Előadást letölteni
1
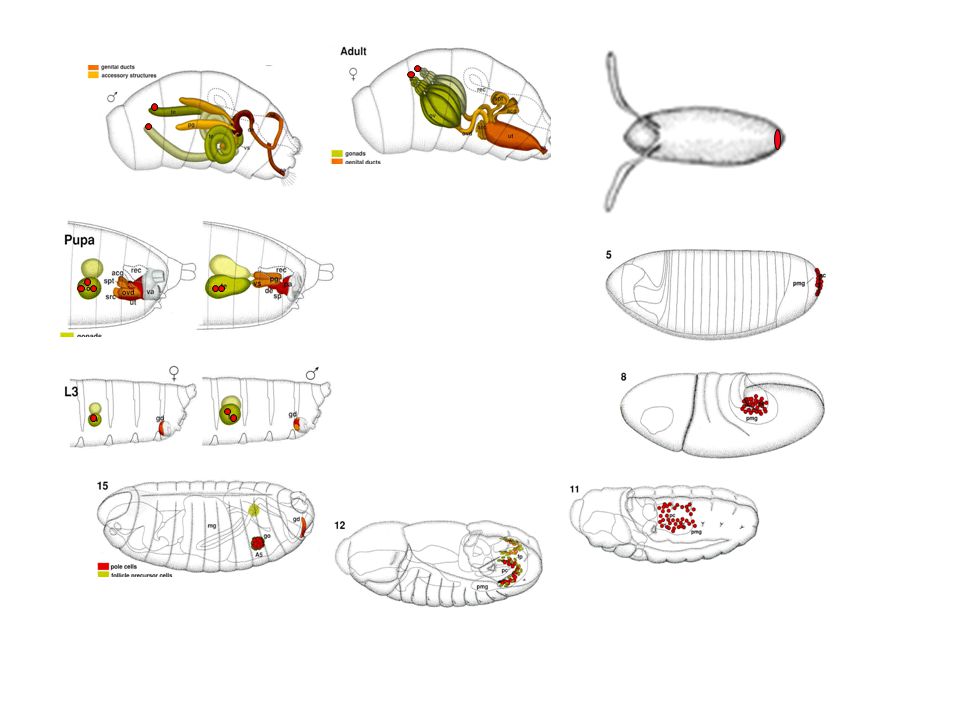
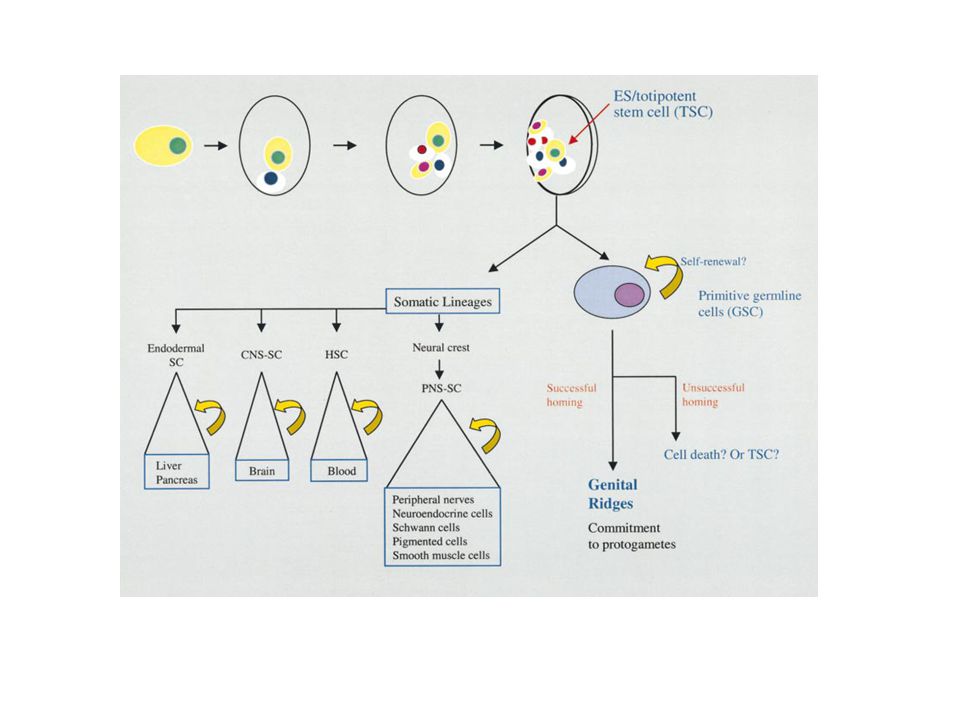
Ivarsejtfejlődés Drosophila melanogasterben

