Előadást letölteni
Az előadás letöltése folymat van. Kérjük, várjon

1
Ionösszetétel szalinitás
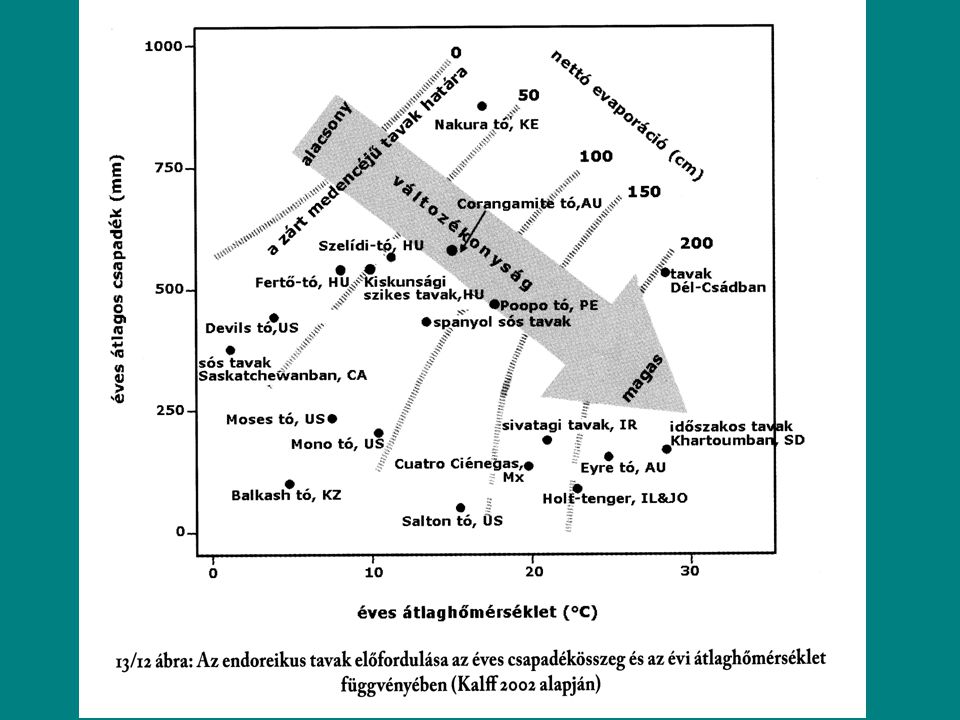
Vízben oldott szilárd anyagok Ionösszetétel szalinitás Szalinitás: Össziontartalom ezrelékben kifejezve. Becslése: Összes oldott anyag Elektromos vezetőképesség Forrás: Kőzettipus Gránit, bazalt dominanciájú vízgyűjtő: lágyvíz Mészkő dominanciájú vízgyűjtő: keményvíz Atmoszférikus precipitáció Savasodás (skandinávia, UK,Benelux államok, Csehország, Németo.) Párolgás-csapadék kapcsolat Nyílt tavak (exorheic): Zárt tavak (endorheic): szalinitásuk magas.
 Párolgás-csapadék kapcsolat. Nyílt tavak (exorheic): Zárt tavak (endorheic): szalinitásuk magas.")
