Előadást letölteni
Az előadás letöltése folymat van. Kérjük, várjon

1
A CO2 asszimilációja: fixáció és redukció
Fotoszintézis III A CO2 asszimilációja: fixáció és redukció

2
1. A C-3-as fotoszintetikus szénredukció (Calvin ciklus)
Jelzett, H14CO3- adagolásával a széndioxid fixálás elsődleges termékei kimutathatók

3
A Calvin ciklus 3 fő szakasza

4
A sötétszakasz kezdete: karboxiláció
A Calvin ciklus folyamán a C3-as úton a széndioxid a ribulóz 1,5 biszfoszfáthoz kötődik és egy feltételezett hat szénatomos intermedier terméken keresztül két molekula glicerinsav-3-foszfáttá alakul át. A glicerinsav-3-foszfát a fotoszintetikus CO2-fixáció első stabil terméke

5
A sötét szakasz legfontosabb biokémiai folyamatai
- karboxilációs fázis ribulóz-1,5-bifoszfát + CO2 = 2 glicerinsav-3-foszfát Katalizálja: a ribulóz-1,5-bifoszfát-karboxiláz, oxigenáz (RUBISCO) - redukció szakasza foszforiláció, az ATP 2/3 részének felhasználása, redukció, a redukált NADPH felhasználása a glicerinsav-3- foszfát redukciójára - regeneráció a ribulóz-1,5-biszfoszfát-karboxiláz regenerálása: 6 glicerinsav-3-foszfát 3 ribulóz-1,5-biszfoszfát+1 trióz- foszfát Nyereség ciklusonként egy triózfoszfát!
 - redukció szakasza. foszforiláció, az ATP 2/3 részének felhasználása, redukció, a redukált NADPH felhasználása a glicerinsav-3- foszfát redukciójára. - regeneráció. a ribulóz-1,5-biszfoszfát-karboxiláz regenerálása: 6 glicerinsav-3-foszfát 3 ribulóz-1,5-biszfoszfát+1 trióz- foszfát. Nyereség ciklusonként egy triózfoszfát!")
6
RUBISCO Ribulóz-1,5-biszfoszfát-karboxiláz-oxigenáz a reakciót katalizáló enzim Kettős enzimaktivitású – karboxiláz és oxigenáz (3:1) Kompetíció, ha mindkét szubsztrátum jelen van.

7
A RUBISCO SZERKEZETE L8S8 alegységek (56 kDa, kloroplasztisz; 14 kDa, sejtmag)
")
8
Rubisco: az oldható fehérjék 40%-a.
N-raktár, a Föld népességének minden tagjára 20 kg jut… Évente 200 milliárd tonna CO2-t fixál

