Előadást letölteni
Az előadás letöltése folymat van. Kérjük, várjon

1
A fotoszintézis molekuláris biológiája

2
A fotoszintézis két fő szakasza: a fényreakciók és a sötét reakció

3
A folyamat jelentősége
Az élővilág energiaforrása Szerves anyagok előállítása szervetlenekből Az ábrán a keményítő szemcsék láthatók A Föld légkörének oxigéntartalma ebből a folyamatból ered

4
A Földre érkező napsugárzás spektruma

5
1. Fényszakasz: A fényt fotoszintetikus pigmentek abszorbeálják
A fényt abszorbeáló pigmentek a cianobaktériumokban és magasabbrendű növényekben: Kolorofill a, b Karotinoidok Fikobilinek (cianobaktériumokban és vörösmoszatokban)
")
6
A fotoszintétikus pigmentek elnyelési spektruma a fény látható tartományában (380 és 760 nm) a különböző hullámhosszúságú sugarak felfogása a növényi anyagok pigmentjei által Hatásspektrum (grafikon) a fotoszintézis mértékét mutatja (pl. fotoszintétikus oxigéntermelés), - azt, hogy a különböző hullámhosszúságú fénysugarak mennyire hatékonyak a fotoszinétikus folyamatokra A prizmával alkotóelemeire bontott fehér fény kék és vörös hullámhosszú komponenseivel megvilágított régió köré gyűlnek az aerob baktériumok a fotoszintetizáló Spirogyra körül Elnyelési spektrum hatásspektrum
 a fotoszintézis mértékét mutatja (pl. fotoszintétikus oxigéntermelés), - azt, hogy a különböző hullámhosszúságú fénysugarak mennyire hatékonyak a fotoszinétikus folyamatokra. A prizmával alkotóelemeire bontott fehér fény kék és vörös hullámhosszú komponenseivel megvilágított régió köré gyűlnek az aerob baktériumok a fotoszintetizáló Spirogyra körül. Elnyelési spektrum. hatásspektrum.")
7
Akcióspektrum

