Előadást letölteni
1
A fotoszintézis élettani és ökofiziológiai vonatkozásai
Fotoszintézis IV. A fotoszintézis élettani és ökofiziológiai vonatkozásai

2
A fotoszintézis fiziológiai és ökológiai vonatkozásai
Limitáló környezeti tényezők: fény, CO2, hőmérséklet -legfontosabb metabolikus lépések: Rubisco aktivitás a RuBP regenerációja trióz-P metabolizmus

3
A fény Energia fluxus (W, J) helyett foton fluxus sűrűség (molm-2s-1)
PAR: fotoszintetikusan aktív sugárzás ( nm, 2000 μmol m-2s-1 teljes napfényben) Abszorbeált, transzmittált és reflektált fény (10-15% veszteség)
 Abszorbeált, transzmittált és reflektált fény. (10-15% veszteség)")
4
Az összes beeső fényenergia %-os hasznosulása…

5
A levél által elnyelt, átengedett és reflektált fény %-os arányai

6
A távoli vörös tartomány jelentősége…

7
Az árnyék elkerülése „fény növények” szármegnyúlás „árnyék növények”
0,2 0,4 0,6 Pfr/Pössz

8
A szomszéd érzékelése

9
A levél hőmérsékletének szabályozása A Bowen arányszám
Közvetlen (vezetéssel és áramlással) hőleadás evaporációs hőleadás A Bowen arányszám alacsony, ha van transzspiráció és a levél és a levegő hőmérséklete megegyezik (pl. öntözött pázsit); - negatív, ha a transzsiráció következtében a levél hőmérséklete kisebb, mint a levegőé (pl. gyapot levél); - nagy (végtelen), ha zártak a sztómák (pl. sivatagi növények, kaktuszok)
 hőleadás. evaporációs hőleadás. A Bowen arányszám. alacsony, ha van transzspiráció és a levél és a levegő hőmérséklete megegyezik (pl. öntözött pázsit); - negatív, ha a transzsiráció következtében a levél hőmérséklete kisebb, mint a levegőé (pl. gyapot levél); - nagy (végtelen), ha zártak a sztómák (pl. sivatagi növények, kaktuszok)")
10
A levél anatómiája fényen és árnyékban
Rosta-hatás, fényvezetés, fényszórás Kloroplasztisz rendeződés és levélmozgás Nap-követő mozgás, dia- és paraheliotropizmus A fény spektrális megoszlása árnyékban: a fitokróm szerepe kompetícíó és adaptáció

11
A levél anatómiája a fényintenzitástól függően
Fényen nőtt levél Árnyékban nőtt levél

12
A fénylevelek nagyméretű oszlopos alapszövettel és több sejtsoros mezofillummal rendelkeznek: így optimálisan hasznosítják a fényt. Az árnyéklevelekben a fénybegyűjtő komplex aránya nagyobb a reakciócentrumhoz viszonyítva, mint a fénylevelekben.

13
A fotoszintetikus apparátus komponenseinek aránya erős és gyenge fényben

14
A jobb fénykihasználást szolgálja a kloroplasztiszok mozgása is
A „szűrőhatás” Felülnézetből: (a) árnyékban; (b) átlagos fényintenzitáson; (c) fénystressznél
 árnyékban; (b) átlagos fényintenzitáson; (c) fénystressznél.")
15
Az epidermisz réteg sejtjei mint fénykollektorok

17
Sok fajnál a levéllemez fénykövető mozgást mutat: heliotropizmus
Eredeti pozíció 4 óra ferde irányból való megvilágítás után Diaheliotropizmus: fénykövető mozgás (maximális besugárzás) Paraheliotropizmus: fénytől való elfordulás (minimális besugárzás)
 Paraheliotropizmus: fénytől való elfordulás (minimális besugárzás)")
18
A fotoszintézis intenzitásának napszaki változása burgonyánál

