,5 x 0,84 ; az elektrontranszport látszólagos sebessége. qP = (Fm, - Ft) : (Fm, - F0,) fotokémiai kioltási koefficiens. qN = (Fm – Fm,) : (Fm - F0,) nem-fotokémiai kioltási koeff. NPQ = (Fm – Fm,) : Fm, nem-fotokémiai kioltás.")
24
A II. fotokémiai rendszer működése. Az e- útja a víztől a PQ pool-ig
Vízbontás: az Mn-enzim S0 -- S4 állapotai az oxigén termelés négyes periódusa 2H2O O2 + 4H+ + 4e- protonok a lumenbe távoznak

25
A víz bontása: a reguláló sapka szerepe

26
P. Joliot és B. Kok impulzus-megvilágításos kísérlete (1975)
")
27
A vízbontó enzim állapotváltozásai
S0 : S1 : S2 : S3 = 0,25 : 0,75 : 0 : 0

28
A Mn szerepe

29
A plasztokinon mint mobilis komponens

30
A plasztokinon mint mobilis komponens

31
Az e- útja a PQ pool-tól a NADPH-ig
A citokróm b6/f komplex működése, felépítése oxidálja a PQH2-t, redukálja a PC-t, H+-t transzlokál a lumenbe oxidálja a sztromában lévő ferredoxint (ciklikus e-tr. !) 4 polipeptid eqiumolárisan; cit-f, cit-b6 (2 hem); Rieske-f. vas-kén; 17 kDa protein 2 e- t ad tovább a P-700 felé 4 H+-t a sztrómából a lumenbe a ciklikus e- transzport növeli a H+ transzportot két kötőhely, 2 ciklus A plasztocianin 10.5 kDa Cu-protein lumenben
 4 polipeptid eqiumolárisan; cit-f, cit-b6 (2 hem); Rieske-f. vas-kén; 17 kDa protein. 2 e- t ad tovább a P-700 felé. 4 H+-t a sztrómából a lumenbe. a ciklikus e- transzport növeli a H+ transzportot. két kötőhely, 2 ciklus. A plasztocianin kDa Cu-protein. lumenben.")
32
A citokróm b6/f komplex molekuláris felépítése

33
A citokróm b6/f komplex működése oxidálja a PQH2-t, redukálja a PC-t
a ciklusos elektrontranszportban oxidálja a ferredoxint proton átadást közvetít a sztrómából a lumenbe Fd Fd -50 mV -150 mV -50 mV -150 mV

34
DCMU: 3-(3,4-diklorofenil)-1,1-dimetilurea
DBMIB: 2,5-dibróm-3-metil-6-izopropil-p-benzokinon Paraquat (metil viologén)
")
35
7. Az I. fotokémiai rendszer működése.
Reakciócentrum: heterodimer kDa fehérje P-700; A0 (10 ps);, A1 (50 ps): elsődleges, másodlagos e-akceptorok (kla; ill. K-vitamin- fillokinon) Vas-kén centrumok: FX, FB, FA (4Fe-4S) PC, ferredoxin-kötő fehérjék Ferredoxin: e- transzport a ferredoxin-NADP+-oxidoreduktáz (FNO) felé; vagy vissza a cit b6/f komplex felé (ciklikus e- - transzport)
;, A1 (50 ps): elsődleges, másodlagos e-akceptorok. (kla; ill. K-vitamin- fillokinon) Vas-kén centrumok: FX, FB, FA (4Fe-4S) PC, ferredoxin-kötő fehérjék. Ferredoxin: e- transzport a ferredoxin-NADP+-oxidoreduktáz (FNO) felé; vagy vissza a cit b6/f komplex felé (ciklikus e- - transzport)")
36
A PS I felépítése Az I. fotokémiai rendszer Reakciócentrum:
heterodimer kDa fehérje P-700; A0 (10 ps);, A1 (50 ps): elsődleges, másodlagos e-akceptorok (kla; ill. K-vitamin- fillokinon) Vas-kén centrumok: FX, FB, FA (4Fe-4S)
;, A1 (50 ps): elsődleges, másodlagos e-akceptorok. (kla; ill. K-vitamin- fillokinon) Vas-kén centrumok: FX, FB, FA (4Fe-4S)")
37
A PS I felépítése

38
A PS I. felépítése és működése
Reakciócentrum: 2 db centrális fehérje P700 klorofilla elsődleges (Kla) és másodlagos (K-vitamin) akceptorok vas-kén centrumok PC és ferredoxin kötő fehérjék Nem ciklusos elektrontranszport: az e- a NADP+ felé, Ciklusos elektrontranszport: e- a citokróm b6/f komplex felé Ferredoxin, ferredoxin-NADP+ reduktáz, NADP+ Eredmény: NADPH + H+
 és másodlagos (K-vitamin) akceptorok. vas-kén centrumok. PC és ferredoxin kötő fehérjék. Nem ciklusos elektrontranszport: az e- a NADP+ felé, Ciklusos elektrontranszport: e- a citokróm b6/f komplex felé. Ferredoxin, ferredoxin-NADP+ reduktáz, NADP+ Eredmény: NADPH + H+")
39
A PS I.belső eletrontranszportja

40
A PS I felépítése molekuláris dimenziók…

41
A ciklikus elektron transzportlánc működése

42
Az elektrontranszport lánc specifikus gátlószerei gyakran fotoszintézisgátló növényvédőszerként kerülnek forgalomba DCMU: jelenleg is használatos Paraquat: kivonták a forgalomból