8
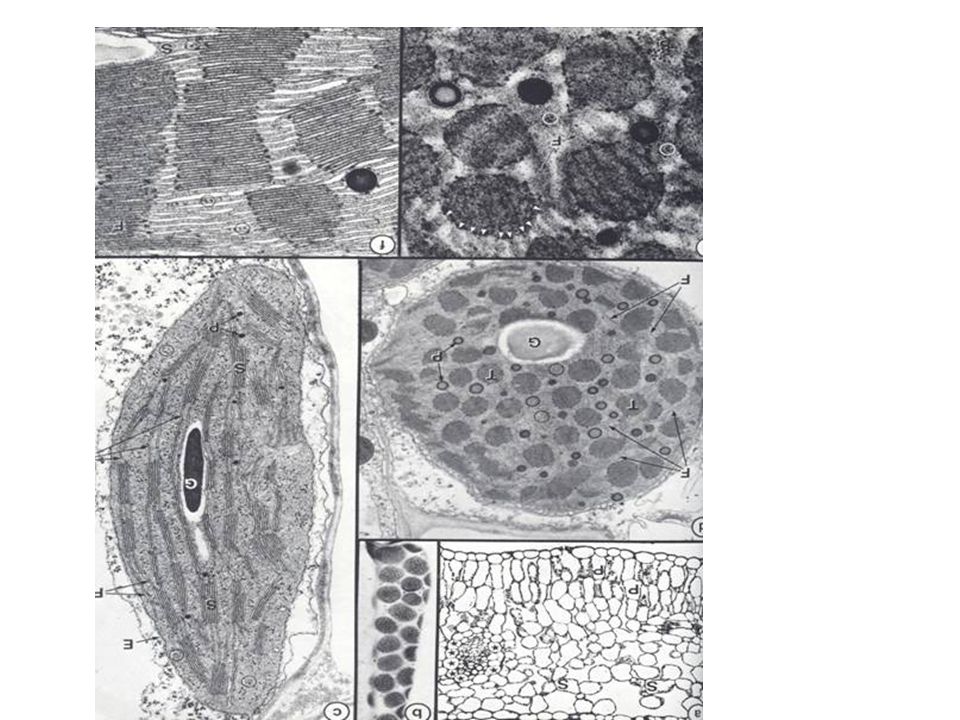
A fotoszinetikus apparátus morfológiája
A fotoszintetikus apparátus alapvető struktúrális egysége a tilakoid. - Ezek önmagukba záródó membránok, melyeknek belső tere az ún. lumen. A prokarióta szervezetekben (pl. cianobaktériumok) a tilakoidok nem határolódnak el a sejt többi részétől. A sejtmagot tartalmazó, eukarióta, szervezetekben (pl. zöld növények) a tilakoidok egy különálló sejtszervben, az ún. kloroplasztiszban találhatók.
 a tilakoidok nem határolódnak el a sejt többi részétől. A sejtmagot tartalmazó, eukarióta, szervezetekben (pl. zöld növények) a tilakoidok egy különálló sejtszervben, az ún. kloroplasztiszban találhatók.")
9
A levél keresztmetszete
a kloroplasztiszok az ún. mezofill sejtekben, valamint a légzőnyílásokat körülvevő záró (ún. quard) sejtekben találhatók C4-es növények esetén a levélereket körülvevő hüvelyparenhima sejtekben is találhatók kloroplasztiszok. A sejtek és kloroplasztiszok elhelyezkedése egy C3-as növényben
 sejtekben találhatók. C4-es növények esetén a levélereket körülvevő hüvelyparenhima sejtekben is találhatók kloroplasztiszok. A sejtek és kloroplasztiszok elhelyezkedése egy C3-as növényben.")
10
A fotoszinetikus apparátus morfológiája
A tilakoidok kétféle szerveződésű formában találhatók: - a granumok külső felületükkel egymáshoz ragadt tilakoidok oszlopszerű képződmények és - a granumok közötti térben - a sztrómában (az őket) összekötő tilakoid lamellák. A klp-iszt kettős membrán veszi körül: - a belső a bekebelezett prokarióta sejt membránja - a külső eukarióta eredetű A kloroplasztisz elektronmikroszkópos képe és szerkezeti modellje
 összekötő tilakoid lamellák. A klp-iszt kettős membrán veszi körül: - a belső a bekebelezett prokarióta sejt membránja. - a külső eukarióta eredetű. A kloroplasztisz. elektronmikroszkópos képe. és szerkezeti modellje.")
11
A fotoszinetikus apparátus morfológiája
A kloroplasztiszok csak fény hatására differenciálódnak a merisztemasejtek proplasztiszaiból. Kialakulásuk során a 2 határolómembrán közül a belső, az organellum hossztengelyével párhuzamosan betüremkedik a sztrómába, majd ezek a belső mebmr.-lemezek lefűződnek, magukba zárva a membránközi (periplasztidális) térből eredő anyagok egy részét, amiből a tilakoidlemezek membránnal körülzárt lumenje jön létre, anélkül, hogy a sztrómával valaha is kapcsolatba kerülne A tilakoidok összefüggő membránrendszert alkotnak. A fotoszintetikus pigmentek csak a tilakoid membr.-ban vannak jelen A tilakoidmembr.-ok szabályos kitüremkedéseket képezve pénztekercs szerűen egymásra tevődött korongokat, ún. granum korongokat hoznak létre - egy granumkorongköteg a gránum. A tilakoid membránok egymást fedik, szorosan egymásra tevődnek (kapcsolt membránok)
 térből eredő anyagok egy részét, amiből a tilakoidlemezek membránnal körülzárt lumenje jön létre, anélkül, hogy a sztrómával valaha is kapcsolatba kerülne. A tilakoidok összefüggő membránrendszert alkotnak. A fotoszintetikus pigmentek csak a tilakoid membr.-ban vannak jelen. A tilakoidmembr.-ok szabályos kitüremkedéseket képezve pénztekercs szerűen egymásra tevődött korongokat, ún. granum korongokat hoznak létre. - egy granumkorongköteg a gránum. A tilakoid membránok egymást fedik, szorosan egymásra tevődnek (kapcsolt membránok)")
12
A kloroplasztisz szerkezete
Folyékony közege a sztróma – benne enzimek, keményítő szemcsék, DNS és riboszómák A sztróma a prokarióta cp-jának felel meg, benne a Calvin ciklus enzimeivel. Membránrendszere tilakoidokból áll – ezek helyenként gránumokba rendeződnek. A tilakoidok belső üregei egy összefüggő, lumen nevezetű hálózatot alkotnak Membránlipidjeik glikozil-gliceridek: nagyobbrészt a MGDG- gránumtilakoidokban, a DGDG-sztrómatilakoidokban helyezkednek el A kloroplasztisz szerkezete

13
- a második fotokémiai rendszer (PSII) főként a gránumokban,
A fotoszint. komplexei (I es II fotokémiai rendszer, citocróm b6f komplex, az ATP-áz és a mobilis fénybegyűjtő antenna) a tilakoid membránba ágyazódva találhatók A fotoszintetikus komplexek eloszlása a tilakoid kompartmentekben nem egyenletes - a második fotokémiai rendszer (PSII) főként a gránumokban, - az első fotokémiai rendszer (PSI) és az ATP-áz főként a sztróma tilakoidokban illetve a gránumok külső részén található. Fény hatására a tilakoid belső terében, a lumenben, megnő a H+ koncentráció, ami az ATP szintézisben hasznosul. A kloroplasztiszon belül jól láthatók a grána- és a sztrómatilakoidok, valamint a nagyméretű keményítő szemcse van
 a tilakoid membránba ágyazódva találhatók. A fotoszintetikus komplexek eloszlása a tilakoid kompartmentekben nem egyenletes. - a második fotokémiai rendszer (PSII) főként a gránumokban, - az első fotokémiai rendszer (PSI) és az ATP-áz főként a sztróma tilakoidokban illetve a gránumok külső részén található. Fény hatására a tilakoid belső terében, a lumenben, megnő a H+ koncentráció, ami az ATP szintézisben hasznosul. A kloroplasztiszon belül jól láthatók a grána- és a sztrómatilakoidok, valamint a nagyméretű keményítő szemcse van.")
15
A fotoszinetikus apparátus morfológiája
A fotoszintézis fényszakaszának folyamataiban a tilakoid-membránok 5-féle szupramolekuláris fehérjeegyüttese vesz részt: -A kettes fotokémiai rendszer PS II (photosystem II), a hozzá kapcsolódó vízbontó komplexszel (Water-splitting complex-WPC, v. Oxigen evolving complex -OEC), a citocróm b6/F komplex, Az egyes fotokémiai rendszer PS I (photosystem I), A kétféle fotokémiai rendszerhez kapcsolódó fénybegyűjtő pigment-protein komplex (Light harvesting complex –LHC), A kloroplasztban levő ATP-szintetáz kapcsolófaktor (CF1-CFo v. CF1Fo) komplex
, a hozzá kapcsolódó vízbontó komplexszel (Water-splitting complex-WPC, v. Oxigen evolving complex -OEC), a citocróm b6/F komplex, Az egyes fotokémiai rendszer PS I (photosystem I), A kétféle fotokémiai rendszerhez kapcsolódó fénybegyűjtő pigment-protein komplex (Light harvesting complex –LHC), A kloroplasztban levő ATP-szintetáz kapcsolófaktor (CF1-CFo v. CF1Fo) komplex.")
16
A fénybegyűjtő pigmentek: Klorofill
Porfirin gyűrű – négy pirrolgyűrűből áll A négy pirrolgyűrű mellett egy öttagú ciklopentanon gyűrű található A konjugációs rendszer kékkel jelölve Magnézium ion – Mg++ - narancs Észter kötéssel csatlakozó fitol – zöld Ez a klorofill a – a többi klorofill kicsit különbözik. porfirin propionsavgyök

