B SEJT DIFFERENCIÁCIÓ A CSONTVELŐBEN
SZABÁLYOZOTT B-SEJT DIFFERENCIÁCIÓ Éretlen-B sejt Pre-B sejt ANTIGÉN FELISMERŐ ÉS JELÁTVÍVŐ RECEPTOR Pro-B sejt ÁTRENDEZETT H+PÓT L JELÁTVIVŐ RECEPTOR NINCS ANTIGÉN FELISMERŐ RECEPTOR
A differenciáció szakaszait a csontvelőben a génátrendeződés foka is jelzi Pre-B sejt receptor kifejeződése B SEJT ÁLLAPOT IgH GÉN KONFIGURÁCIÓ Őssejt Korai pro-B Késői pro-B Nagy pre-B Csíravonal DH to JH VH to DHJH VHDHJH Az Ig L-lánc még nincs átrendezve
Fejlődő B sejtek Csontvelői sztróma sejtek
B B Stroma sejt
A CSONTVELŐI SZTRÓMA SEJTEK TÁPLÁLJÁK/SEGÍTIK A FEJLŐDŐ B- LIMFOCITÁKAT 1. Specifikus sejt-sejt kölcsönhatás a sztróma sejtek és a B-sejtek között Sejt-sejt kölcsönhatás Termelt faktorok – CITOKINEK Kolónia stimuláló faktorok Növekedési faktorok Differenciációs faktorok 2. A sztróma sejtek citokineket termelnek B Sztróma sejt A citokinek és a közvetlen sejt-sejt kölcsönhatások a differenciáció során változnak
A fejlődés egyes szakaszaiban a citokinek és sejtkölcsönhatások változnak HSC Korai pro-B c-Kit Receptor Tyrosine kináz Stem cell factor (SCF) Sejthez kötött növekedési faktor VLA-4 (Integrin) Sejt adhéziós molekulák VCAM-1 (Ig superfamily) Sztróma sejt
A fejlődés egyes szakaszaiban a citokinek és sejtkölcsönhatások változnak Interleukin-7 receptor Interleukin-7 Növekedési faktor Korai pro-B VLA-4 (Integrin) VCAM-1 (Ig superfamily) Késői pro-B Pre-B Sztróma sejt
Nem kell tudni Development A korai limfocita alakok IL receptorokon keresztüli túlélő szignálokat igényelnek. Ez hatással van a progenitorok számára, és az antigén receptor átrendeződésre is. IL7 kell mind a T mind B progenitoroknak. Jak1, 3; Ras, MAPK, útvonalakat aktivál. IL7, yc lánca vagy JAK-3 kiütés a erősen gátolja a limfocita fejlődést. BCL-2 családot szabályoz. BCL-2 overexpresszió helyreállítja T-sejtek esetében (de nem B) az IL7 hiány okozta fejlődési hibát. BCL-2, BCLxL sem kell a fejlődéshez. MCL-1 hiánya növeli az apoptózist, megállítja a fejlődést az átrendeződés előtt, mint az IL7 hiánya. MCL-1 rövid félélet idejű antiapototikus BCL-2 család tag. Ubiquitnnel bomlik. Foszforiláción ill ubin kersztül gyorsan szabályozódhat. Proapoptotikus? Bad def nincs hatása, Bim deficiens rezisztens a növekedési faktor megvonásra. IL7 hiány tüneteit a Bim hiány csaknem teljesen helyreállítja. PUMA deficiens is hasonló hatású. Szineregiztikusak. (B-sejen a PUMának nincs hatása, csak Bim) Bax hiány részlegesen helyreállít, (de nem Bsejteken) azaz Bim aktiválja Bax-ot. Bax, Bak kettős kiütés teljesen rezisztens B és T sejteken is növekedési faktor hiányra. Nem kell tudni
Y A B sejt fejlődés szakaszai Nagy pre-B sejt Pre-receptor Őssejt - HSC Korai pro-B sejt Késői pro-B sejt Csíravonal DH to JH VH to DHJH VHDHJH Perifériás érett B sejt Kis pre-B sejt Éretlen B sejt Y Receptor H+L Az egyes érési fázisokat sejtfelszíni markerek megjelenése (vagy eltűnése) jellemzi
Pre- B sejt receptor H-lánc VHDHJH V-preB l5 CHm Iga & Igb jelátvivő molekulák Átmenetileg jelenik meg a membránban produktív VHDHJH CHm átrendeződés után VpreB/l5 – pót L-lánc, a membrán kifejeződéshez szükséges A pre-B sejt receptor liganduma nem ismert?