19
Napszaki változások az Opuntia ficus-indica esetében

20
A fényintenzitás szerepe
Fényintenzitás és [CO2], O2 Hőmérséklet, a kvantumhasznosítás hőmérséklet függése

21
A CO2 fixáció a fényintenzitás függvényében
A CO2 fixáció a fényintenzitás függvényében: a dózisgörbe kompenzációs pont ("sötét légzés") lineáris szakasz (kvantumhatásfok!) telítődés (metabolikus hatásfok)
 lineáris szakasz (kvantumhatásfok!) telítődés (metabolikus hatásfok)")
22
A fotoszintézis intenzitása a fényintenzitással növekszik: a kompenzációs pontnál a légzés CO2 termelése egyenlő a fotoszintézis CO2 fogyasztásával

23
A fénynövények fotoszintetikus hatékonysága magasabb telítődési értéket mutat és kompenzációs pontjuk is magasabban van, mint az árnyéknövényeké

24
Egy árnyéknövény fotoszintézisének intenzitása

25
Fotoszintézis és komponensei ökológiai méretekben

26
Nagy fényintenzitás (fénystressz) esetén reaktív oxigén formák keletkeznek, ami degradálja a fotoszintetikus apparátust: fénygátlás
 esetén reaktív oxigén formák keletkeznek, ami degradálja a fotoszintetikus apparátust: fénygátlás")
27
A fotoszintézis fénygátlása (fotoinhibíció)
")
28
Akceptor oldali fotoinhibíció
Donor oldali fotoinhibíció P680+/Feo- TyrZ+/P680+