17
A fotoszintetikus szervezetek legjellemzőbb pigmentjei a klorofillok, amelyek a kék ill. vörös tartományban nyelnek el. A gyűrű közepén egy Mg atom található, a 4-es számú pirrolgyűrűhöz pedig egy 20 szénatomos alkohol, a fitol, csatlakozik A klorofillok - első gerjesztett állapotához vörös (430nm), - második gerjesztett állapotához pedig kék (660 nm) abszorpciós sáv tartozik Klorofillok A klorofill-a és klorofill-b szerkezete
, - második gerjesztett állapotához pedig kék (660 nm) abszorpciós sáv tartozik. Klorofillok. A klorofill-a és klorofill-b szerkezete.")
18
A karotinoidok járulékos pigmentek
Az oxigéntermelő szervezetek reakció-centrumaiban, valamint a prokarióták antennáiban a klorofillok mellett karotinok is találhatók, amelyek a kék tartományban ( nm körül) nyelnek el. A karotinok ún. járulékos pigmentek - feladatuk: - a hatékonyabb fényelnyelés elősegítése, - a klorofill molekulákat védik a gerjesztett állapotban bekövetkező fénykárosodástól.
 nyelnek el. A karotinok ún. járulékos pigmentek - feladatuk: - a hatékonyabb fényelnyelés elősegítése, - a klorofill molekulákat védik a gerjesztett állapotban. bekövetkező fénykárosodástól.")
19
A kloroplasztisz szerkezete
A tilakoidok két formája a sztróma tilakoid és a grána tilakoid. Ez a két forma a molekuláris szinten is különbözik.

20
A pigment-protein komplexek rendezetten helyezkednek a tilakoidmembránban

21
A fényenergia abszorpciója
A gerjesztett állapot energiája kisugárzódhat: fény (fluoreszcencia) vagy hő formájában, melyek a fotoszintézis szempontjából veszteséget jelenthet. Átadódhat szomszédos általában alacsonyabb gerjesztési energiájú pigment molekuláknak, igen nagy szerepeük van a gerjesztési energiának a reakciócentrumhoz való továbbításában. Illetve, a gerjesztett állapotban lejátszódhat a töltésszétválást eredményező fotokémiai reakció A fényenergia abszorpciója
 vagy. hő formájában, melyek a fotoszintézis szempontjából veszteséget jelenthet. Átadódhat szomszédos általában alacsonyabb gerjesztési energiájú pigment molekuláknak, igen nagy szerepeük van. a gerjesztési energiának a reakciócentrumhoz való továbbításában. Illetve, a gerjesztett állapotban lejátszódhat a töltésszétválást eredményező fotokémiai reakció. A fényenergia abszorpciója.")
23
Az oxigéntermelő fotoszintetizáló szervezetekben univerzálisan megtalálható:
- a klorofll-a, amelynek kék ill. vörös abszorbciós maximuma 430 ill. 660 nm körül van. Ezek a molekulák a fénybegyűjtés mellett fotokémiai reakcióra is képesek. Mind a PSI, mind a PSII reakciócentrumában speciális klorofill-a molekulák, valószínűleg dimerek vagy multimerek, alkotják a fényindukált töltésszétválásban résztvevő ún. reakciócentrum klorofillokat. A prokarióta cianobaktériumokban csak klorofill-a fordul elő. Az eukarióta fotoszintetizáló szervezetek antenna komplexeiben azonban a klorofill-a mellett egy másik tipusú klorofill is található, ami a klorofill-a kék és vörös abszorpciós sávjai közötti spektrumtartomány jobb lefedését szolgálja. Magasabb rendű növényekben és zöldalgákban az extra klorofill a klorofill-b, ami a klorfill-a-tól csak annyiban különbözik, hogy a második pirrolgyűrűhöz nem metil, hanem formil csoport kötődik. A barna- és kovamoszatokban a klorofill-b helyett klorofill-c, a vörös algákban pedig klorofill-d található.