A nagy pre-B sejtek további éréséhez helyes keretű VHDHJH kapcsolat szükséges Large pre-B További differenciálódás Pre-B cell receptor kapcsolódik a sztróma sejthez Human IgG3 Heavy Chain nucleotide sequence ATGAAACANCTGTGGTTCTTCCTTCTCCTGGTGGCAGCTCCCAGATGGGTCCTGTCCCAGGTGCACCTGCAGGAGTCGGGCCCAGGACTGGGGAAGCCTCCAGAGCTCAAAACCCCACTTGGTGACACAACTCACACATGCCCACGGTGCCCAGAGCCCAAATCTTGTGACACACCTCCCCCGTGCCCACGGTGCCCAGAGCCCAAATCTTGTGACACACCTCCCCCATGCCCACGGTGCCCAGAGCCCAAATCTTGTGACACACCTCCCCCGTGCCCNNNGTGCCCAGCACCTGAACTCTTGGGAGGACCGTCAGTCTTCCTCTTCCCCCCAAAACCCAAGGATACCCTTATGATTTCCCGGACCCCTGAGGTCACGTGCGTGGTGGTGGACGTGAGCCACGAAGACCCNNNNGTCCAGTTCAAGTGGTACGTGGACGGCGTGGAGGTGCATAATGCCAAGACAAAGCTGCGGGAGGAGCAGTACAACAGCACGTTCCGTGTGGTCAGCGTCCTCACCGTCCTGCACCAGGACTGGCTGAACGGCAAGGAGTACAAGTGCAAGGTCTCCAACAAAGCCCTCCCAGCCCCCATCGAGAAAACCATCTCCAAAGCCAAAGGACAGCCCGAGGAGATGACCAAGAACCAAGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTACCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAACTACAACACCACGCCTCCCATGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGACAAGAGCAGGTGGCAGCAGGGGAACATCTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCGCTACACGCAGAAGAGCCTCTCCCTGTCTCCGGGTAAATGA 1. keret leolvasása MKXLWFFLLLVAAPRWVLSQVHLQESGPGLGKPPELKTPLGDTTHTCPRCPEPKSCDTPPPCPRCPEPKSCDTPPPCPRCPEPKSCDTPPPCXXCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDXXVQFKWYVDGVEVHNAKTKLREEQYNSTFRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPEEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYNTTPPMLDSDGSFFLYSKLTVDKSRWQQGNIFSCSVMHEALHNRYTQKSLSLSPGK* Fejlődés megreked 2. Keret leolvasása (nincs fehérje) * 3. Keret leolvasása ETXVVLPSPGGSSQMGPVPGAPAGVGPRTGEASRAQNPTW*
Y A pre-B sejt receptor stimulálása sejtosztódást vált ki Nagy pre-B Large Pre-B Sok nagy pre-B sejt azonos pre-B sejt receptorokkal Proliferáció Large pre-B Y Éretlen B sejt L-lánc kifejeződés IgM a membránban IgM Osztódás leáll Pre-B receptor eltűnik Kis pre-B Intracellular VDJCH chain VL-JL átrendeződés
A pre-B sejt receptor stimulálásának következményei Gátolja a további H-lánc átrendeződést Könnyű lánc átrendeződés 3. Kiváltja a belépést a sejtciklusba 4. Túlélő szignál Nagy Pre-B Ismeretlen ligand 1. Biztosítja az egyedi specificitást Sztróma sejt ALLÉL KIZÁRÁS
