Előadást letölteni
Az előadás letöltése folymat van. Kérjük, várjon

1
A fényenergia hasznosítása
Fotoszintézis A fényenergia hasznosítása

2
A fotoszintézis két fő szakasza: a fényreakciók és a sötét reakció

3
14-33 A folyamat jelentősége
Az élővilág energia -forrása Szerves anyagok előállítása szervetlenekből Az ábrán a keményítő szemcsék láthatók A Föld légkörének oxigéntartalma ebből a folyamatból ered

5
1. Fényszakasz: A fényt fotoszintetikus pigmentek abszorbeálják
A fényt abszorbeáló pigmentek a cianobaktériumokban és magasabbrendű növényekben: Kolorofill a, b Karotinoidok Fikobilinek (cianobaktériumokban és vörösmoszatokban)
")
6
A prizmával alkotóelemeire bontott fehér fény kék és vörös hullámhosszú komponenseivel megvilágított régió köré gyűlnek az aerob baktériumok a fotoszintetizáló Spirogyra körül

7
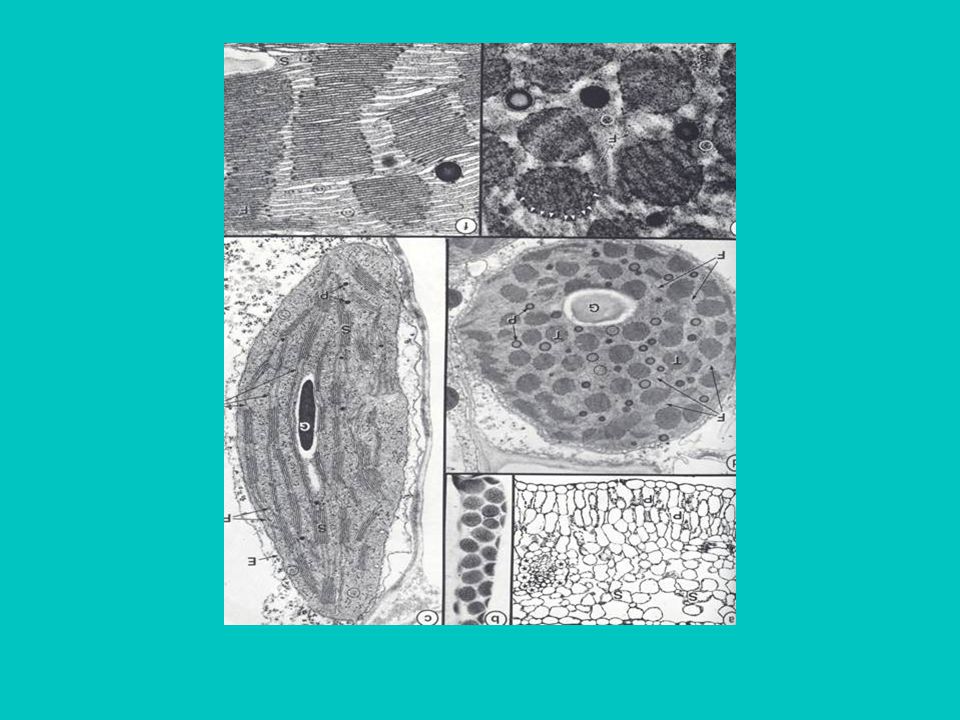
A levél keresztmetszete

8
14-35b A kloroplasztisz szerkezete
Kettős borítómembrán határolja Folyékony közege a sztróma –benne enzimek, keményítőszemcsék, DNS és riboszómák Membránrendszere tilakoidokból áll– ezek helyenként gránumokba rendeződik A tilakoidok belső üregei egy összefüggő, lumen nevezetű hálózatot alkotnak

10
14-41 A fénybegyűjtő pigmentek: Klorofill
Porfirin gyűrű – négy pirrolgyűrűből áll A négy pirrolgyűrű mellett egy öttagú ciklopentanon gyűrű található A konjugációs rendszer kékkel jelölve Magnézium ion – Mg++ - narancs Észter kötéssel csatlakozó fitol – zöld Ez a klorofilla – a többi klorofill kicsit külőnbözik.