24
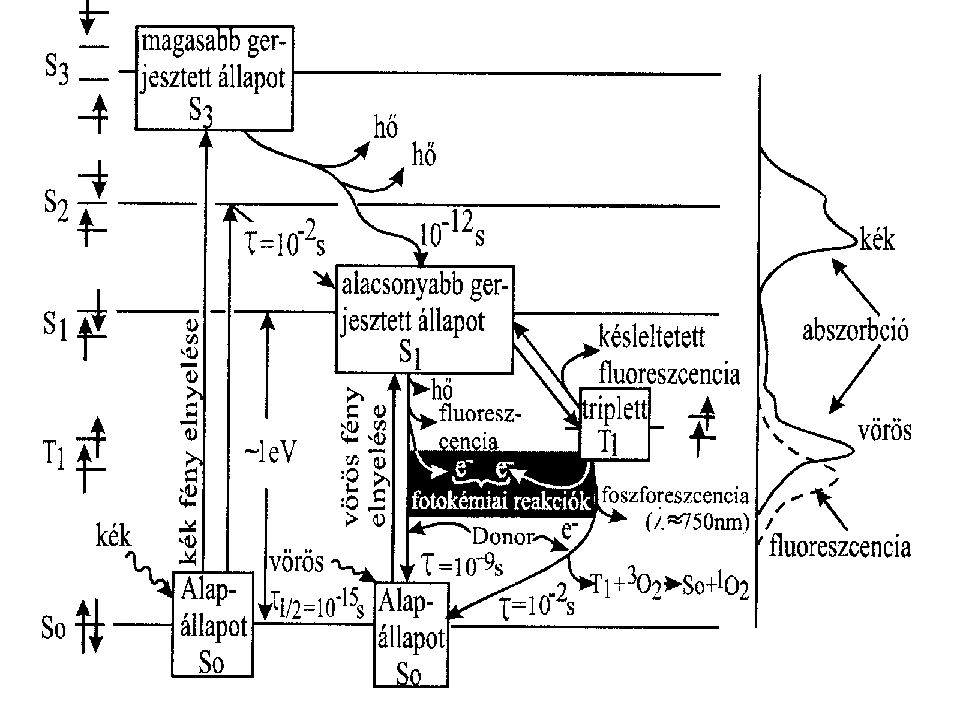
A fény abszorpciója és a gerjesztési energia sorsa
Fotoszintetikusan aktív fény ~ nm-ig Klorofill molekula: alapállapot (S0), Egy p-elektron kötő pályáról lazító pályára lép. szinglet (S1- és S2-) és triplet (T1) állapotok vörös fény elnyelése: S1 állapot kék fény elnyelése: S2 állapot A gerjesztési energia sorsa: hővé alakul, fluoreszcencia, fotokémiai reakciók (redox folyamatok) energia migráció vagy energia transzfer (azonos vagy kémiailag eltérő pigmenteknek adódik át az energia)
, Egy p-elektron kötő pályáról lazító pályára lép. szinglet (S1- és S2-) és triplet (T1) állapotok vörös fény elnyelése: S1 állapot. kék fény elnyelése: S2 állapot. A gerjesztési energia sorsa: hővé alakul, fluoreszcencia, fotokémiai reakciók (redox folyamatok) energia migráció vagy energia transzfer (azonos vagy kémiailag eltérő pigmenteknek adódik át az energia)")
25
A klorofill fluoreszcenciája akkor is a vörös hullámhossz tartományba esik, ha kék fényt nyel el
Ok: az S2-S1 átmenet mindig hő formájában disszipálódik

26
Az antennapigmentek az elnyelt fényenergiát a reakciócentrum klorofill a felé továbbítják
A fotoszintézis első lépése a fény elnyelése, ami fehérjékhez kötött pigment molekulák révén történik. A pigmentekben bekövetkező fényelnyelés a molekulát gerjesztett állapotba hozza (ez a fotoszintetikus pigmenteknél az 1. vagy 2. szinglett gerjesztett állapot). Energiagazdag RC Energiaszegény A fényenergia vándorlása: antenna pigmentek között: külső és belső antennák reakciócentrumok klorofill a molekulájához: II. fotokémiai rendszer: P680, I. fotokémiai rendszer: P700
. Energiagazdag. RC. Energiaszegény. A fényenergia vándorlása: antenna pigmentek között: külső és belső antennák. reakciócentrumok klorofill a molekulájához: II. fotokémiai rendszer: P680, I. fotokémiai rendszer: P700.")
27
Energiaátadás két pigment molekula között akkor történik, ha a donor molekula abszorpciós vagy fluoreszcencia spektruma részben fedi egymást

28
Energia migráció, energia transzfer és fotokémiai folyamatok
Az antenna klorofillok gerjesztett állapotba kerülnek a fényabszorpció következményeként Energia átadás: rezonancia Energia továbbadás elektron átadás által oxidáció és redukció