2
Az Escherichia coli biofilm életciklusa
Az Escherichia coli biofilm életciklusa. (1) A szabadon élő baktériumok hordozó felületre tapadnak, majd (2) tapadást elősegítő sejten kívüli állományt fejlesztenek. (3,4) Létrejön a biofilm amely érése során sajátságos alakot vesz fel, és belső testfelépítést alakít ki. (5) A biofilmet alkotó baktériumok osztódása során újra szabadon élő baktériumok jönnek létre, melyek alkalmas környezetben újabb biofilmet fejleszthetnek ki Konfliktus az egyes sejtek között
 A szabadon élő baktériumok hordozó felületre tapadnak, majd (2) tapadást elősegítő sejten kívüli állományt fejlesztenek. (3,4) Létrejön a biofilm amely érése során sajátságos alakot vesz fel, és belső testfelépítést alakít ki. (5) A biofilmet alkotó baktériumok osztódása során újra szabadon élő baktériumok jönnek létre, melyek alkalmas környezetben újabb biofilmet fejleszthetnek ki. Konfliktus az egyes sejtek között.")
3
A: Volvocidae ostoros fotoszintetizáló alga család tagjai átmenetet mutatnak az (A) egysejtű, (B,C) sejtdifferenciációt nem mutató többsejtű, valamint (D,E,F) a nagy belső szaporodó (gonídium) és a kicsi külső, steril testi sejtekből álló soksejtes életformák között. Chlamydomonas reinhardtii; B: Gonium pectorale; C: Eudorina elegans; D: Pleodorina californica; E: Volvox carteri; F: Volvox aureus. Nincs konfliktus az egyes sejtek között

4
Érvényes ez a séma a magasabb rendűekre is?