43
Az elektron transzportlánc inhibitorainak hatóhelyei

44
Ferredoxin-NADP reduktáz
LUMEN SZTRÓMA

45
Ferredoxin-NADP+-oxidoreduktáz
33 kD protein (sejtmagban kódolt) tilakoid sztróma felőli oldalán
 tilakoid sztróma felőli oldalán.")
46
A kemiozmotikus mechanizmus. Fotofoszforiláció.
Peter Mitchell 1963; Nobel díj 1978: Kapcsolódás a H+ elektrokémiai potenciálgradiens és a sejt munkavégzése között szelektíven permeábilis membránon keresztül: μH = p = ψ – 2.3RTpH /F elektrokémiai proton membrán pH gradiens H+ gradiens mozgató potenciál p = ψ – 59pH (mV) ATP szintáz (F0-F1 típusú ATPáz) – fotofoszforiláció
 ATP szintáz (F0-F1 típusú ATPáz) – fotofoszforiláció.")
47
8. A fotoszintetikus foszforiláció: a Mitchell-f
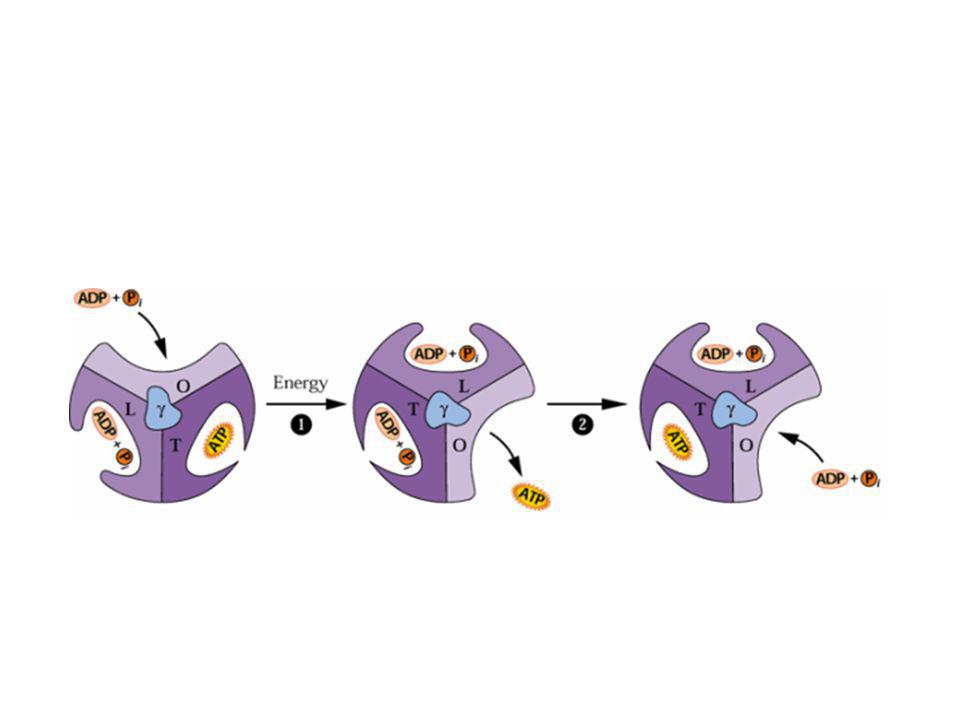
8. A fotoszintetikus foszforiláció: a Mitchell-f. kemiozmotikus mechanizmus Jagendorf: az ATP képződés mechanizmusa, bizonyítás CF0/CF1 ATP szintáz felépítése ATP szintézis

48
Az ATP szintáz „F0F1-ATPáz”

49
A fotoszintetikus foszforiláció: kemiozmotikus modell
proton elektrokémiai potenciálgrádiens a lumen és a sztróma között: ezt használja fel az ATP- szintáz enzim az ATP szintézisére ADP-ből és anorganikus foszfátból

51
A H+-koncentrációkülönbség és az ATP képződés
A két folyamat a mitokondriumban és a kloroplasztiszban hasonló módon játszódik le. Mindkét esetben az ATP szintézis egy protontranszlokáló enzim, az ATP szintáz, segítségével megy végbe. A protonáramlás a magasabb, lúgosabb pH felé tart Az enzim aktív helyei a „gomba fejében” helyezkednek el. Az ATP a mitokondriumi matrixban illetve a kloroplasztisz beli sztrómában keletkezik.

52
P N Baktérium Kívül Belül (citoplazma) Mitokondrium Intermembrán tér
A protongradiens irányítottsága különböző rendszerekben: („protokémiailag” pozitív : P oldal „protokémiailag” negatív: N oldal) P N Baktérium Kívül Belül (citoplazma) Mitokondrium Intermembrán tér Matrix Kloroplasztisz Lumen Sztróma
 P. N. Baktérium. Kívül. Belül (citoplazma) Mitokondrium. Intermembrán tér. Matrix. Kloroplasztisz. Lumen. Sztróma.")
53
Az elektontranszportlánc és a foszforiláció
A két folyamat között olyan szoros a kapcsolat hogy ATP-képződés nélkül az elektrontanszportlánc leáll és fordítva. Szétkapcsoló anyagok megszüntetik ezt a szoros kapcsolatot. Ilyen szétkapcsoló anyagok a következők: CCCP, valinomicin, gramicidin D, DCPIP stb. CCCP = karbonilcianid m-klorofenil hidrazon DCPIP = 2,6-diklorofenol indofenol

54
ÖSSZEFOGLALÁS

55
Köszönöm a figyelmet!

Hasonló előadás