29
A zöld növények, algák és cianobaktériumok fotoszintézisében elsősorban hasznosuló spektrumtartomány 400 és 700 nm között található, amit fotoszintetikusan aktív sugárzásnak (Photosynthetically Active Radiation, PAR) neveznek. A nemoxigéntermelő baktérimokban található baktérioklorofillok abszoipciója az infravörös tartomány felé tolódott ( nm) a víz alatti fényviszonyokhoz való alkalmazkodás eredményeként. A fénybegyűjtő pigmentek protein vázhoz kötődve antenna komplexekbe szerveződnek. Mind a PSI mind a PSII komplex rendelkezik belső antennával, ami a reakciócentrum része illetve ahhoz szorosan csatlakozik. Mindkét fotokémiai rendszerhez tartozik még egy-egy külső antenna komplex is. - LHCI (Light Harvesting Chlorophylls) az LHCI az első fotokémiai rendszerhez kötődik, míg az - LHCII, a fotoszintézis fényigényétől függően a PS II-höz kapcsolódik vagy leválik róla, ilymódon szabályozva a PSII-höz juttatott energia mennyiségét.
 a víz alatti fényviszonyokhoz való alkalmazkodás eredményeként. A fénybegyűjtő pigmentek protein vázhoz kötődve antenna komplexekbe szerveződnek. Mind a PSI mind a PSII komplex rendelkezik belső antennával, ami a reakciócentrum része illetve ahhoz szorosan csatlakozik. Mindkét fotokémiai rendszerhez tartozik még egy-egy külső antenna komplex is. - LHCI (Light Harvesting Chlorophylls) az LHCI az első fotokémiai rendszerhez kötődik, míg az. - LHCII, a fotoszintézis fényigényétől függően a PS II-höz kapcsolódik vagy leválik róla, ilymódon szabályozva a PSII-höz juttatott energia mennyiségét.")
30
A fénygyűjtő komplex és a reakciócentrum
A fénygyűjtő komplex (LHC II. és I.) a kísérőpigmentek által elnyelt fényenergiát a reakciócentrum klorofillokhoz közvetíti. A fénybegyűjtő komplexek pigmentjei magasabbrendű növényekben klorofill a és b, karotin és xantofill molekulák, gyakran fehérjékhez kötődve találhatók. A gerjesztett pigmentek energiája rezonancia által átadódik a reakciócentrumba. A fényenergia kémiai energiává alakulása: töltésszeparációval Donor Kla Akceptor; Donor Kla Akceptor Donor Kla Akceptor; Donor Kla Akceptor
 a kísérőpigmentek által elnyelt fényenergiát a reakciócentrum klorofillokhoz közvetíti. A fénybegyűjtő komplexek pigmentjei magasabbrendű növényekben klorofill a és b, karotin és xantofill molekulák, gyakran fehérjékhez kötődve találhatók. A gerjesztett pigmentek energiája rezonancia által átadódik a reakciócentrumba. A fényenergia kémiai energiává alakulása: töltésszeparációval. Donor Kla Akceptor; Donor Kla Akceptor. Donor Kla Akceptor; Donor Kla Akceptor")
31
Energiaszint változások a fotoszintézis folyamán
Minél nagyobb az anyag redukálóképessége, annál negatívabb a redox potenciálja Az első fényreakció a PS II ben történik (680 nm) Az elektron energiája csökken az elektrontranszport során Újragerjesztés a PS I-ben (700 nm). Energia tárolás ATP és NADPH formájában
 Az elektron energiája csökken az elektrontranszport során. Újragerjesztés a PS I-ben (700 nm). Energia tárolás ATP és NADPH formájában.")
32
Az elektrontranszportlánc két formája
A nem ciklusos elektrontranszport (Z séma) A ciklikus variációban a ferredoxin a citokróm b6/f komplexnek adja át az elektront. A fény ily módon a PSI egyedüli részvételével körbe hajtja az elektronokat és így hoz létre H+ koncentráció-különbséget a sztróma és a luminális tér között. Ez ATP szintézisre használódik fel.
 A ciklikus variációban a ferredoxin a citokróm b6/f komplexnek adja át az elektront. A fény ily módon a PSI egyedüli részvételével körbe hajtja az elektronokat és így hoz létre H+ koncentráció-különbséget a sztróma és a luminális tér között. Ez ATP szintézisre használódik fel.")
33
ferodoxin

34
ehhez kapcsolódnak a feofitin (primer e- akceptor),
1. A PS II felépítése - reakciócentrum: D1 és D2 fehérjék: ehhez kapcsolódnak a feofitin (primer e- akceptor), a QA és QB kinon akceptorok, D1 fehérje tirozin aminosava (primer e- donor Mn2+, Mn3+ ionok (vízbontás) - reguláló sapka - proximális antenna - disztális antenna 2. Funkciói vízbontás: 2H2O O2 + 4H+ + 4e- proton keletkezik a lumenben, elektron lép az e- transzportláncba a plasztokinon redukciója
, a QA és QB kinon akceptorok, D1 fehérje 161. tirozin aminosava (primer e- donor. Mn2+, Mn3+ ionok (vízbontás) - reguláló sapka. - proximális antenna. - disztális antenna. 2. Funkciói. vízbontás: 2H2O O2 + 4H+ + 4e- proton keletkezik a lumenben, elektron lép az e- transzportláncba. a plasztokinon redukciója.")
35
A magasabbrendű fotoszintetizáló szervezetek (zöld növények, algák és a cianobaktériumok) a természetben egyedülálló képessége a víz fényindukált elbontása protonokká és molekuláris oxigénné. E folyamat révén a Földön kimeríthetetlen mennyiségben rendelkezésre álló víz szolgál a fotoszintetikus elektrontranszport forrásaként. Melléktermékként pedig a magasabbrendű szervezetek számára nélkülözhetetlen oxigén keletkezik. A fényindukált vízbontást a tilakoid membránba ágyazódott nagyméretű pigment-protein komplex, az ún. második fotokémiai rendszer (PSII) katalizálja. - a folyamat során keletkező elektronokat a membrán lipid fázisában diffundáló plasztokinon (PQ) molekulák továbbítják a citokróm bf komplex felé, - a tilakoid lumenbe bocsátott protonok pedig a transzmembrán proton gradiens kialatításához járulnak hozzá.
 katalizálja. - a folyamat során keletkező elektronokat a membrán lipid fázisában diffundáló plasztokinon (PQ) molekulák továbbítják a citokróm bf komplex felé, - a tilakoid lumenbe bocsátott protonok pedig a transzmembrán proton gradiens kialatításához járulnak hozzá.")
36
A másodikfotókémiai rendszer szerepe a fotoszintetikus elektrontranszportban