11
Klorofillok

12
A karotinoidok járulékos pigmentek

13
A pigment-protein komplexek rendezetten helyezkednek a tilakoidmembránban

14
A fényenergia abszorpciója

15
A fény abszorpciója és a gerjesztési energia sorsa
Fotoszintetikusan aktív fény ~ nm-ig Klorofill molekula: alapállapot (S0), Egy p- elektron kötő pályáról lazító pályára lép. szinglet (S1 és S2) és triplet (T1) állapotok vörös fény elnyelése: S állapot kék fény elnyelése: S2 állapot A gerjesztési energia sorsa: hővé alakul, fluoreszcencia, fotokémiai reakciók (redox folyamatok) energia migráció vagy energia transzfer (azonos vagy kémiailag eltérő pigmenteknek adódik át az energia)
, Egy p- elektron kötő pályáról lazító pályára lép. szinglet (S1 és S2) és. triplet (T1) állapotok. vörös fény elnyelése: S1 állapot. kék fény elnyelése: S2 állapot. A gerjesztési energia sorsa: hővé alakul, fluoreszcencia, fotokémiai reakciók (redox folyamatok) energia migráció vagy energia transzfer. (azonos vagy kémiailag eltérő pigmenteknek adódik át az energia)")
16
A klorofill fluoreszcenciája akkor is a vörös hullámhossz tartományba esik, ha kék fényt nyel el
Ok: az S2-S1 átmenet mindig hő formájában disszipálódik

17
A fluoreszcencia imázs analízis a lézerrel egy ponton gerjesztett fluoreszcencia terjedését mutatja a levél felszínén (a megvilágított minták effektív kvantumhatékonyságának a képe)
")
18
A növényzet fluoreszcenciája műholdas felvételeken is detektálható
Kaliforniai öböl, algavirágzás, október 6., számítógépes színek, a kék az alacsony, a vörös a nagy intenzitású fluoreszcencia

19
Az antennapigmentek az elnyelt fényenergiát a reakciócentrum klorofilla felé továbbítják
A fényenergia vándorlása: antenna pigmentek között: külső és belső antennák reakciócentrumok klorofill a molekulájához: II. fotokémiai rendszer: P680, I. fotokémiai rendszer: P700

20
Energiaátadás két pigment molekula között akkor történik, ha a donor molekula abszorpciós vagy fluoreszcencia spektruma részben fedi egymást