9
A Calvin ciklus, részletesebben

10
1. A C-3-as fotoszintetikus szénredukció (Calvin ciklus)
i) Karboxiláció ribulóz 1,5-biszfoszfát karboxiláz/oxigenáz (Rubisco) L8S8 alegységek (56 kDa, kloroplaszt; 14 kDa, sejtmag) Karboxiláz: ΔG, = kcal mol-1 Rubisco affinitása a CO2-hoz nagy Ru 1,5-biszfoszfát + CO2 intermedier 2,3-P-glicerát ii) Redukció - foszforiláció (+ATP) - tényleges redukció (NADPH +H+) gliceraldehid-3-P iii) Regeneráció 6 mól G-3-P 3 mól Ru-1,5-diP + 1 C-3 a nyereség
 Karboxiláció ribulóz 1,5-biszfoszfát karboxiláz/oxigenáz (Rubisco) L8S8 alegységek (56 kDa, kloroplaszt; 14 kDa, sejtmag) Karboxiláz: ΔG, = kcal mol-1 Rubisco affinitása a CO2-hoz nagy Ru 1,5-biszfoszfát + CO2 intermedier 2,3-P-glicerát ii) Redukció - foszforiláció (+ATP) - tényleges redukció (NADPH +H+) gliceraldehid-3-P iii) Regeneráció 6 mól G-3-P 3 mól Ru-1,5-diP + 1 C-3 a nyereség.")
11
iv) Sztoichiometriák. a ciklusra:
iv) Sztoichiometriák a ciklusra: 5 RuBP + 5 CO2 + 9 H2O + 16 ATP + 10 NADPH 6 RuBP + 14 HOPO H ADP + 10 NADP+ autokatalízis! nettó: CO H2O + 12 NADPH + 18 ATP 1 Fruktóz-6-P + 12 NADP+ + 6 H+ +18 ADP + 17 Pi azaz 2 mol NADPH + 3 mol ATP / 1 mol CO2 v) Termodinamikai hatékonyság Fény + sötét reakció : 33% Csak a sötét reakció: 90 % vi) A ciklus szabályozása - fényfüggő ionmozgások (H+/Mg2+ csere) - a Rubisco aktivációja karbamát képzéssel - thiol-alapú oxidációs-redukciós rendszer - a trióz-P kompartmentalizációja (kloroplasztisz/citoplazma)
 Sztoichiometriák a ciklusra: 5 RuBP + 5 CO2 + 9 H2O + 16 ATP + 10 NADPH 6 RuBP + 14 HOPO H ADP + 10 NADP+ autokatalízis! nettó: 6 CO H2O + 12 NADPH + 18 ATP 1 Fruktóz-6-P + 12 NADP+ + 6 H+ +18 ADP + 17 Pi azaz 2 mol NADPH + 3 mol ATP / 1 mol CO2. v) Termodinamikai hatékonyság Fény + sötét reakció : 33% Csak a sötét reakció: 90 % vi) A ciklus szabályozása - fényfüggő ionmozgások (H+/Mg2+ csere) - a Rubisco aktivációja karbamát képzéssel - thiol-alapú oxidációs-redukciós rendszer - a trióz-P kompartmentalizációja (kloroplasztisz/citoplazma)")
12
A Calvin ciklus jelentősége
Három pentózfoszfátból 3 molekula széndioxid megkötésével 6 triózfoszfát keletkezik A három pentózfoszfát regenerálódik A 3 széndioxid molekulából a nettó eredmény 1 exportálódó triózfoszfát Ez cukrok (szacharóz), zsírsavak, aminosavak szintézisére szolgál A folyamat során 9 ATP és 6 NADPH használódik fel
, zsírsavak, aminosavak szintézisére szolgál. A folyamat során 9 ATP és 6 NADPH használódik fel.")
13
A Calvin ciklus számos enzimét a fényfüggő ferredoxin-tioredoxin rendszer aktiválja
NADP: gliceraldehid-3-foszfát dehidrogenáz Fruktóz-1,6-biszfoszfát foszfatáz Szedoheptulóz-1,7-biszfoszfát foszfatáz Ribulóz-5-foszfát kináz