37
A PSII reakciócentrumát a D1 és D2 fehérjékből álló heterodimer alkotja, amihez kötődnek a fényindukált elektrontranszport redox komponensei. A reakciócentrumhoz közvetlenül csatlakozik, és feltehetően annak integráns részét képezi a citokróm b-559. Ez a redox fehérje két alegységből áll és egy hem csoportot tarralmaz. A PSII belső antennáját két, klorofill-a-t kötő protein alegység, a CP43 és CP47 alkotja. Külső antennaként a klorofill-a-t és klorofill-b-t kötő LHCII (vagy LHCb) szolgál, ami a PSII komplexhez elektrosztatikus kölcsönhatás révén kapcsolódik. A redox-aktív és klorofillkötő alegységek mellett a PSII tartalmaz még számos kisebb méretű, membránba ágyazódott alegységet is, amelyek feltehetően strukturális szerepet játszanak. A komplex lumenális oldalán található három vizoldékony protein, amelyek a vízbontó rendszer kialakításában játszanak strukturális szerepet.
 szolgál, ami a PSII komplexhez elektrosztatikus kölcsönhatás révén kapcsolódik. A redox-aktív és klorofillkötő alegységek mellett a PSII tartalmaz még számos kisebb méretű, membránba ágyazódott alegységet is, amelyek feltehetően strukturális szerepet játszanak. A komplex lumenális oldalán található három vizoldékony protein, amelyek a vízbontó rendszer kialakításában játszanak strukturális szerepet.")
38
A PSII szerkezete kicserélni

39
Az elektrontranszportlánc felépítése
- membránhoz kötött komponensek PS II, citokróm b6/f komplex, PS I, ferredoxin-NADP+ reduktáz - mobilis komponensek plasztokinon pool, PQH2 (membránban, PS II és citokróm b6/f komplex között) plasztocianin, PC (lumen, citokróm b6/f komplex és PS I. közt) ferredoxin (PS I. és ferredoxin-NADP+ reduktáz között)
 plasztocianin, PC (lumen, citokróm b6/f komplex és. PS I. közt) ferredoxin (PS I. és ferredoxin-NADP+ reduktáz. között)")
40
A citokróm b6/f komplex felépítése
A két fotokémiai rendszer közötti elektrontranszportot a citokróm b6f komplex közvetíti. Funkciója a kétszeresen redukált plasztokinon PQH2 oxidációja és az elektronok továbbítása a plasztocianin közvetítésével a PSI felé. Ezen elektrontranszport folyamat során a cit. b6f complex protont pumpál a sztrómából a tilakoid lumenbe és így fontos szerepet játszik a membrán két oldala között fény hatására kialkuló proton gradiens létrehozásában. A komplexnek három, redox funkcióval rendelkező alegysége van: - a citokróm-bb, ami egy alacsony és magas potenciálú hem csoportot (bL és bH) tartalmaz; - a citokróm-f, ami egy hem csoportot csoportot tartalmaz; továbbá - egy vas-kén protein, aminek redox-aktív eleme - egy Fe2S2 centrum.
 tartalmaz; - a citokróm-f, ami egy hem csoportot csoportot tartalmaz; továbbá. - egy vas-kén protein, aminek redox-aktív eleme. - egy Fe2S2 centrum.")
41
citokróm b6f komplex Az izolált komplex dimer szerveződést mutat, amelynek teljes molekulatömege kb. 210 kDa. A dimer forma aktivitása kb. 5-6-szor nagyobb mint a monomer formáé, ami arra utal, hogy a tilakoid membránban található aktív komplex is dimer. A b6f komplex érdekes tulajdonsága, hogy egy szorosan kötődő klorofill-a molekulát tartalmaz, amelynek szerepe egyenlőre nem ismert.