21
14-43 A fénygyűjtő komplex és a reakciócentrum
A fénygyűjtő komplex (LHC II. és I.) a kísérőpigmentek által elnyelt fényenergiát a reakciócentrum klorofillokhoz közvetíti. A fénybegyűjtő komplexek pigmentjei magasabbrendű növényekben klorofill a és b, karotin és xantofill molekulák, gyakran fehérjékhez kötődve. A gerjesztett pigmentek energiája rezonancia által átadódik a reakciócentrumba. A fényenergia kémiai energiává alakulása: töltésszeparációval Donor Kla Akceptor; Donor Kla Akceptor Donor Kla Akceptor; Donor Kla Akceptor
 a kísérőpigmentek által elnyelt fényenergiát a reakciócentrum klorofillokhoz közvetíti. A fénybegyűjtő komplexek pigmentjei magasabbrendű növényekben klorofill a és b, karotin és xantofill molekulák, gyakran fehérjékhez kötődve. A gerjesztett pigmentek energiája rezonancia által átadódik a reakciócentrumba. A fényenergia kémiai energiává alakulása: töltésszeparációval. Donor Kla Akceptor; Donor Kla Akceptor. Donor Kla Akceptor; Donor Kla Akceptor")
22
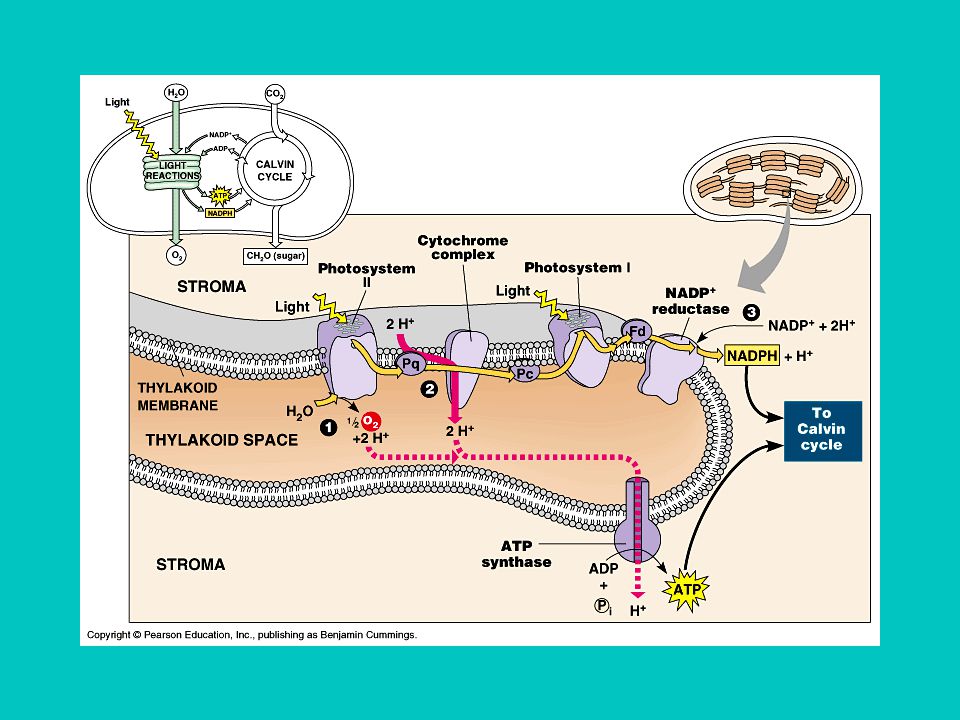
14-47 Energiaszint változások a fotoszintézis folyamán
Minél nagyobb az elektron energiatartalma annál negatívabb a redox potenciálja Az első fényreakció a PS II ben történik (680 nm) Az elektron energiája csökken az elektrontranszport során Újragerjesztés a PS I-ben (700 nm). Energia tárolás ATP és NADPH formájában.
 Az elektron energiája csökken az elektrontranszport során. Újragerjesztés a PS I-ben (700 nm). Energia tárolás ATP és NADPH formájában.")
23
14-47 Az elektrontranszportlánc két formája
A nem ciklusos elektrontranszport (Z séma) A ciklikus variációban a ferredoxin a citokróm b6/f komplexnek adja át az elektront. A fény ily módon a PSI egyedüli részvételével körbe hajtja az elektronokat és így hoz létre H+ koncentráció-különbséget a sztróma és a luminális tér között. Ez ATP szintézisre használódik fel.
 A ciklikus variációban a ferredoxin a citokróm b6/f komplexnek adja át az elektront. A fény ily módon a PSI egyedüli részvételével körbe hajtja az elektronokat és így hoz létre H+ koncentráció-különbséget a sztróma és a luminális tér között. Ez ATP szintézisre használódik fel.")
25
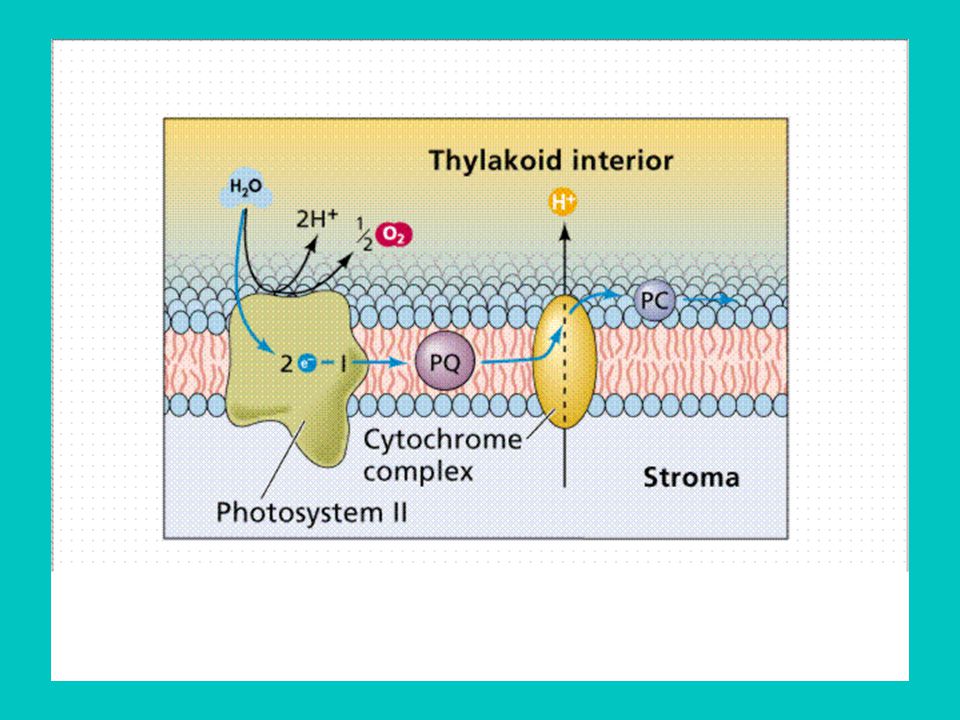
1. A PS II felépítése - reakciócentrum: D1 és D2 fehérjék: ehhez kapcsolódnak a feofitin (primer e- akceptor), a QA és QB kinon akceptorok, D1 fehérje tirozin aminosava (primer e- donor) Mn2+, Mn3+ ionok (vízbontás) - reguláló sapka - proximális antenna - disztális antenna 2. Funkciói vízbontás: 2H2O O2 +4H+ + 4e- proton keletkezik a lumenben, elektron lép az e- transzportláncba a plasztokinon redukciója
 Mn2+, Mn3+ ionok (vízbontás) - reguláló sapka. - proximális antenna. - disztális antenna. 2. Funkciói. vízbontás: 2H2O O2 +4H+ + 4e- proton keletkezik a lumenben, elektron lép az e- transzportláncba. a plasztokinon redukciója.")
26
A PSII szerkezete