5
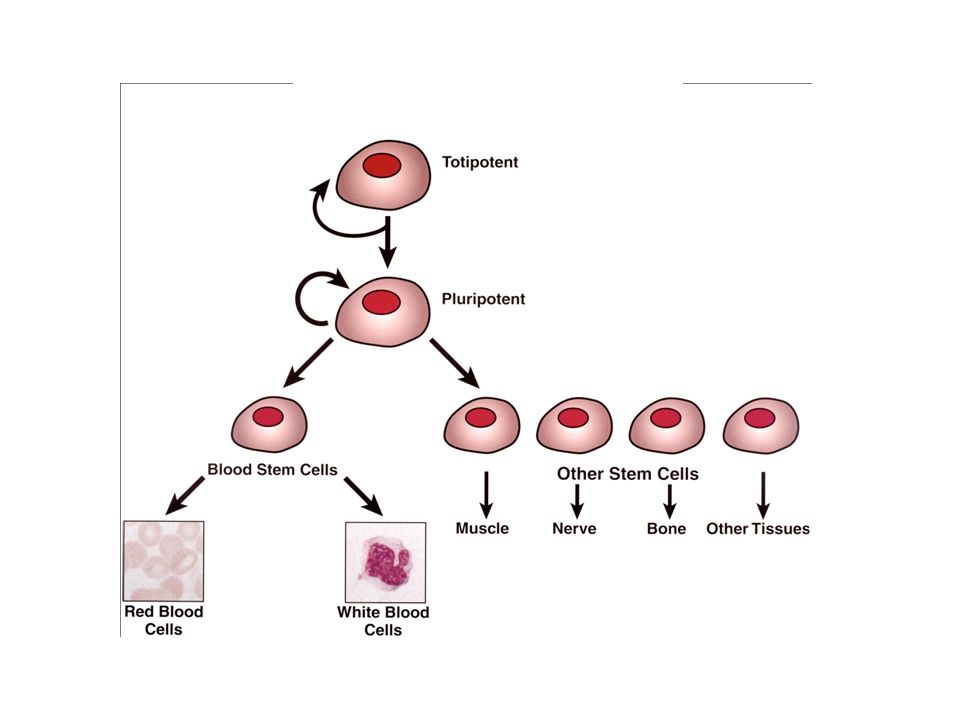
1978: emberi sztemsejtek vérből
1981: első egérből származó sztemsejt- tenyészetek 1988: Embrionális sztemsejt-tenyészet hörcsögből 1997: Dolly klónozás (totipotencia) 1997: rák sztemsejtek felefedezése
 1997: rák sztemsejtek felefedezése.")
7
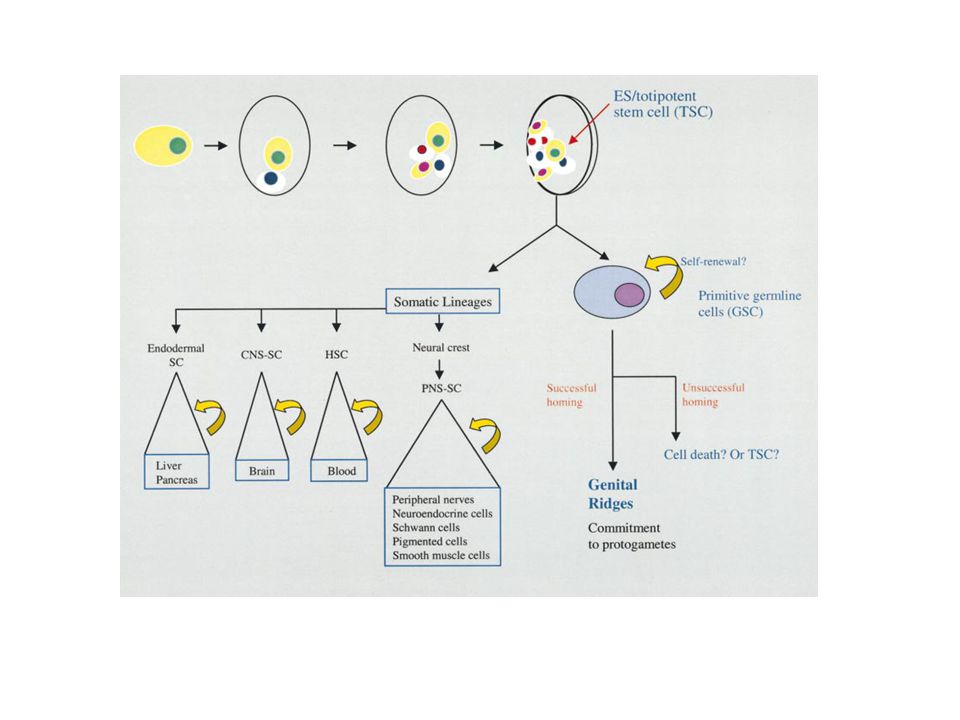
Niche

8
Niche Transit amplifying steps

9
Transit amplifying steps
Niche Primordial germ cells Transit amplifying steps

10
Stem Cells: Units of Development,
Units of Regeneration, and Units in Evolution Cell, Vol. 100, 157–168, January 7, 2000, Irving L. Weissman Stanford University School of Medicine Department of Pathology Stanford University Medical Center Stanford, California 94305–5323