14
A C-2-es fotorespirációs (karbon oxidációs, glikolát) ciklus
A Rubisco kettős aktivitásának következménye…

15
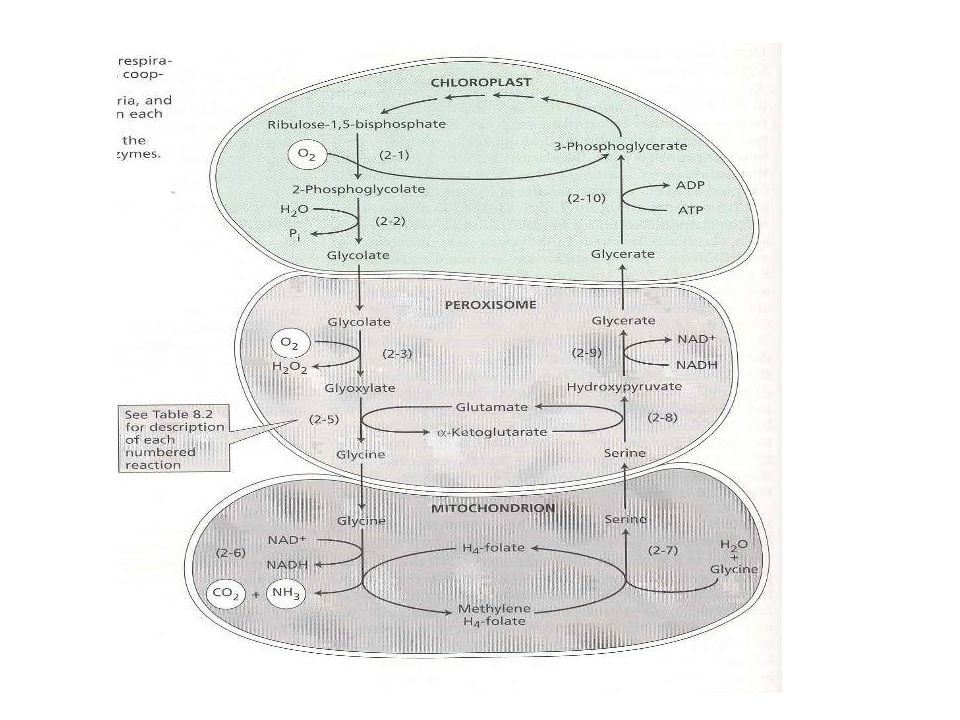
2. A C-2-es fotorespirációs (karbon oxidációs, glikolát) ciklus
- a Rubisco oxigenáz aktivitása következtében ("parazita") - CO2/O2 koncentrációk, hőmérséklet függő - a kloroplasztisz, peroxiszóma és a mitokondrium közreműködésével A.) A kloroplasztiszban: Ru-1,5-BP + O2 2-P-glikolát + 3-P glicerát P-glikolát + H2O glikolát
 - CO2/O2 koncentrációk, hőmérséklet függő. - a kloroplasztisz, peroxiszóma és a mitokondrium közreműködésével. A.) A kloroplasztiszban: Ru-1,5-BP + O2 2-P-glikolát + 3-P glicerát. P-glikolát + H2O glikolát.")
16
B.) A peroxiszómában (1):
glikolát + O2 glioxilát = H2O2 kataláz reakció glioxalát + glutamát glicin + α-ketoglutarát C.) A mitokondriumban: glicin + NAD+ + H4-folát NADH + H+ + CO2 + NH3 + metilén H4-folát metilén H4-folát + H2O + glicin szerin + H4-folát D.) A peroxiszómában (2): szerin + α-ketoglutarát hidroxipiruvát + glutamát hidroxipiruvát + NADH + H+ glicerát + NAD+ E.) A kloroplasztiszban: glicerát + ATP 3-P-glicerát + ADP + H+
 A mitokondriumban: glicin + NAD+ + H4-folát NADH + H+ + CO2 + NH3 + metilén H4-folát. metilén H4-folát + H2O + glicin szerin + H4-folát. D.) A peroxiszómában (2): szerin + α-ketoglutarát hidroxipiruvát + glutamát. hidroxipiruvát + NADH + H+ glicerát + NAD+ E.) A kloroplasztiszban: glicerát + ATP 3-P-glicerát + ADP + H+")
18
C3 és C4 levélszerkezet A C4-es növények anatómiája szembeszökően különbözik a C3-as növényekétől. Csak a C4-es növényekben található a hüvelyparenchima (bundle sheath) vagy Kranz sejtek. A széndioxid elsődleges kötését a PEP karboxiláz enzim katalizálja a mezofill sejtek citoplazmájában.
 vagy Kranz sejtek. A széndioxid elsődleges kötését a PEP karboxiláz enzim katalizálja a mezofill sejtek citoplazmájában.")
19
A kukoricalevél tipikus Kranz-anatómiája