2
Vízben oldott szilárd anyagok
vízgyűjtő geológiája – kemény kőzetről alacsony ion tartalmú víz folyik le Gránit gneisz vulkáni kőzet homokkő agyag mészkő

3
Legfontosabb anionok és kationok
Konzervatív ionok: Koncentrációjuk csak kissé változik az élővilág hatására Na, K, Mg, Cl Dinamikus ionok Az élőlények metabolizmusa nagymértékben hat a koncentrációjuk változására Ca, C, SO4, Legfontosabb ionok: Katnionok: Ca2+ > Mg2+ >Na+ > K+ Anionok: CO2- / HCO3+ > SO42+ > Cl-

4
Sós tavak Nincs kifolyás Párolgás meghaladja a hozzáfolyást
A hozzáfolyás nem biztosítja a stabil vízszintet Csoportosításuk hiposzalin : mg/L ≈ 3-20 ‰ ≈ S/cm mezoszalin : mg/L ≈ ‰ ≈ S/cm hiperszalin : > mg/L ≈ >50 ‰ ≈ > S/cm Na dominancia, de szikes, sziksós tavak (Ca, Mg dominancia) Cl dominancia Nyomelemek magas koncentrációja Osmotikus stessz Speciális élővilág: Artemia sp. (sórák), Halobacteria, Dunaliella, Spirulina Halak 140 ‰-ig élnek meg (fogaspontyok (Cyprinodon fajok) algák-, zooplankton fogyasztók főként a madarak (flamingók)
 Cl dominancia. Nyomelemek magas koncentrációja. Osmotikus stessz. Speciális élővilág: Artemia sp. (sórák), Halobacteria, Dunaliella, Spirulina. Halak 140 ‰-ig élnek meg (fogaspontyok (Cyprinodon fajok) algák-, zooplankton fogyasztók főként a madarak (flamingók)")
7
változás a víz ion-összetételében
Emberi hatás,savasodás Rajna ionösszetétele

8
Vas, mangán és néhány kis mennyiségben előforduló fém
Fe: A foszfor hozzáférhetőségét befolyásolja Fe2+ - vízben oldódik (FeS nem), míg az Fe3+ nem. Oxidatív, pH 7,5-7,7 környezetben Fe3+ - (Fe(OH)3) kicsapódik, a foszfor adszorbeálódik a vashidroxid felületén és kiülepszik (koprecipitáció). A toxikus nehézfémek is kicsapódnak a vas precipitátumokkal.
, míg az Fe3+ nem. Oxidatív, pH 7,5-7,7 környezetben Fe3+ - (Fe(OH)3) kicsapódik, a foszfor adszorbeálódik a vashidroxid felületén és kiülepszik (koprecipitáció). A toxikus nehézfémek is kicsapódnak a vas precipitátumokkal.")
9
Mn: hasonlóan viselkedik, mint a vas, de a mangánvegyületek oldhatósága jobb.

10
Ólom erősen toxikus. Ólomterhelés maximuma 1970-es évek, de már az ókori kultúrákban is kimutatható. Az üledékek ólomtartalma csökkenő tendenciát mutat. Higany Akut idegrendszeri problémát okoz (Minamoto-kór) Alumínium <4,5 pH oldékony. A magas alumínium koncentráció összefüggésbe hozható a Parkinson, Alzheimer kórral.
 Alumínium. <4,5 pH oldékony. A magas alumínium koncentráció összefüggésbe hozható a Parkinson, Alzheimer kórral.")
11
Szilícium A Föld második leggyakoribb eleme
A kovaalgák számára nélkülözhetelen. Eloszlásuk vízoszlopban a kovaalgák produkciójától függ. Partikulált szilícium Szilikát tartalmú ásványok kovaalgák

12
Kén- és nitrogénforgalom
Élő és élettelen formában felvehető Esszenciális aminosavak összetevője (cisztein, metionin) A fehérjeszerkezetben a szulfhidril (-SH) csoportok közötti diszulfid hidak stabilizálják a szerkezetet S-tartalmú szerves anyag lebomlása, az anaerob körülmények között szulfátredukció következtében megváltozhat a vízi környezet amely meghatározza: Más elemek körforgalmát (pl. Fe) A vízi ökoszisztémák produkcióját alapvetően az élőközösség jellegét De nincs kén limitáció a vizekben
 A fehérjeszerkezetben a szulfhidril (-SH) csoportok közötti diszulfid hidak stabilizálják a szerkezetet. S-tartalmú szerves anyag lebomlása, az anaerob körülmények között szulfátredukció következtében megváltozhat a vízi környezet amely meghatározza: Más elemek körforgalmát (pl. Fe) A vízi ökoszisztémák produkcióját. alapvetően az élőközösség jellegét. De nincs kén limitáció a vizekben.")