15
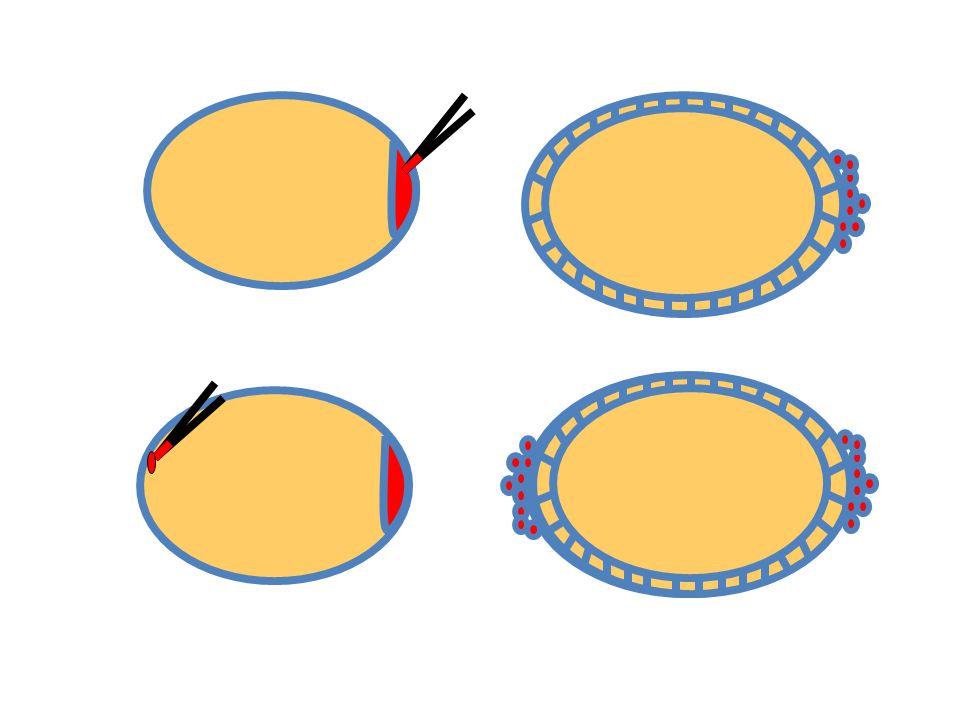
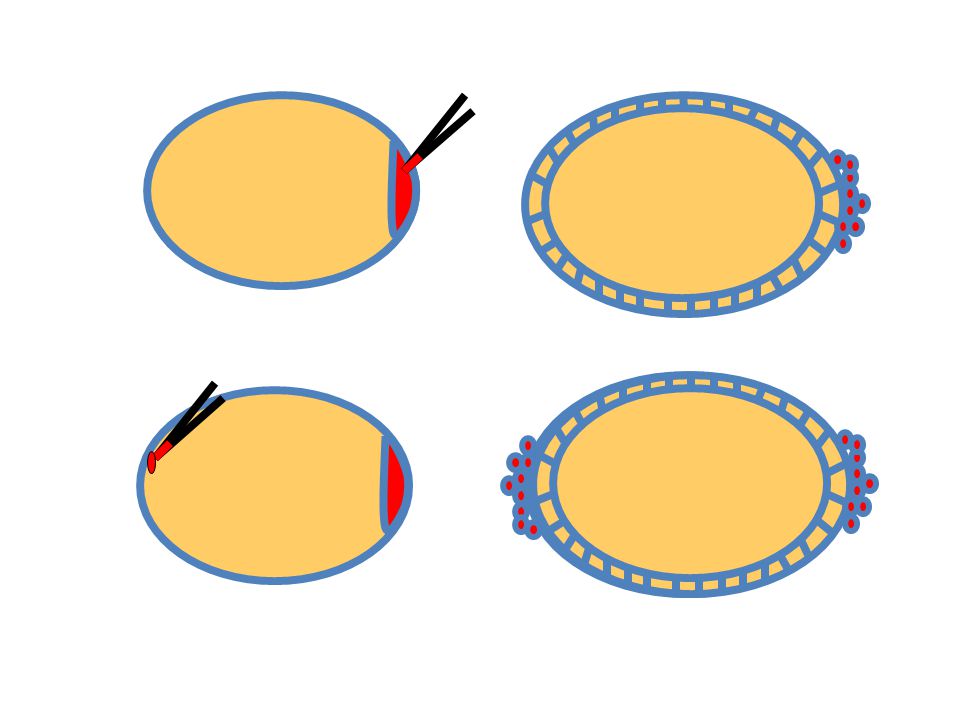