AZ IMMUNOGLOBULIN GÉN SZEGMENSEK ÁTRENDEZŐDÉSÉNEK SORRENDJE D – J rekombináció V – DJ rekombináció VDJ – δ transzkripció δ transzláció Pót könnyű lánc V – J rekombináció VJ – (vagy VJ - ) transzkripció vagy transzláció B-sejt Szecernált IgM ellenanyag mIgD mIgM
A B-LIMFOCITÁK FEJLŐDÉSE A CSONTVELŐBEN Limfoid előalak Sajátot felismerő B-sejtek Sejtfelszíni molekulák ellen MHC fehérjék A hemopoetikus sejtekre jellemző közös receptorok apoptózis, klonális deléció Oldott molekulák ellen Háztartási gének anyagcsere termékek funkcionális válaszképtelenség anergia c-kit/CD44 Korai pro-B RAG-1/RAG-2 Késői pro-B H átrendeződés H átrendeződés Pót L Pre-B L átrendeződés B Szelekció klonális deléció Éretlen B B Egyéb specificitás B B PERIFERÁLIS NYIROKSZERVEK
PERIFÉRIÁS NYIROKSZERVEK Potenciális B-sejt készlet CSONTVELŐ Saját struktúra Saját felismerés Klonális deléció PERIFÉRIÁS NYIROKSZERVEK Hozzáférhető B-sejt készlet
A GÉNÁTRENDEZŐDÉS LEHETŐSÉGEI V-D-J 1.allél 2.allél mRNS Fehérje receptor Átrendeződés gátlása V-J V-J Nem sikeres Sikeres mRNS fehérje mRNS fehérje
A T SEJTEK ÉRÉSE ÉS DIFFERENCIÁLÓDÁSA A TÍMUSZBAN
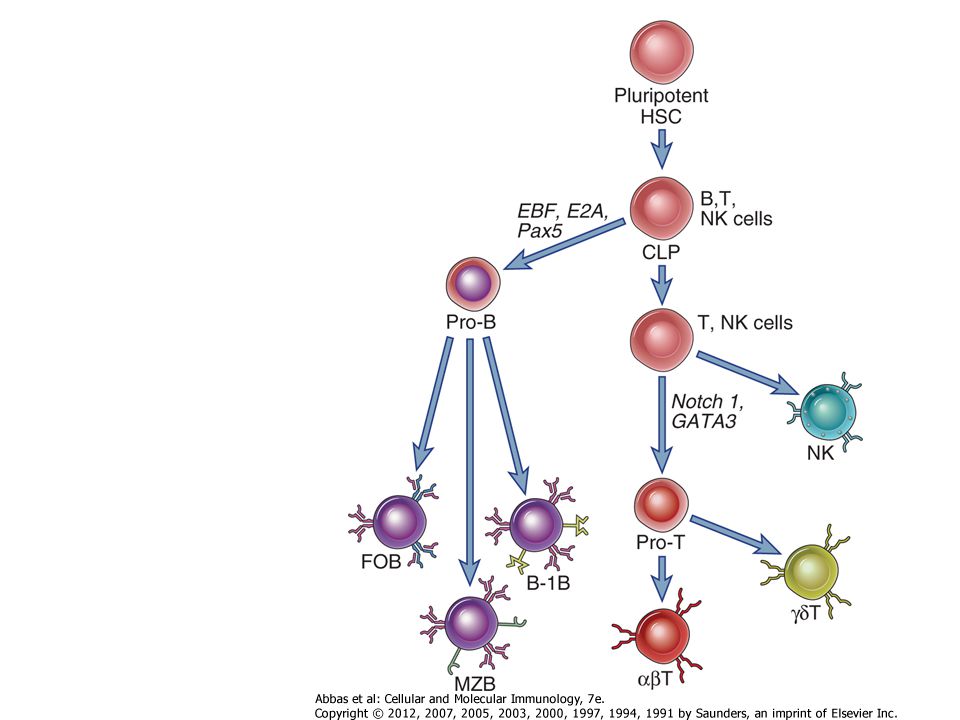
T- SEJT FEJLŐDÉS NK sejt Érett-B Pro-B Pre-B Pro-T Pre-T Érett-T Limfoid előalak NK sejt Érett-B c-kit/CD44 H átrendeződés Pót L L átrendeződés Szelekció klonális deléció B Pro-B Pre-B RAG-1/RAG-2 Pro-T Pre-T -átrendeződés Pre-T -átrendezés T Lymphoid progenitor: PU.1, IKAROS, C-kit + CD44 + kozos gamma lanc expresszió, ProT sejt: GATA3 expresszió, CD25 expresszió, IL7 fuggo osztodas, RAG expresszio Szelekció klonális deléció T Érett-T
A TÍMUSZBAN ZAJLÓ T SEJT ÉRÉSI FOLYAMATOK Az NK sejtek kialakulása – nincs TCR 2. A γδ és az αβ TCR kialakulása 3. Szelekciós folyamatok αβTCR – pozitív szelekció – negatív szelekció 4. A CD4+ és CD8+ T sejtek szétválása Korai pre-T Pót α-lánc Lck szignál β átrendezés IL-7 függő proliferáció Pro-T sikertelen β-lánc γδ T-sejt Nincs szelekció α átrendezés Késői pre-T CD4+CD8+ CD4+CD8+ Double positive αβ NKT-sejt sikertelen α-lánc nincs pozitív szelekció negatív szelekció αβCD4+ αβCD8+