13
A kén eredete: Kőzet Légköri ülepedés
Szulfátok: szulfid vagy elemi kén tartalmú kőzetekből, talajokból víz jelenlétében kénsav formájában oldódnak ki Csökkenti pH-t így más ásvány oldódását befolyásolja Légköri ülepedés Vulkanikus gázok Biogén folyamatok Emberi tevékenység H2S kibocsátás → SO3 → SO3 → H2SO4 Kén tartalmú fosszilis tüzelőanyagok SO2 → H2SO4

14
A vízben oldott formában található kén döntő mennyiségét a szulfátok alkotják. A növények kénfelvétele alapvetően szulfát formájában történik, azonban a szerves anyag lebomlása során kénhidrogén keletkezik. Oxidatív körülmények között a H2S hamar oxidálódik szulfáttá. Elvileg lehetséges ugyan a H2S/SO42- egyensúly kiszámítása (feltéve, hogy a pH és a redoxviszonyokat ismerjük), de természetes vizekben ez nem vezet eredményre, tekintve, hogy a folyamatokat a kémiai átalakulásoknál gyorsabb mikrobiális anyagcsere folyamatok mediálják. Eutróf környezetben bakteriális tevékenység miatt, 100 mV redoxpotenciál alatt a H2S felhalmozódhat. Az erősen alkalikus vizekben e két forma mellett a H2S disszociációjából eredő HS- és S2- is előfordulhat. Alacsony redoxpotenciál és pH mellett a H2S részleges oxidációja kénkiválást (S0) eredményez.
, de természetes vizekben ez nem vezet eredményre, tekintve, hogy a folyamatokat a kémiai átalakulásoknál gyorsabb mikrobiális anyagcsere folyamatok mediálják. Eutróf környezetben bakteriális tevékenység miatt, 100 mV redoxpotenciál alatt a H2S felhalmozódhat. Az erősen alkalikus vizekben e két forma mellett a H2S disszociációjából eredő HS- és S2- is előfordulhat. Alacsony redoxpotenciál és pH mellett a H2S részleges oxidációja kénkiválást (S0) eredményez.")
15
Zalavári víz üledékkémiája

16
Főbb bakteriális anyagcsere típusok:
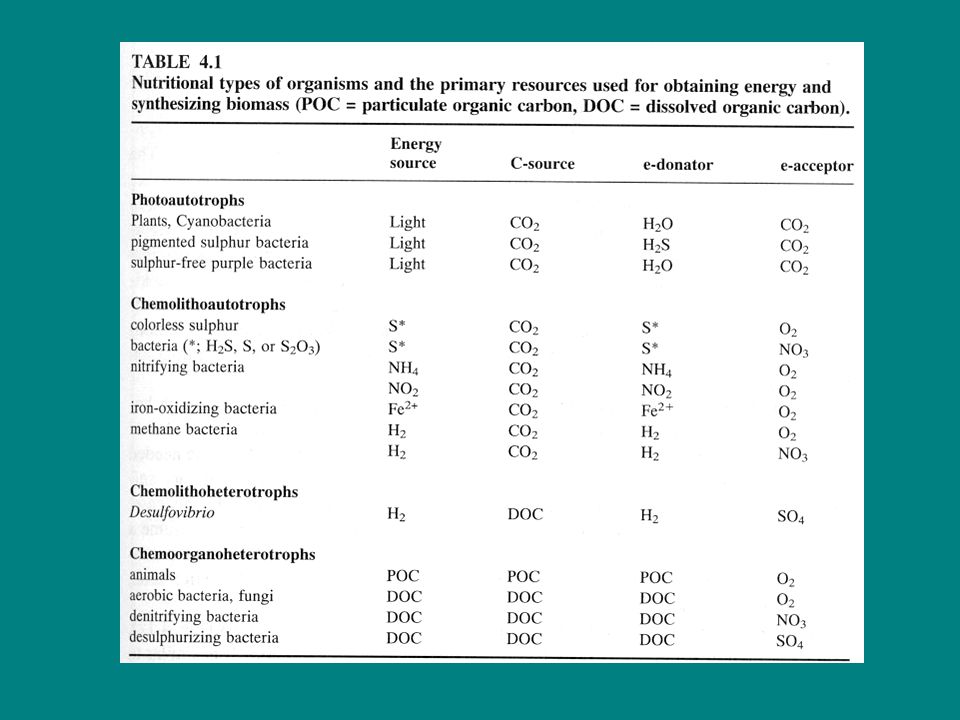
Ha egy baktérium anyagcsere típusát meg szeretnénk határozni, három kérdésre kell választ keresnünk: Energiaforrás: fény → fototróf, Kémiai → kemotróf Elektrondonor: szervetlen (S-, NH4, Fe2+, H2) → litotróf szerves → organotróf Szénforrás: szervetlen (CO2 vagy HCO3-) → autotróf szerves → heterotróf Ennek megfelelően egy olyan baktériumot, mely fotoszintetizál, tehát a fényt használja energiaforrásul (fototróf), s a fotoszintetikus elektrontranszportláncának elektrondonora szervetlen vegyület, pl. kénhidrogén (litotróf), továbbá a szerves anyagainak előállításához szervetlen szenet pl. széndioxidot használ (autotróf) e három sajátságot összefoglaló néven fotolitoautotrófnak nevezünk. Oxifototróf
 → litotróf. szerves → organotróf. Szénforrás: szervetlen (CO2 vagy HCO3-) → autotróf. szerves → heterotróf. Ennek megfelelően egy olyan baktériumot, mely fotoszintetizál, tehát a fényt használja energiaforrásul (fototróf), s a fotoszintetikus elektrontranszportláncának elektrondonora szervetlen vegyület, pl. kénhidrogén (litotróf), továbbá a szerves anyagainak előállításához szervetlen szenet pl. széndioxidot használ (autotróf) e három sajátságot összefoglaló néven fotolitoautotrófnak nevezünk. Oxifototróf.")