27
Az elektrontranszportlánc felépítése
- membránhoz kötött komponensek PS II, citokróm b6/f komplex, PS I, ferredoxin-NADP+ reduktáz - mobilis komponensek plasztokinon pool, PQH2 (membránban, PS II. és citokróm b6/f komplex között) plasztocianin, PC (lumen, citokróm b6/f komplex és PS I. közt) ferredoxin (PS I. és ferredoxin-NADP+ reduktáz között)
 plasztocianin, PC (lumen, citokróm b6/f komplex és PS I. közt) ferredoxin (PS I. és ferredoxin-NADP+ reduktáz között)")
29
A citokróm b6/f komplex működése
oxidálja a PQH2-t, redukálja a PC-t a ciklusos elektrontranszportban oxidálja a ferredoxint proton átadást közvetít a sztrómából a lumenbe

30
A PS I. felépítése és működése
Reakciócentrum: 2 db centrális fehérje P700 klorofilla elsődleges (Kla) és másodlagos (K-vitamin) akceptorok vas-kén centrumok PC és ferredoxin kötő fehérjék Nem ciklusos elektrontranszport: az e- a NADP+ felé, Ciklusos elektrontranszport: e- a citokróm b6/f komplex felé Ferredoxin, ferredoxin-NADP+ reduktáz, NADP+ Eredmény: NADPH + H+
 és másodlagos (K-vitamin) akceptorok. vas-kén centrumok. PC és ferredoxin kötő fehérjék. Nem ciklusos elektrontranszport: az e- a NADP+ felé, Ciklusos elektrontranszport: e- a citokróm b6/f komplex felé. Ferredoxin, ferredoxin-NADP+ reduktáz, NADP+ Eredmény: NADPH + H+")
31
14-49 A H+-koncentrációkülönbség és az ATP képződés
A két folyamat a mitokondriumban és a kloroplasztiszban hasonló módon játszódik le. Mindkét esetben az ATP szintézis egy protontranszlokáló enzim, az ATP szintáz, segítségével megy végbe. A proton áramlás a magasabb, lúgosabb pH felé tart Az enzim aktív helyei a „gomba fejében” helyezkednek el. Az ATP a mitokondriumi matrixban illetve a kloroplasztisz beli sztrómában keletkezik.