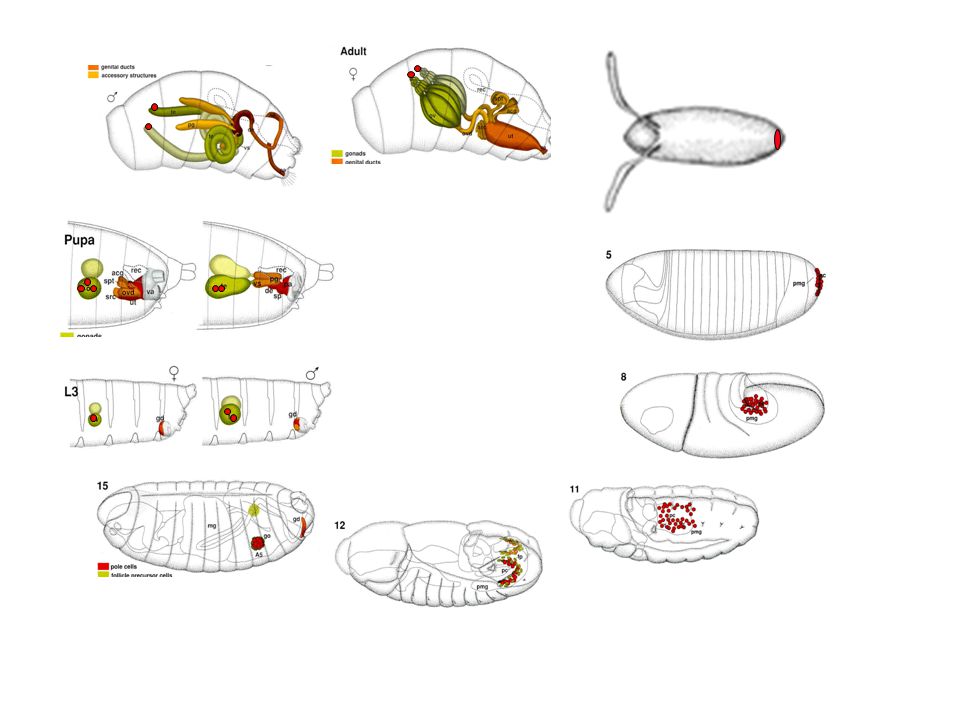
Drosophila ivarsejt sztemsejt és ivarplazma