20
3. A CO2 fixáció C-4-es útja (CO2 koncentráló mechanizmus)
- primér fixációs termék C-4 sav (oxálecetsav) - foszfoenol-piruvát (PEP) karboxilálódik (HCO3-) - a C-4-es és a C-3 ciklus térben elválasztódik: speciális anatómia (mezofill és hüvelyparenchima) - trópusi, szubtrópusi növényekben
 - foszfoenol-piruvát (PEP) karboxilálódik (HCO3-) - a C-4-es és a C-3 ciklus térben elválasztódik: speciális anatómia (mezofill és hüvelyparenchima) - trópusi, szubtrópusi növényekben.")
21
Az alapciklus lépései:
i) PEP karboxiláció a MEZOFILL sejtek citoplazmájában oxálecetsav malát, aszpartát ii) A C-4 sav transzportja a HÜVELYPARENCHIMA-ba iii) A C-4 sav (almasav, aszparaginsav) dekarboxilációja a CO2 fixációja a C-3-as ciklusban iv) A C-3 sav (piruvát, alanin) transzportja MEZOFILL
 PEP karboxiláció a MEZOFILL sejtek citoplazmájában. oxálecetsav malát, aszpartát. ii) A C-4 sav transzportja a HÜVELYPARENCHIMA-ba. iii) A C-4 sav (almasav, aszparaginsav) dekarboxilációja. a CO2 fixációja a C-3-as ciklusban. iv) A C-3 sav (piruvát, alanin) transzportja MEZOFILL.")
23
A dekarboxilációs mechanizmus alapján 3 típus:
i) NADP-almasav enzim (NADP-ME, kloroplasztisz) ii) NAD-almasav enzim (NAD-ME, mitokondrium) iii) Foszfoenol-piruvát -karboxikináz (PEP-CK, citoplazma) A CO2 koncentrálódása 8-10-szeres a C-3-as levélhez képest Energiamérleg: + 2 ATP/ CO2; nyereség: nincs fotorespiráció Fény-regulált kulcsenzimek: PEP karboxiláz NADP-almasav dehidrogenáz (tioredoxin) Piruvát-ortofoszfát-dikináz
 NADP-almasav enzim (NADP-ME, kloroplasztisz) ii) NAD-almasav enzim (NAD-ME, mitokondrium) iii) Foszfoenol-piruvát -karboxikináz (PEP-CK, citoplazma) A CO2 koncentrálódása 8-10-szeres a C-3-as levélhez képest. Energiamérleg: + 2 ATP/ CO2; nyereség: nincs fotorespiráció. Fény-regulált kulcsenzimek: PEP karboxiláz. NADP-almasav dehidrogenáz (tioredoxin) Piruvát-ortofoszfát-dikináz.")
24
A dekarboxilációs mechanizmus alapján 3 típus:

25
5. A szaharóz és a keményítő szintézise

26
5. A szaharóz és a keményítő szintézise
- keményítő: a kloroplasztiszban - szaharóz: a citoplazmában közös lépések a trióz-P-tól a glukóz-1-P-ig izoenzimek: pl.; Fruktóz-1,6-biszfoszfát foszfatáz kloroplasztban: tioredoxin citoszólban: fruktóz-2,6-biszfoszfát keményítő: ADP-glukóz; szaharóz: UDP-glukóz Trióz-P megoszlás: Pi/trióz-P transzlokátor (antiport) Pi reguláló szerep Regulátor a citoszólban: fruktóz-2,6-biszfoszfát Fr-1,6-BP Fr-6-P átalakulásnál (, inhibitor; , aktivátor)
 Pi reguláló szerep. Regulátor a citoszólban: fruktóz-2,6-biszfoszfát. Fr-1,6-BP Fr-6-P átalakulásnál (, inhibitor; , aktivátor)")
27
A keményítő és szaharóz bioszintézis lépései
Fr2,6,biszfoszfát gátlás

28
A cukorszintézis regulátora: a fruktóz-2,6-biszfoszfát

29
A fotoszintézis élettani és ökofiziológiai vonatkozásai
Fotoszintézis IV. A fotoszintézis élettani és ökofiziológiai vonatkozásai

30
A fotoszintézis fiziológiai és ökológiai vonatkozásai
Limitáló környezeti tényezők: fény, CO2, hőmérséklet -legfontosabb metabolikus lépések: Rubisco aktivitás a RuBP regenerációja trióz-P metabolizmus