42
A citokróm b6/f komplex működése
oxidálja a PQH2-t, redukálja a PC-t a ciklusos elektrontranszportban oxidálja a ferredoxint proton átadást közvetít a sztrómából a lumenbe

43
Az első (PSI) fotokémiai rendszer
Az oxigéntermelő fotoszintetikus szervezetekben két fotokémiai rendszer működik: - a második fotokémiai rendszer funkciója a víz fényindukált elbontása. E folyamatból származó elektronok mobilis plasztokinon molekulák közvetítésével, a citokróm b6f komplexen keresztül jutnak az első fotokémiai rendszerhez (PSI). A PSI alapvető funkciója a Calvin ciklusban hasznosuló NADPH termelése, ami PSI lineáris elektrontranszportjának eredménye. Emellett a PSI körül lejátszódhat ciklikus elektrontranszport is, amelynek során protonok pumpálódnak a sztrómából a tilakoid lumenbe. Az így kialakult H+ grádiens ATP szintézisben hasznosul.
. A PSI alapvető funkciója a Calvin ciklusban hasznosuló NADPH termelése, ami PSI lineáris elektrontranszportjának eredménye. Emellett a PSI körül lejátszódhat ciklikus elektrontranszport is, amelynek során protonok pumpálódnak a sztrómából a tilakoid lumenbe. Az így kialakult H+ grádiens ATP szintézisben hasznosul.")
44
A PS I. felépítése és működése
Reakciócentrum: 2 db centrális fehérje P700 klorofill-a elsődleges (Kla) és másodlagos (K-vitamin) akceptorok vas-kén centrumok PC és ferredoxin kötő fehérjék Nem ciklusos elektrontranszport: az e- a NADP+ felé, Ciklusos elektrontranszport: e- a citokróm b6/f komplex felé Ferredoxin, ferredoxin-NADP+ reduktáz, NADP+ Eredmény: NADPH + H+
 és másodlagos (K-vitamin) akceptorok. vas-kén centrumok. PC és ferredoxin kötő fehérjék. Nem ciklusos elektrontranszport: az e- a NADP+ felé, Ciklusos elektrontranszport: e- a citokróm b6/f komplex felé. Ferredoxin, ferredoxin-NADP+ reduktáz, NADP+ Eredmény: NADPH + H+")
45
A PSI szerepe a ciklikus (elektrontranszportban

46
A PSI szerepe a lineáris elektrontranszportban

47
A H+-koncentrációkülönbség és az ATP képződés
A két folyamat a mitokondriumban és a kloroplasztiszban hasonló módon játszódik le. Mindkét esetben az ATP szintézis egy protontranszlokáló enzim, az ATP szintáz, segítségével megy végbe. A protonáramlás a magasabb, lúgosabb pH felé tart Az enzim aktív helyei a „gomba fejében” helyezkednek el. Az ATP a mitokondriumi matrixban illetve a kloroplasztisz beli sztrómában keletkezik.

48
A fotoszintetikus foszforiláció: kemiozmotikus modell
proton elektrokémiai potenciálgrádiens a lumen és a sztróma között: ezt használja fel az ATP- szintáz enzim az ATP szintézisére ADP-ből és anorganikus foszfátból

49
Az ATP szintézis folyamatának sémája, amely szerint az ATP-áz működésének hajtóereje a transzmembrán proton gradiens

50
2. A fotoszintézis sötét szakasza: Calvin ciklus
Jelzett, H14CO3- adagolásával a széndioxid fixálás elsődleges termékei kimutathatók

51
Szétkapcsoló anyagok megszüntetik ezt a szoros kapcsolatot.
Az elektontranszportlánc és a foszforiláció A két folyamat között olyan szoros a kapcsolat hogy ATP-képződés nélkül az elektrontanszportlánc leáll és fordítva. Szétkapcsoló anyagok megszüntetik ezt a szoros kapcsolatot. Ilyen szétkapcsoló anyagok a következők: CCCP, valinomicin, gramicidin D, DCPIP stb.

52
A sötét szakasz legfontosabb biokémiai folyamatai
- karboxilációs fázis ribulóz-1,5-bifoszfát + CO2 = 2 glicerinsav-3-foszfát Katalizálja: a ribulóz-1,5-bifoszfát-karboxiláz, oxigenáz (RUBISCO) - redukció szakasza foszforiláció, az ATP 2/3 részének felhasználása, redukció, a redukált NADPH felhasználása a glicerinsav-3- foszfát redukciójára - regeneráció a ribulóz-1,5-bifoszfát-karboxiláz regenerálása: 6 glicerinsav-3-foszfát 3 ribulóz-1,5-bifoszfát+1 trióz- foszfát Nyereség ciklusonként egy triózfoszfát!
 - redukció szakasza. foszforiláció, az ATP 2/3 részének felhasználása, redukció, a redukált NADPH felhasználása a glicerinsav-3- foszfát redukciójára. - regeneráció. a ribulóz-1,5-bifoszfát-karboxiláz regenerálása: 6 glicerinsav-3-foszfát 3 ribulóz-1,5-bifoszfát+1 trióz- foszfát. Nyereség ciklusonként egy triózfoszfát!")
53
RUBISCO Ribulóz-1,5-biszfoszfát-karboxiláz-oxigenáz a reakciót katalizáló enzim Kettős enzimaktivitású – karboxiláz és oxigenáz (3:1) Kompetíció, ha mindkét szubsztrátum jelen van.