17
Preformált ><Epigenetikus

19
Preformáció-Epigenezis, melyik az elsődleges?
Extavour szerint az epigenezis, az ivarsejtek később tanulják meg magukat ellátni ivarsejtfajtorokkal. Sztemsejt-ivarsejt hasonlóság. Cassandra Extavour szerint is az ivarsejtek egyfajta sztemsejtek. Van erre kísérleti bizonyíték?

20
PNAS November 25, vol no Kísérletes bizonyíték az ivarsejt-szemsejt ekvivalencia a Drosophila ivarsejtfejlődés során A poláris plazma vizsgálatával általános ivarsejt jellemzőket vizsgálunk

21
Az ivarsejtek egyfajta sztemsejtek, melyek generációról generációra átadódnak.
Az ivarsejtek differenciációja eltér a testi sejtek differenciációjától. Az ivarsejtek esetében inkább egy fejlődéstani alapállapot megőrzéséről van szó.

23
Oskar RNS, Staufen fehérje (Anne Ephrussi, Daniel st Johnston)
")
24
Anterior oskar RNS ektopikus ivarsejteket eredményez (Anne Ephrussi)
Lokalizált RNS és annak helyspecifikus transzlációja

25
Klasszikus genetikai megközelítés

26
Az anyai ivarsejtfaktorok hiányában nem fűződnek le embrionális ivarsejtek

27
ampR White ori P transzpozáz rosy LacZ A B Bier et al. Genes Dev (9): A kanamicin R white ori H transzpozáz rosy LacZ B Smith et al Genetics 135: 1063-

28
Az aktin kötő tropomiozin részt vesz az OSKAR fehérje lokalizációban
vt Anterior oskar Tropomiozin/Tropomiozin Tropomiozin/Tropomiozin, Anterior oskar

29
nincs polyA nincs promóter saját promóter saját polyA Stop-Start SAS
A klaszikus transzpozon rendszerek hatékonyságának növelése géncsapdázással EGFP hs-neo1 w+ nincs polyA SAS SDS nincs promóter saját promóter polyA saját polyA Stop-Start Jankovics Ferenc Lukácsovich Tamás

30
Az oskar fehérje az aktin vázhoz horgonyzódik ki
Vad típus Moesin Jankovics Ferienc

31
A klasszikus megközelítés korlátai pleiotrópia, redundancia
Genetics 153: (September 1999) An Exploration of the Sequence of a 2.9-Mb Region of the Genome of Drosophila melanogaster M. Ashburner, S. Misra, J. Roote, S. Lewis, R. Blazej, T. Davis, C. Doyle, R. Galle, R. George, N. Harris, G. Yartzell, D. Harvey, L. Hong, K. Houston, R. Hoskins, G. Johnson, C. Martin, A. Moshrefi, M. Palazzolo, M. Reese, A. Spradling, G. Tsang, K. Wan, K. Whitelaw, B. Kimmel, S. Celniker and G.M. Rubin. A contiguous sequence of nearly 3 Mb from the genome of Drosophila melanogaster has been sequenced from a series of overlapping P1 and BAC clones. This region covers 69 chromosome polytene bands on chromosome arm 2L, including the genetically well-characterized "Adh region." A computational analysis of the sequence predicts 218 protein-coding genes, 11 tRNAs, and 17 transposable element sequences. At least 38 of the protein-coding genes are arranged in clusters of from 2 to 6 closely related genes, suggesting extensive tandem duplication. The gene density is one protein-coding gene every 13 kb; the transposable element density is one element every 171 kb. Of 73 genes in this region identified by genetic analysis, 49 have been located on the sequence; P-element insertions have been mapped to 43 genes. Ninety-five (44%) of the known and predicted