A T LIMFOCITÁK SZELEKCIÓJA A TÍMUSZBAN Tok alatti terület Az elsődleges T sejt készlet zöme MHC-specifikus (V gének) 1-2% egy allotípusra A T sejt készlet fókuszálása a saját MHC felismerésre) A felesleges klónok eltávolítása Az önveszélyes klónok eltávolítása KÖZPONTI TOLERANCIA A T sejt készlet fókuszálása az idegen felismerésre Egyénre szabott optimalizált T sejt készlet a periférián A CD4 és a CD8 részt vesz a pozitív szelekcióban IL-7 függő osztódás Kéreg CD4-CD8- DN β+preTα TCRαβ CD4+CD8+ DP TCR- sMHC+sP sMHC+iP iMHC+iP szelekció Kéreg/velő határ nincs egerben 50 millio t-sejt eloalakbol -bol kb 1 millio T sejt fejlodik ki – szelekció Velő – AICD PERIFÉRIÁS TOLERANCIA AICD – aktiváció indukált apoptózis αβTCR αβTCR CD4+ CD8+
MHC-I + peptid komplexek CD8 toborzás a szinapszisba A KETTŐS POZITÍV (DP) T SEJTEK POZITÍV SZELEKCIÓJA A CD4 ÉS CD8 EGYSZERESEN POZITÍV (SP) SEJTEK FEJLŐDÉSÉT IS KIVÁLTJA CD4+CD8+ CD4+CD8+ Tímusz epitél sejt MHC-I + peptid komplexek CD8 toborzás a szinapszisba MHC-II + peptid komplexek CD4 toborzás a szinapszisba EGYMÁST KÖVETŐ α GÉN ÁTRENDEZŐDÉS POZITÍV SZELEKCIÓ 3 – 4 NAPIG BARE LYMPHOCYTE SYNDROME (BLS) MHC-I hiányában – nincs CD8+ sejt MHC-II hiányában – nincs CD4+ sejt
POZITÍV SZELEKCIÓ NEGATÍV SZELEKCIÓ Tímuszon belüli tanulási folyamat - nincs instrukció a specificitásra Kis aviditású saját MHC + saját peptid + TCR kölcsönhatás Tímusz epitél sejtek felszínén Saját peptidek összetétele, koncentrációja (idegen peptidek nincsenek jelen) Alacsony peptid dózis pozitív szelekciót eredményez – speciális ligand A DP T sejtek 80-90%-a hal meg a pozitív szelekcióban (2%/MHC) PASSZÍV SEJT HALÁL A NEGLEKCIÓ MIATT NEGATÍV SZELEKCIÓ Centrális tolerancia Nagy aviditású saját MHC + saját peptid + TCR kölcsönhatás Bármely tímuszon belüli antigén prezentáló sejt részvételével Epitél sejtek, csontvelői eredetű makrofágok és dendritikus sejtek Saját peptidek összetétele, koncentrációja Magas peptid dózis negatív szelekciót eredményez – speciális ligand Perifériás szövetek antigénjei? AKTÍV AKTIVÁCIÓ INDUKÁLTA SEJTHALÁL A saját MHC + idegen peptid felismerésére képes T-limfociták érésének előfeltétele a saját MHC + saját peptidekkel való gyenge kölcsönhatás SAJÁT MHC FELISMERÉSÉRE KÉPES DE A SAJÁTTAL SZEMBEN TOLERÁNS T-SEJT KÉSZLET KIALAKULÁSA A PERIFÉRIÁN FIZIOLÓGIÁS KÜSZÖB
A POZITÍV ÉS NEGATÍV SZELEKCIÓ EGYENSÚLYI SZEREPE A HOZZÁFÉRHETŐ T LIMFOCITA KÉSZLET KIALAKÍTÁSÁBAN Pozitív szelekció aránya Homozigóta szülők Heterozigóta gyerek Az MHC molekulák száma A negatív szelekció aránya nő az MHC számmal
Receptor editing