18
Baktériumoknak alapvető szerepe:
Szulfát redukció: Fehérjebontó baktériumok (heterotróf) fehérjék (-SH csoport) → HS- és H2S Pseudomonas, Chromobacter Bacterium Szulfátredukáló baktériumok SO42-, SO32- és S → HS- és H2S Heterotróf vagy autotróf -SO4 oxigénjét használják a szervesanyag oxidációjához (szulfát légzés) Desulfovibrio, Desulfotomaculum (anaerobikus)
 fehérjék (-SH csoport) → HS- és H2S. Pseudomonas, Chromobacter Bacterium. Szulfátredukáló baktériumok. SO42-, SO32- és S → HS- és H2S. Heterotróf vagy autotróf. -SO4 oxigénjét használják a szervesanyag oxidációjához (szulfát légzés) Desulfovibrio, Desulfotomaculum (anaerobikus)")
19
Szulfát oxidáció: Fotoszintetikus (színes) kénbaktériumok
H2S-t fotoszintetikus elektrontranszportláncuk elektrondonorául használják. Fotolitoautotrófok, anaerobok Zöld kénbaktériumok (Chlorobacteriaceae): Chlorobium, Pelodyction A zöld kénbaktériumok az oxidációs terméket (elemi kéntől a szulfátig) leadják a vízbe, bíbor baktériumok (Thiorhodaceae): Chromatium, Rhodothece A bíbor-kénbaktériumok pedig nem (ez esetben kén a végtermék, mert a további oxidáció ozmotikusan aktív ionok keletkezéséhez vezetne, s ennek következményei a sejt számára végzetesek: kipukkad). Mély tavakban e baktériumok a rétegzett periódus alatt fordulnak elő, mégpedig esetenként nagy sűrűségben a metalimnionban. Ide még lejut a fény, de az anoxikus hipolimnionból a szükséges redukált kénvegyülethez (H2S) is hozzájutnak.
: Chlorobium, Pelodyction. A zöld kénbaktériumok az oxidációs terméket (elemi kéntől a szulfátig) leadják a vízbe, bíbor baktériumok (Thiorhodaceae): Chromatium, Rhodothece. A bíbor-kénbaktériumok pedig nem (ez esetben kén a végtermék, mert a további oxidáció ozmotikusan aktív ionok keletkezéséhez vezetne, s ennek következményei a sejt számára végzetesek: kipukkad). Mély tavakban e baktériumok a rétegzett periódus alatt fordulnak elő, mégpedig esetenként nagy sűrűségben a metalimnionban. Ide még lejut a fény, de az anoxikus hipolimnionból a szükséges redukált kénvegyülethez (H2S) is hozzájutnak.")