31
A jobb fénykihasználást szolgálja a kloroplasztiszok mozgása is
A „szűrőhatás” Felülnézetből: (a) árnyékban; (b) átlagos fényintenzitáson; (c) fénystressznél
 árnyékban; (b) átlagos fényintenzitáson; (c) fénystressznél.")
32
A fotoszintetikus apparátus komponenseinek aránya erős és gyenge fényben

33
A fotoszintézis intenzitása a fényintenzitással növekszik: a kompenzációs pontnál a légzés CO2 termelése egyenlő a fotoszintézis CO2 fogyasztásával

34
Nagy fényintenzitás (fénystressz) esetén reaktív oxigén formák keletkeznek, ami degradálja a fotoszintetikus apparátust: fénygátlás
 esetén reaktív oxigén formák keletkeznek, ami degradálja a fotoszintetikus apparátust: fénygátlás")
35
A széndioxid Biokémiai reakciói a parciális nyomástól (mólfrakció x össznyomás) függenek - az üvegház hatás -kutikula (átjárhatatlan) -gáz fázis: sztómanyílás sztóma alatti üreg sejtközötti légterek - folyadék fázis: sejtfal, citoszól, kloroplasztisz limitáló tényező: a sztómaellenállás (regulálható!)
 -gáz fázis: sztómanyílás. sztóma alatti üreg. sejtközötti légterek. - folyadék fázis: sejtfal, citoszól, kloroplasztisz. limitáló tényező: a sztómaellenállás (regulálható!)")
36
C3-as és C4-es növények fotoszintézise a külső és az intercelluláris tér CO2 koncentrációjának függvényében

37
A C4-es növények fotoszintetikus hatékonyságának hőmérsékleti optimuma magasabb értéknél található, mint a C3-as növényeké