29
A xantofill (VAZ) ciklus
Szerepe: Energia disszipáció, nem-fotokémiai kioltás

30
A xantofill (VAZ) ciklus
Sztróma Lumen pH optimum: 7,5 pH optimum: 5,2

31
A széndioxid Biokémiai reakciói a parciális nyomástól (mólfrakció x össznyomás) függenek - az üvegház hatás -kutikula (átjárhatatlan) -gáz fázis: sztómanyílás sztóma alatti üreg sejtközötti légterek - folyadék fázis: sejtfal, citoszól, kloroplasztisz limitáló tényező: a sztómaellenállás (regulálható!)
 -gáz fázis: sztómanyílás. sztóma alatti üreg. sejtközötti légterek. - folyadék fázis: sejtfal, citoszól, kloroplasztisz. limitáló tényező: a sztómaellenállás (regulálható!)")
32
CO2 asszimiláció az intercelluláris CO2 konc. (ci) függvényében
- ci számolható a CO2 asszimiláció (A) és sztómaellenállás (r), ill. vezetőképesség (g = 1/r) ismeretében: ci = ca - (1.6 AP/g), ahol ca = CO2 parciális nyomása a levegőben, P = atmoszférikus nyomás. C3-as és C4es növények, CO2-kompenzációs pont Vízhasznosítási hatékonyság (WUE) WUE = A/E = (ca - ci)/1.6(ei - ea) E = evaporáció, e = a víz parciális nyomásai. 1/WUE = transpirációs hányados
 és sztómaellenállás (r), ill. vezetőképesség (g = 1/r) ismeretében: ci = ca - (1.6 AP/g), ahol ca = CO2 parciális nyomása a levegőben, P = atmoszférikus nyomás. C3-as és C4es növények, CO2-kompenzációs pont. Vízhasznosítási hatékonyság (WUE) WUE = A/E = (ca - ci)/1.6(ei - ea) E = evaporáció, e = a víz parciális nyomásai. 1/WUE = transpirációs hányados.")
33
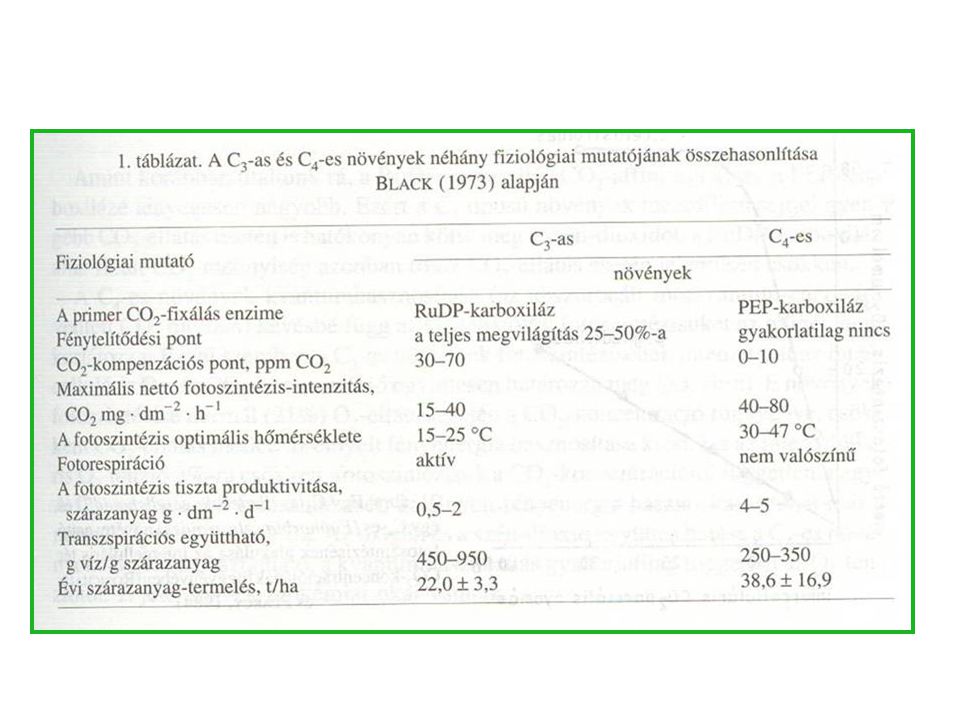
C3 és C4 levélszerkezet A C4-es növények anatómiája szembeszökően különbözik a C3-as növényekétől. Csak a C4-es növényekben találhatók a jellegzetes morfológiájú hüvelyparenchima (bundle sheath) vagy Kranz sejtek. A széndioxid elsődleges kötését a foszfoenol-piroszőlősav (PEP) karboxiláz enzim katalizálja a mezofill sejtek citoplazmájában.
 vagy Kranz sejtek. A széndioxid elsődleges kötését a foszfoenol-piroszőlősav (PEP) karboxiláz enzim katalizálja a mezofill sejtek citoplazmájában.")
35
C3-as és C4-es növények fotoszintézise a fényintenzitás (A) és az intercelluláris tér CO2 koncentrációjának (B) függvényében A B C-4 C-4 C-3 C-3 Fényintenzitás

36
C3-as és C4-es növények fotoszintézise a külső és az intercelluláris tér CO2 koncentrációjának függvényében

37
A C4-es növények fotoszintetikus hatékonyságának hőmérsékleti optimuma magasabb értéknél található, mint a C3-as növényeké

38
A kvamtumhasznosítási tényező

40
12C és 13C megkülönböztetés
Atmoszférikus CO2 : 12C – 98,9% 13C – 1,1% 14C – 10-10% -izotóp összetétel, δ13C‰ = (Rminta/Rstandard –1) x 1000 -izotóp diszkrimináció, Δ13C‰ = (Rlevegő/Rnövény –1) x 1000 -C3-as növények δ13C = -28‰ rubisco: δ13C = -30‰ -C4-es δ13C = -14‰ PEP karboxiláz: δ13C = ‰ CAM növények: δ13C = a két érték között (vízellátottságtól függően) -a C4-es út 7 millió évvel ezelőtt fejlődött ki.
 x izotóp diszkrimináció, Δ13C‰ = (Rlevegő/Rnövény –1) x C3-as növények δ13C = -28‰ rubisco: δ13C = -30‰ -C4-es δ13C = -14‰ PEP karboxiláz: δ13C = ‰ CAM növények: δ13C = a két érték között (vízellátottságtól függően) -a C4-es út 7 millió évvel ezelőtt fejlődött ki.")
41
12C és 13C megkülönböztetés

42
Köszönöm a figyelmet!