20
színtelen kénbaktériumok
kemolitoautotrófok (ezért is színtelenek: nem színezik őket fotoszintetikus pigmentek) s a szulfitot, kenet, tioszulfátot oxidálják: H2S + ½ O2 → S + H2O S + H2O + 3/2 O2 → SO H+ S2O32- + H2O + 2O2 → 2SO H+ Thiothrix, Beggiatoa, Thiobacillus
 s a szulfitot, kenet, tioszulfátot oxidálják: H2S + ½ O2 → S + H2O. S + H2O + 3/2 O2 → SO H+ S2O32- + H2O + 2O2 → 2SO H+ Thiothrix, Beggiatoa, Thiobacillus.")
21
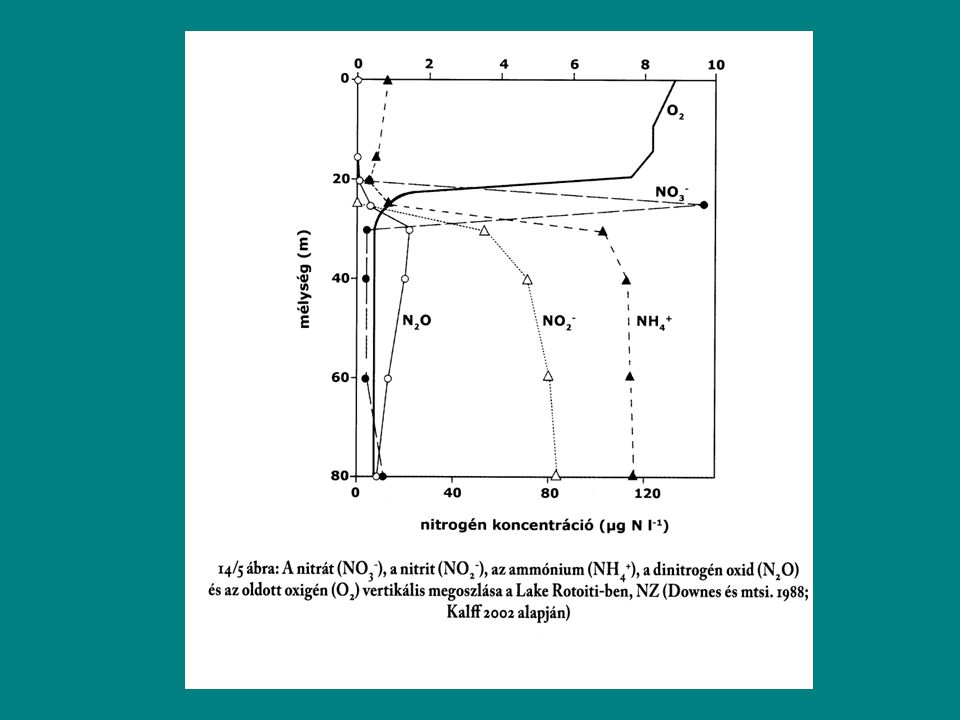
Nitrogénforgalom Központi szerep a vizek anyagforgalmában
Nagy mennyiségben igénylik a szervezetek Primerprodukciót meghatározza N-limitáció N-formák: Elemi N2 Szerves-N Ammónia (NH3), ammónium-ion (NH4+) Nitrit (NO2-) Nitrát (NO3-)
, ammónium-ion (NH4+) Nitrit (NO2-) Nitrát (NO3-)")
22
N-forrás N-veszteség Csapadék Nitrogén fixáció
Befolyó vizek (felszíni- és talajvizek) N-veszteség Elfolyás Denitrifikáció kiülepedés az üledékbe Szennyezéstől távoli helyen 2% per év növekedés
 N-veszteség. Elfolyás. Denitrifikáció. kiülepedés az üledékbe. Szennyezéstől távoli helyen 2% per év növekedés.")
23
Nitrogén fixáció: Cianobaktériumok (Kékalgák) Baktériumok
N2 → NH4+ → NO2-, NO3- Cianobaktériumok (Kékalgák) Heterociszta Fény és ATP igényes (energia) Aphanizomenon, Anabaena, Nostoc Baktériumok Heterotróf N2-kötők Azotobakter, Clostridium pasteurianum Fotoszintetizálók Közel az összes fotoszintetizáló baktérium képes N2-kötésre Mikorrhiza wetland-eken Actinomyceta mycorrhiza gombák Alnus, Myrica (mírtusz) gyökerében
 Heterociszta. Fény és ATP igényes (energia) Aphanizomenon, Anabaena, Nostoc. Baktériumok. Heterotróf N2-kötők. Azotobakter, Clostridium pasteurianum. Fotoszintetizálók. Közel az összes fotoszintetizáló baktérium képes N2-kötésre. Mikorrhiza wetland-eken. Actinomyceta mycorrhiza gombák Alnus, Myrica (mírtusz) gyökerében.")