38
A floem transzport Malpighi, 1686. gyűrűzés

40
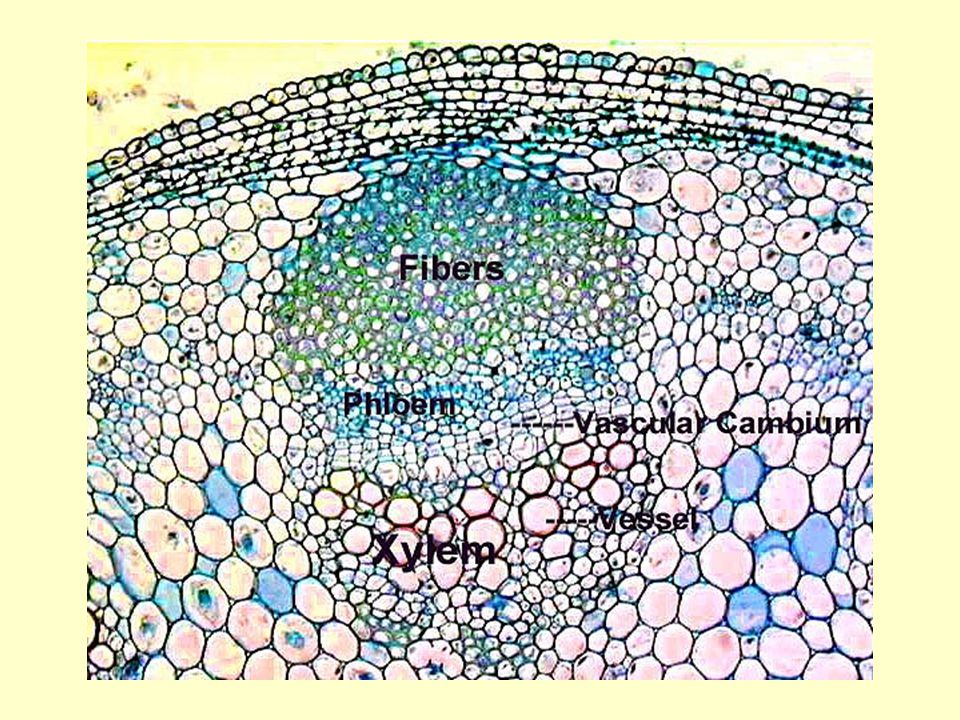
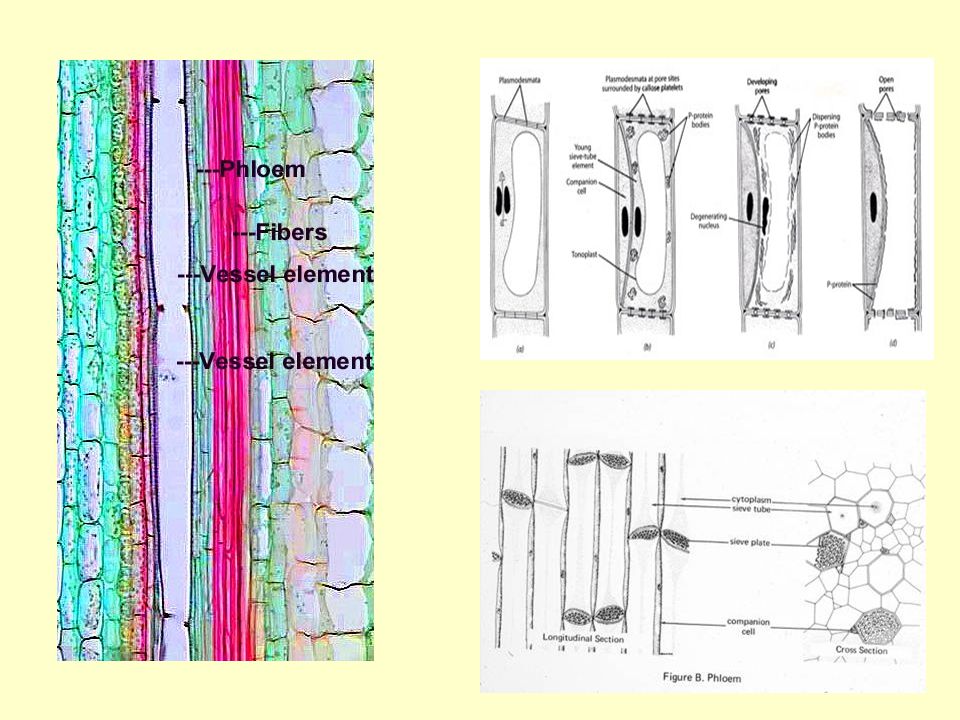
A floem legfontosabb elemei:
- rosta elemek rostacső elem (zárvatermők) → rostacső rostasejt (nyitvatermők) - kísérősejtek (funkcionális kísérők) zárvatermők: egy anyasejt utódai közönséges kísérősejt, transzfer sejtek, intermedier sejtek nyitvatermők: nem egy anyasejt utódai albuminos sejtek - floem parenchima, rostsejtek, latexet tartalmazó sejtek
 → rostacső. rostasejt (nyitvatermők) - kísérősejtek (funkcionális kísérők) zárvatermők: egy anyasejt utódai. közönséges kísérősejt, transzfer sejtek, intermedier sejtek. nyitvatermők: nem egy anyasejt utódai. albuminos sejtek. - floem parenchima, rostsejtek, latexet tartalmazó sejtek.")
42
A rostacső elemek élő sejtek
Nincs: sejtmag, tonoplaszt, mikrotubulusok, mikrofilamentumok, Golgi apparátus, riboszóma Van: PM, mitokondrium, degenerált plasztiszok, sima ER P-protein

43
A kísérősejtek típusai
Transzfer sejt Intermedier sejt Közönséges kísérősejt

44
Anatómiai és fejlődésélettani alapelvek a transzlokáció útvonalának kialakulásában
Közeli pozíció: felső, érett levél → csúcs alsó, érett levél → gyökér A felhasználóhely fontossága: merisztéma → váltás → gyümölcs Közvetlen nyalábösszeköttetés: egymás alatti, fölötti szervek vertikális sorok → ortosztichia Flexibilitás: anasztomózisok, alternatív utak