32
A fotoszintetikus foszforiláció: kemiozmotikus modell
proton elektrokémiai potenciálgrádiens a lumen és a sztróma között: ezt használja fel az ATP- szintáz enzim az ATP szintézisére ADP-ből és anorganikus foszfátból

33
14-49 Az elektontranszportlánc és a foszforiláció
A két folyamat között olyan szoros a kapcsolat hogy ATP-képződés nélkül az elektrontanszportlánc leáll, és fordítva. Szétkapcsoló anyagok megszüntetik ezt a szoros kapcsolatot. Ilyen szétkapcsoló anyagok a következők: CCCP, valinomicin, gramicidin D, DCPIP stb.

34
2. A fotoszintézis sötét szakasza: Calvin ciklus
Jelzett, H14CO3- adagolásával a széndioxid fixálás elsődleges termékei kimutathatók

35
A sötét szakasz legfontosabb biokémiai folyamatai
- karboxilációs fázis ribulóz-1,5-bifoszfát + CO2 = 2 glicerinsav-3-foszfát Katalizálja: a ribulóz-1,5-bifoszfát-karboxiláz, oxigenáz (RUBISCO) - redukció szakasza foszforiláció, az ATP 2/3 részének felhasználása, redukció, a redukált NADPH felhasználása a glicerinsav-3- foszfát redukciójára - regeneráció a ribulóz-1,5-bifoszfát-karboxiláz regenerálása: 6 glicerinsav-3-foszfát 3 ribulóz-1,5-bifoszfát+1 trióz- foszfát Nyereség ciklusonként egy triózfoszfát!
 - redukció szakasza. foszforiláció, az ATP 2/3 részének felhasználása, redukció, a redukált NADPH felhasználása a glicerinsav-3- foszfát redukciójára. - regeneráció. a ribulóz-1,5-bifoszfát-karboxiláz regenerálása: 6 glicerinsav-3-foszfát 3 ribulóz-1,5-bifoszfát+1 trióz- foszfát. Nyereség ciklusonként egy triózfoszfát!")
36
Kettős enzimaktivitású – karboxiláz és oxigenáz (3:1)
14-38 RUBISCO Ribulóz-1,5-biszfoszfát-karboxiláz-oxigenáz a reakciót katalizáló enzim Kettős enzimaktivitású – karboxiláz és oxigenáz (3:1) Kompetíció, ha mindkét szubsztrátum jelen van.
 Kompetíció, ha mindkét szubsztrátum jelen van.")
37
A RUBISCO SZERKEZETE

38
14-38 A sötétszakasz kezdete: karboxiláció
A Calvin ciklus folyamán a C3-as úton a széndioxid a ribulóz 1,5 biszfoszfáthoz kötődik és egy feltételezett hat szénatomos intermedier termék közbeejtésével két molekula glicerinsav-3-foszfáttá alakul át. A glicerinsav-3-foszfát a fotoszintetikus CO2-fixáció első stabil terméke

39
14-39 A Calvin ciklus jelentősége
Három pentózfoszfátból 3 molekula széndioxid megkötésével 6 triózfoszfát keletkezik A három pentózfoszfát regenerálódik A 3 széndioxid molekulából a nettó eredmény 1 exportálódó triózfoszfát Ez cukrok (szacharóz), zsírsavak, aminosavak szintézisére szolgál A folyamat során 9 ATP és 6 NADPH használódik fel
, zsírsavak, aminosavak szintézisére szolgál. A folyamat során 9 ATP és 6 NADPH használódik fel.")
40
Az UV-B sugárzás hatása a növényekre