54
A RUBISCO SZERKEZETE

55
A sötétszakasz kezdete: karboxiláció
A Calvin ciklus folyamán a C3-as úton a széndioxid a ribulóz 1,5 biszfoszfáthoz kötődik és egy feltételezett hat szénatomos intermedier termék közbeejtésével két molekula glicerinsav-3-foszfáttá alakul át. A glicerinsav-3-foszfát a fotoszintetikus CO2-fixáció első stabil terméke

56
A Calvin ciklus jelentősége
Három pentózfoszfátból 3 molekula széndioxid megkötésével 6 triózfoszfát keletkezik A három pentózfoszfát regenerálódik A 3 széndioxid molekulából a nettó eredmény 1 exportálódó triózfoszfát Ez cukrok (szacharóz), zsírsavak, aminosavak szintézisére szolgál A folyamat során 9 ATP és 6 NADPH használódik fel
, zsírsavak, aminosavak szintézisére szolgál. A folyamat során 9 ATP és 6 NADPH használódik fel.")
57
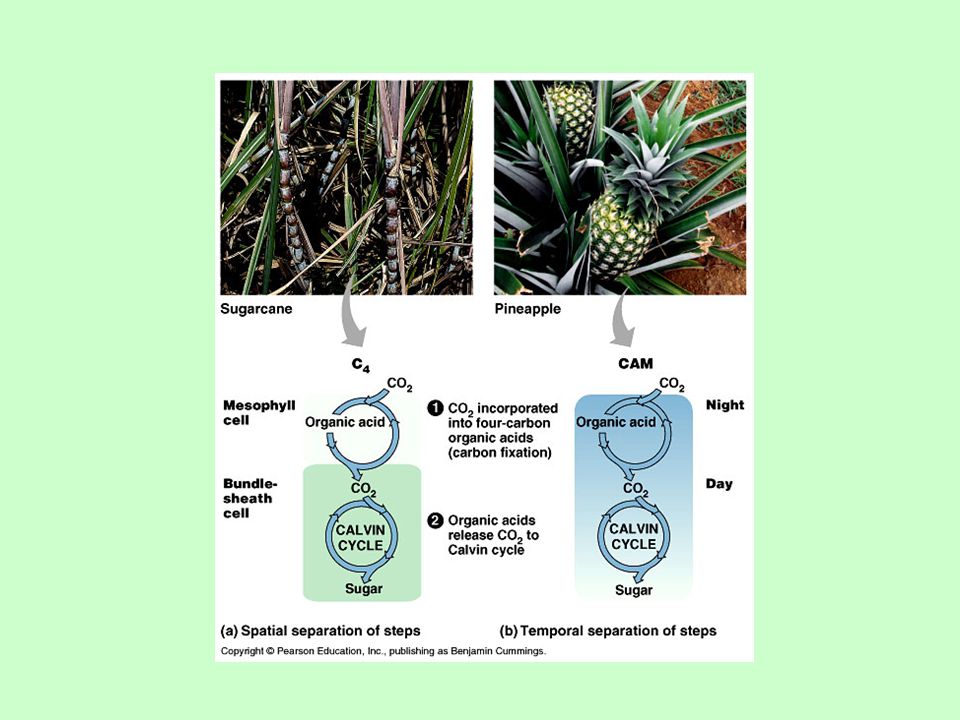
C3 és C4 levélszerkezet A C4-es növények anatómiája szembeszökően különbözik a C3-as növényekétől. Csak a C4-es növényekben találhatók a jellegzetes morfológiájú hüvelyparenchima (bundle sheath) vagy Kranz sejtek. A széndioxid elsődleges kötését a foszfoenol-piroszőlősav (PEP) karboxiláz enzim katalizálja a mezofill sejtek citoplazmájában.
 vagy Kranz sejtek. A széndioxid elsődleges kötését a foszfoenol-piroszőlősav (PEP) karboxiláz enzim katalizálja a mezofill sejtek citoplazmájában.")
60
Nagy fényintenzitás (fénystressz) esetén reaktív oxigén formák keletkeznek, ami degradálja a fotoszintetikus apparátust: fénygátlás
 esetén reaktív oxigén formák keletkeznek, ami degradálja a fotoszintetikus apparátust: fénygátlás")
61
Az elektrontranszportlánc specifikus gátlószerei gyakran fotoszintézisgátló növényvédőszerként kerülnek forgalomba DCMU: jelenleg is használatos Paraquat: kivonták a forgalomból

62
A fénylevelek nagyméretű oszlopos alapszövettel és több sejtsoros mezofillummal rendelkeznek: így optimálisan hasznosítják a fényt. Az árnyéklevelekben a fénybegyűjtő komplex aránya nagyobb a reakciócentrumhoz viszonyítva, mint a fénylevelekben.

63
A jobb fénykihasználást szolgálja a kloroplasztiszok mozgása is
Felülnézetből: (a) árnyékban; (b) átlagos fényintenzitáson; (c) fénystressznél
 árnyékban; (b) átlagos fényintenzitáson; (c) fénystressznél.")
64
Sok fajnál a levéllemez fénykövető mozgást mutat: heliotropizmus

65
A fotoszintézis intenzitása a fényintenzitással növekszik: a kompenzációs pontnál a légzés CO2 termelése egyenlő a fotoszintézis CO2 fogyasztásával

66
A fénynövények fotoszintetikus hatékonysága magasabb telítődési értéket mutat és kompenzációs pontjuk is magasabban van, mint az árnyéknövényeké

67
A C4-es növények fotoszintetikus hatékonyságának hőmérsékleti optimuma magasabb értéknél található, mint a C3-as növényeké

Hasonló előadás












 gáz, vagy gázelegy Lézerátmenet: elektronszintek között (UV és látható lézerek) rezgési szintek.>")







>")