genes match a Drosophila EST, and 144 (66%) have clear similarities to proteins in other organisms. Genes known to have mutant phenotypes are more likely to be represented in cDNA libraries, and far more likely to have products similar to proteins of other organisms, than are genes with no known mutant phenotype. Over 650 chromosome aberration breakpoints map to this chromosome region, and their nonrandom distribution on the genetic map reflects variation in gene spacing on the DNA. This is the first large-scale analysis of the genome of D. melanogaster at the sequence level. In addition to the direct results obtained, this analysis has allowed us to develop and test methods that will be needed to interpret the complete sequence of the genome of this species.
 An Exploration of the Sequence of a 2.9-Mb Region of the Genome of Drosophila melanogaster. M. Ashburner, S. Misra, J. Roote, S. Lewis, R. Blazej, T. Davis, C. Doyle, R. Galle, R. George, N. Harris, G. Yartzell, D. Harvey, L. Hong, K. Houston, R. Hoskins, G. Johnson, C. Martin, A. Moshrefi, M. Palazzolo, M. Reese, A. Spradling, G. Tsang, K. Wan, K. Whitelaw, B. Kimmel, S. Celniker and G.M. Rubin. A contiguous sequence of nearly 3 Mb from the genome of Drosophila melanogaster has been sequenced from a series of overlapping P1 and BAC clones. This region covers 69 chromosome polytene bands on chromosome arm 2L, including the genetically well-characterized Adh region. A computational analysis of the sequence predicts 218 protein-coding genes, 11 tRNAs, and 17 transposable element sequences. At least 38 of the protein-coding genes are arranged in clusters of from 2 to 6 closely related genes, suggesting extensive tandem duplication. The gene density is one protein-coding gene every 13 kb; the transposable element density is one element every 171 kb. Of 73 genes in this region identified by genetic analysis, 49 have been located on the sequence; P-element insertions have been mapped to 43 genes. Ninety-five (44%) of the known and predicted genes match a Drosophila EST, and 144 (66%) have clear similarities to proteins in other organisms. Genes known to have mutant phenotypes are more likely to be represented in cDNA libraries, and far more likely to have products similar to proteins of other organisms, than are genes with no known mutant phenotype. Over 650 chromosome aberration breakpoints map to this chromosome region, and their nonrandom distribution on the genetic map reflects variation in gene spacing on the DNA. This is the first large-scale analysis of the genome of D. melanogaster at the sequence level. In addition to the direct results obtained, this analysis has allowed us to develop and test methods that will be needed to interpret the complete sequence of the genome of this species.")
32
Genetikai Interakción alapuló megközelítés
osk mRNS konc. TmIIeg TmIIel4 osk54 E(To) TmIIeg TmIIel4 osk54 Jankovics Ferenc
 TmIIeg9 + + TmIIel4 osk54. Jankovics Ferenc.")
33
A Rab11 mutációja az oskar RNS lokalizációs hibáját okozza
wt Rab11 Rab11 Rab11 Jankovics Ferenc