24
N-formák és N-transzorfációk Ammonia (NH4+/NH4OH)
Az ammónia (NH3) a vízben lúgként viselkedik, mivel protonfelvételre képes: NH3 + H2O = OH- + NH4+ NH4+ + H2O = H3O + NH3 Forrás: heterotróf baktérium általi lebontó folyamatok végterméke Vízi szervezetek exkrétuma NH4+/NH4OH pH függő Az ammónia a vízben ammónium-ionként (NH4+), ill. disszociálatlan ammónium-hidroxidként (NH4OH) fordul elő. ammónium-hidroxidként (NH4OH) erősen toxikus 8-as pH alatt ammónium ion (NH4+) 10,5-nél magasabb pH értékeken jellemző az NH4OH forma magas pH-n az ammónia (NH3) 3000:1 300:1 30:1 1:1 pH 6 pH 7 pH 8 pH 9,5 NH4+/NH4OH
 a vízben lúgként viselkedik, mivel protonfelvételre képes: NH3 + H2O = OH- + NH4+ NH4+ + H2O = H3O + NH3. Forrás: heterotróf baktérium általi lebontó folyamatok végterméke. Vízi szervezetek exkrétuma. NH4+/NH4OH pH függő. Az ammónia a vízben ammónium-ionként (NH4+), ill. disszociálatlan ammónium-hidroxidként (NH4OH) fordul elő. ammónium-hidroxidként (NH4OH) erősen toxikus. 8-as pH alatt ammónium ion (NH4+) 10,5-nél magasabb pH értékeken jellemző az NH4OH forma. magas pH-n az ammónia (NH3) 3000:1. 300:1. 30:1. 1:1. pH 6. pH 7. pH 8. pH 9,5. NH4+/NH4OH.")
25
Az ammóniumot számos alga- és magasabbrendű növényfaj tudja közvetlen nitrogénforrásul felhasználni, egyúttal ez a „legolcsóbb” szervetlen N-forrás, tekintve, hogy az aminokba (H2N— ) való beépüléshez nem kell sejten belüli (energiaigényes!) oxidációs-szám változásokon keresztülmennie. Heterotrófok termelik Eloszlása: Oligtróf tóban alacsony konc. kis fokú dekompozíció miatt Trofogén zónában konc. szintén alacsony az algák asszimilációja miatt Ha a hipolimnium szerves anyagban gazdag, akkor az NH4+-N accumulálódik, anaerob környezetben fokozódik bakteriális nitrifikáció miatt A szerves nitrogénvegyületek (aminosavak, urea, karbamid, metilamin, stb.) forrása a szerves anyagok bomlása. Lehet oldott vagy alakos állapotban, akár az elhalt, szervezetek törmelékeiben akár az élő planktonban. A nitrit (NO2-) átmeneti redox állapota miatt általában igen kis mennyiségben található meg vizeinkben.
 forrása a szerves anyagok bomlása. Lehet oldott vagy alakos állapotban, akár az elhalt, szervezetek törmelékeiben akár az élő planktonban. A nitrit (NO2-) átmeneti redox állapota miatt általában igen kis mennyiségben található meg vizeinkben.")