45
A floemnedv anyagai és analízisének módszerei

46
A floemnedv gyűjtésének módszerei
Hipokotil átvágása: ricinus pozitív nyomás kallóz szintáz gátlása (EDTA) Hátrány: kevés fajnál működik, sebzés hatása Levéltetvek szipókáin keresztül
 Hátrány: kevés fajnál működik, sebzés hatása. Levéltetvek szipókáin keresztül.")
47
Vizes oldat Cukrok: . nem redukálók szacharóz: (~1 M!) raffinóz sor Cukoralkoholok: mannitol Aminosavak: Glu, Gln N-fixálók: amidok, ureidek Hormonok Nem: Ca2+, Fe2+, NO3-, SO42-

48
A floem feltöltődése: a kloroplasztisztól a rostaelemig: rövid távú transzport
lépés: triózfoszfát kilépése a kloroplasztiszból a citoszolba, szacharóz szintézis 2. lépés: a szacharóz transzportja a mezofillumsejtektől a rostacső elemekig 3. lépés: a rostacsőelem feltöltése

49
A feltöltődés mechanizmusai: szállítás hosszú távon
Apoplasztikus út a. / passzív diffúzió b./ aktív transzport ATP-áz a kísérősejtek és a rostacső elemek PM-jén proton-szacharóz kotranszport (SUC2; SUT1) általános
 általános.")
50
A szacharóz leadása a floemből és átalakulása a felhasználóhelyeken

51
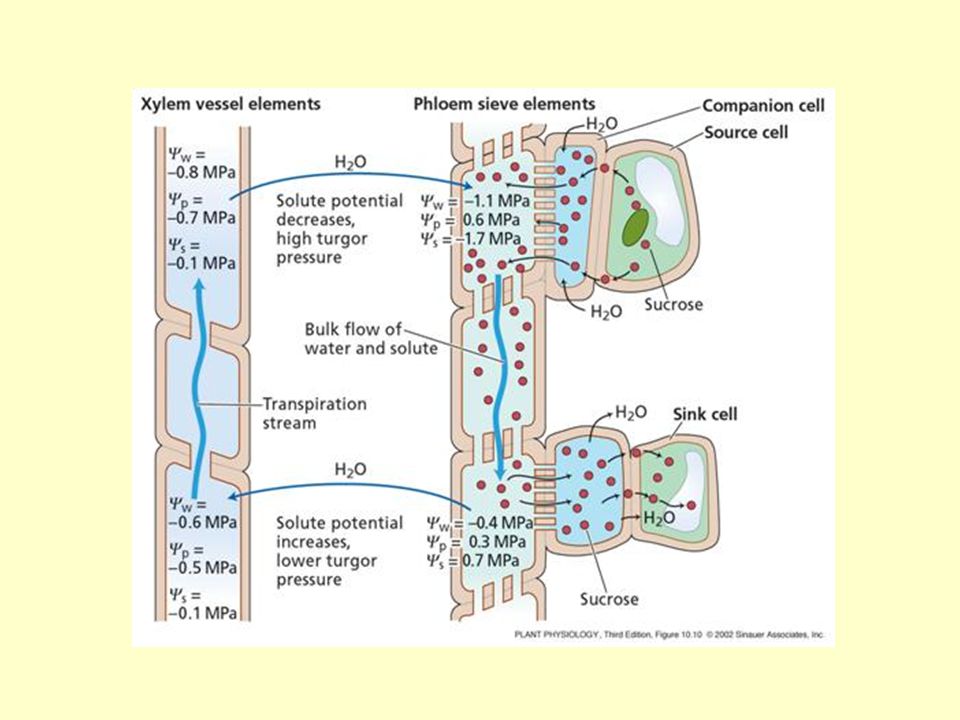
A floemtranszport mechanizmusa
Münch hipotézise, tömegáramlási modell: Feltételek: folyamatos út a rostalemez pórusain nincs kétirányú transzport egy rostacső elemben alacsony energiaigény pozitív nyomásgrádiens

Hasonló előadás