34
The role of membranes and membrane trafficking in RNA localization
Robert S. Cohen Biol. Cell (2005) 97, 5–18 A membránfüggő RNS transzport általános modellje
 97, 5–18. A membránfüggő RNS transzport általános modellje.")
35
; + + ; + osk mRNS konc. stau osk TmII stau osk TmII mutáció
Hoffman Gyula, Szuperák Milán

36
Ivarsejthiányos fenotípus
penetranciája Megvizsgált mutáns vonalak (600 db Maroy Péter) Szuperák Milán
 Szuperák Milán.")
37
Agoston V, Csermely P, Pongor S.
Multiple weak hits confuse complex systems: a transcriptional regulatory network as an example. Agoston V, Csermely P, Pongor S. Phys Rev E Stat Nonlin Soft Matter Phys May;71(5 Pt 1): Epub 2005 May 26. Hálózati szemlélet
: Epub 2005 May 26. Hálózati szemlélet.")
38
A fordított genetikai megközelítés

39
Az ivarsejtekben kifejeződő gének azonosítása
CG9925 Fly-FISH BDGP egyéb 502 ivarvonalban kifejeződő gén CG9924 CG15443 gcl CG8778

40
2 1 Ago3/ Aubergine/ Ago2 Ago1 Eszköz: a teljes genom szekvenálás utáni időszak jellegzetes reverz genetikai eljárása az RNSi Nature 451, (2008)
")
41
Injektálás optimalizálása saját készítésű kettős szálú RNS-ekkel
(lehet ivarsejtben csendesíteni!) Alkalmas ivarsejt markergén kiválasztása (nosMoe-GFP), mozi optimalizálása Ivarplazma és embrionális ivarsejt RNS-ek keresése adatbázisokból, egyéb irodalomból 500 db ivarplazma RNS elleni dsRNS Elsődleges, második screen Jankovics Ferenc, Köllőd Borbála, Henn László, Bujna Ágnes, Kiss Nóra, Velkeyné Krausz Ildikó, Ugrainé Szatmári Margit, Vilmos Péter, Michael Boutros,
 Alkalmas ivarsejt markergén kiválasztása (nosMoe-GFP), mozi optimalizálása. Ivarplazma és embrionális ivarsejt RNS-ek keresése adatbázisokból, egyéb irodalomból. 500 db ivarplazma RNS elleni dsRNS. Elsődleges, második screen. Jankovics Ferenc, Köllőd Borbála, Henn László, Bujna Ágnes, Kiss Nóra, Velkeyné. Krausz Ildikó, Ugrainé Szatmári Margit, Vilmos Péter, Michael Boutros,")
44
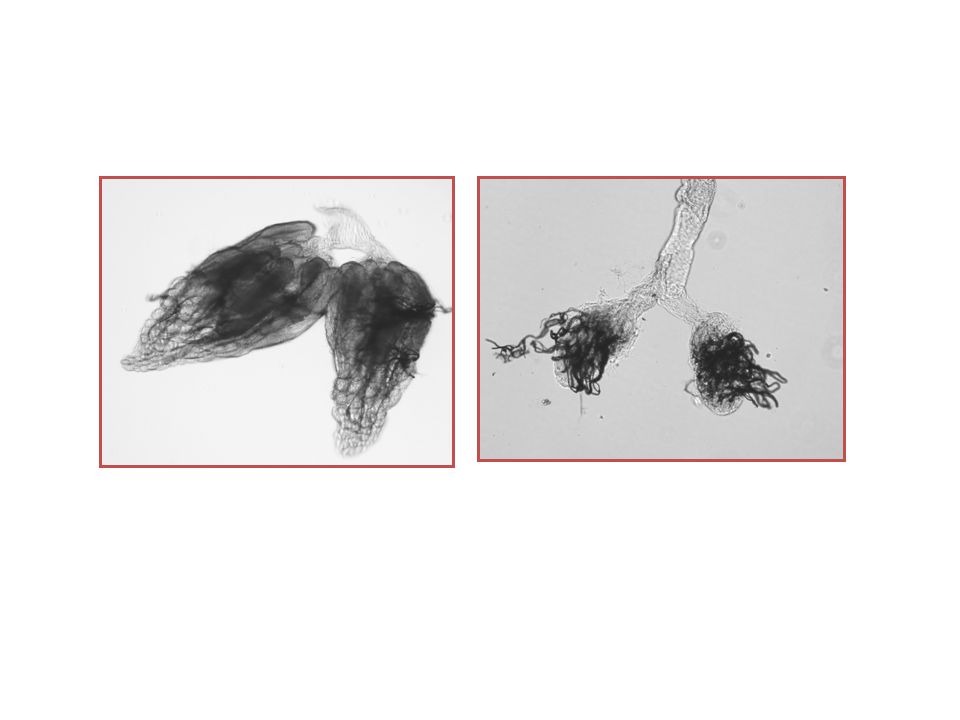
Embrionális ivarsejt-fenotípusok

45
A fascetto fenotípusa lárva korban
Embrió fascetto RNAi Petefészek Here Vasa Fasciclin III Traffic jam vad fascettoE07

46
A soksejtű életforma kialakulása a sztemsejtek kialakulásának köszönhető
Az ivarsejtek valójában speciális sztemsejtek Az epigenetikus ivarsejtfejlődés elsődleges a preformált ivarsejtfejlődés az előbbinek egy speciális esete A Drosophila ivarplazmája kiváló modell az ivarsejtfaktorok azonosításához A Drosophila ivarplazma összeszerveződésének kulcseleme az oskar RNS lokalizációja és helyspecifikus transzlációja A genetikai analízis három módszere klasszikus és fordított genetika, genetikai interakciós rendszerek Hálózatokban való gondolkodás szükségessége