41
A földi légkör oxigéntartalmának kialakulása és változása a földtörténet folyamán

42
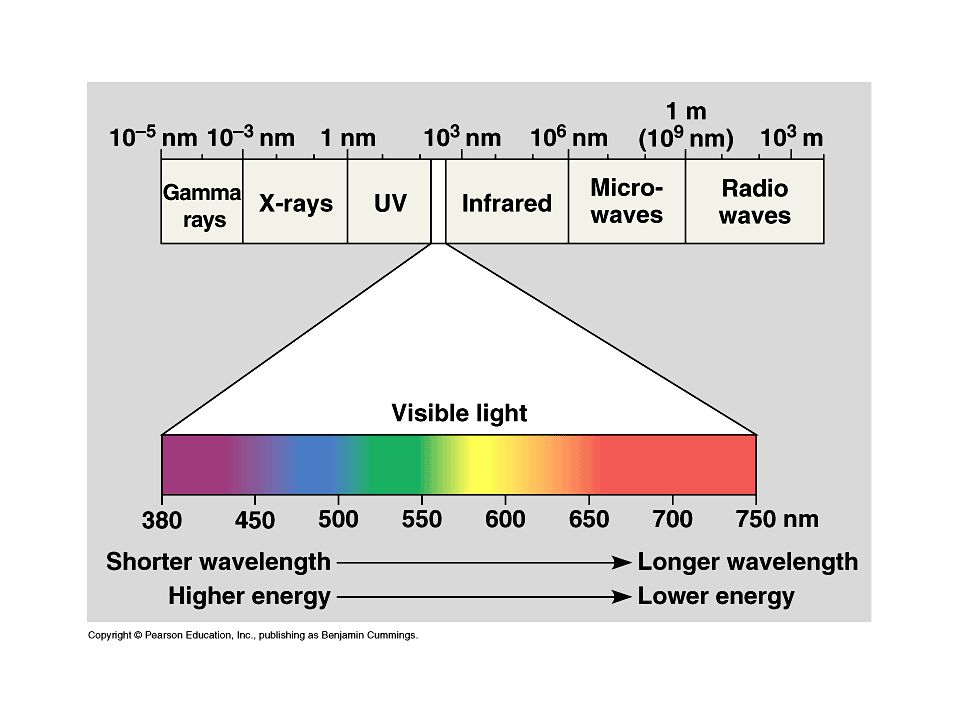
Az UV sugárzás a nm hullámhossz tartományba eső, a látható fénynél nagyobb energiájú elektromágneses sugárzás

43
UV-C sugárzás: 150-280 nm → a sztratoszférikus ózonréteg elnyeli
UV-B sugárzás: nm → a sztratoszférikus ózonréteg részben elnyeli UV-A sugárzás: nm → eléri a földfelszínt A sztratoszférikus ózonréteg kialakulása: oxigéntermelő, fotoszintetizáló szervezetek →O2 UV-C fotonok abszorpciója után O atomok → O atomok kombinálódása O2 molekulákkal → O3

44
Az UV-B sugárzás nagysága függ:
a Nap emissziójától a Nap-Föld távolságtól a Nap állásától a földrajzi szélességtől az ózonrétg vastagságától a felhőzettől a légkör aeroszol tartalmától a légnyomástól a tengerszint feletti magasságtól és a felszín UV-visszaverő képességétől

45
Björn és Murphy modellje (1985) szerint:
a normál UV-B sugárzás: 3,0 kJ m-2 nap-1 15 %-os ózoncsökkenésnél: 3,8 kJ m-2 nap-1 (átlag) 5000 m magasságban: 10,0 kJ m-2 nap-1 (46 oN, tiszta égbolt)
 5000 m magasságban: 10,0 kJ m-2 nap-1 (46 oN, tiszta égbolt)")
46
(a színek az ózonoszlop nagyságát Dobson egységekben jelölik)
A troposzférikus ózonmennyiség nagyságának éves változása és elhelyezkedése (a színek az ózonoszlop nagyságát Dobson egységekben jelölik)
")
47
Az Északi Sark fölött kialakuló ózonlyuk (sztartoszférikus) a múlt század utolsó évtizedeiben (a kék szín az ózonréteg elvékonyodását jelöli)
 a múlt század utolsó évtizedeiben (a kék szín az ózonréteg elvékonyodását jelöli)")
48
A sztratoszférában mért átlagos évi ózonhiány 1979-1993. között

49
Az UV-B sugárzást abszorbeáló molekulaféleségek növényekben

50
A szárazföldet meghódító növények ki voltak téve a felszínt elérő UV-B sugárzásnak.

51
A növényekben UV-B sugárzás hatására kialakuló morfológiai változások

52
Az UV-B sugárzás hatására akkumulálódó antociánok (vörös) és az epidermiszben felhalmozódó flavonoidok (sárga fluoreszcencia) kiszűrik a káros hatású sugarakat Kukorica csíranövények Fenyőtű keresztmetszeti képe

53
Az UV-B sugárzás hatása a növényekre
abszorbeálják: nukleinsavak, fehérjék, flavoproteinek morfológiai változások: biomassza csökken csökkent szármagasság, rövidebb internódiumok oldalelágazások száma nől csökkent levélfelület, sztómaszám, levélpödrődés vastag levél, vastag epidermisz UV-szűrő pigmentek akkumulációja az UV-B tolerancia egyensúlyt jelent a károsító folyamatok és a javító mechanizmusok között.

54
Az UV-B sugárzás legveszélyesbb hatása a DNS molekulában kialakuló timin dimerek képződése

55
A rezisztens növények hatékony javító mechanizmusokkal rendelkeznek: az ún. fotoliáz enzim aktiválódik és hasítja a ciklobután gyűrűt

56
Vannak olyan mutáns növények, amelyek magas UV-B terhelést is problémametesen túlélnek: ilyen a lúdfű uvi1 mutánsa Tanulság: az evolúció az emberi faj kipusztulása után is folytatódni fog

57
A természetes környezetet érő extra UV-B sugárzás hatásának mérésére speciális szűrőket használnak.
Ez egy mérőállomás Argentínában.

58
A fenolkarbonsavak, a lignin és a flavonoidok az aromás aminosav fenilalaninból szintetizálódnak.
Részben védelmet jelentenek a növény számára. A lignin az emészthetőséget csökkentő salakanyag, hosszútávú szén depozit.

59
A galluszsavból származó tanninok a növényevő állatok által elfogyasztott növényi táplálék élvezhetőségét befolyásolják. Fanyar íz, reagál a nyál fehérjéivel. (tannin: cseranyag) Szintézisük UV-B hatására fokozódik
 Szintézisük UV-B hatására fokozódik.")
60
AZ UV-B HATÁSA A SZÁRAZFÖLDI ÖKOSZISZTÉMÁRA
Közvetlen hatás: csökkent biomassza produkció Mikroorganizmusok közti interakció vízi élőhelyen: a zooplankton a növényi plankton karotinoid pigmentjeit használja védő faktorként A flavonoidok: virág színanyagok, növény-mikróba interakció közvetítők Tanninok: fehérjékkel reagálnak: a táplálék emészthetősége, élvezeti értéke csökken Lignin: a növényi hulladék mikroorganizmusok általi bonthatósága csökken (hosszú élettartamú széndepó) Az UV-B a növényi hulladék fotodegradációját közvetlenül serkenti (nyílt területeken: füves puszta, düne). A mikroorganizmusok (baktériumok, gombák) lebontó működését gátolja.
 Az UV-B a növényi hulladék fotodegradációját közvetlenül serkenti (nyílt területeken: füves puszta, düne). A mikroorganizmusok (baktériumok, gombák) lebontó működését gátolja.")
Hasonló előadás








 gáz, vagy gázelegy Lézerátmenet: elektronszintek között (UV és látható lézerek) rezgési szintek.>")