26
A N oxidációs száma a N-körforgalomban szerepet játszó főbb folyamatokban

27
Nitrogénforgalom

28
Oldott (DON)és partikulált szerves nitrogén (PON)
Nitrifikáció NH4+ → NO2- → NO3- O2 igényes Energiatermelő folyamat -> C-asszimiláció Nitrosomonas: NH4+ → NO2- pH közel neutrális, tág hómérsékleti tartomány Néhány metán-oxidáló baktérium is képes. Nitrobacter: NO2- → NO3- Nem toleráns alacsony hőm.-el és magas pH-val szemben Oldott (DON)és partikulált szerves nitrogén (PON) DON: akár 50 %-a az összesN-nek Aminosavak, polipeptidek PON: Plankton, szeszton A PON 5-10 x nagyobb mennyiségben található, de az eutrofizációval az arány közelít az 1:1 a trofolitikus zónában. A vízi növények, algák és cianobaktériumok választanak ki N-gazdag szerves anyagokat, amik a bakteriális hurok tápanyagforrása. Bakteriális hurok: Heterotróf baktériumok -> heterotróf flagellata/ciliata Gazdag litorális zónájú tavak DON forrása a makrofita vegetáció. DON forrás: edényes növények lebomlása
és partikulált szerves nitrogén (PON) DON: akár 50 %-a az összesN-nek. Aminosavak, polipeptidek. PON: Plankton, szeszton. A PON 5-10 x nagyobb mennyiségben található, de az eutrofizációval az arány közelít az 1:1 a trofolitikus zónában. A vízi növények, algák és cianobaktériumok választanak ki N-gazdag szerves anyagokat, amik a bakteriális hurok tápanyagforrása. Bakteriális hurok: Heterotróf baktériumok -> heterotróf flagellata/ciliata. Gazdag litorális zónájú tavak DON forrása a makrofita vegetáció. DON forrás: edényes növények lebomlása.")
29
Denitrifikáció NO3- → NO2- → NH4+ → N2 Nitrátredukcó:
Nitrát-asszimiláció Algák, magasabb rendű növények Bakteriális denitrifikáció Pseudomonas, Escherichia, Bacillus, Micrococcus Achromobacter Nitrátlégzés: a nitrát, nitrit az elektronacceptor

31
Nitrogénforgalom

32
Nitrogénforgalom A földi környezet alapvető jellemvonása, hogy a N legnagyobb mennyisége a légkörben található N2 gáz formájában. Ennek megkötésére, vagyis biológiailag hozzáférhetővé tételére csak néhány szevezetcsoport képes. A folyamatot N2-kötésnek nevezzük. A fent tárgyalt, s az összességében az élőlények számára rendelkezésre álló N mennyiség csökkenését eredményező folyamatok könnyen rövid idő alatt N-hiányt eredményeznének, ha azokat nem ellensúlyozná a légköri N2 megkötésének folyamata. A légköri N2 megkötésére számos aerob és anaerob baktérium (sok közülük szimbionta) és más prokarióta szervezet képes. Utóbbiak közt a vizekben kiemelkednek a cianobaktériumok (kékalgák). Ezek egyik csoportja, a Nostocales rendbe tartozó fajok (pl. Anabaena, Aphanizomenon, Cylindrispermopsis, Gloeotrichia, Nostoc) a legjelentősebbek, bár anaerob körülmények közti N2 kötést más rendben (Chroococcales; pl. Microcystis) is kimutattak. A N2-kötést az un. nitrogenáz enzimkomplex végzi. Ennek két alegysége közül az egyik molibdént és vasat is tartalmaz, a másik csak vasat (emiatt molibdénszegény vízben gátolt lehet a N2-kötés). Az enzimkomplex működését az oxigén erősen gátolja, ezért a folyamat csak anaerob körülmények közt mehetne végbe, ha nem létezne több olyan biológiai evolúciós alkalmazkodási mechanizmus, mely mégiscsak lehetővé teszi az aerob körülmények közti N2-kötést. Ezek közül a három, egyidejűleg működő legfontosabb a következő: 1) specializált, vastag falú sejtek; 2) bakteriális fotoszintézis és 3) szimbionta baktériumok jelenléte.
 és más prokarióta szervezet képes. Utóbbiak közt a vizekben kiemelkednek a cianobaktériumok (kékalgák). Ezek egyik csoportja, a Nostocales rendbe tartozó fajok (pl. Anabaena, Aphanizomenon, Cylindrispermopsis, Gloeotrichia, Nostoc) a legjelentősebbek, bár anaerob körülmények közti N2 kötést más rendben (Chroococcales; pl. Microcystis) is kimutattak. A N2-kötést az un. nitrogenáz enzimkomplex végzi. Ennek két alegysége közül az egyik molibdént és vasat is tartalmaz, a másik csak vasat (emiatt molibdénszegény vízben gátolt lehet a N2-kötés). Az enzimkomplex működését az oxigén erősen gátolja, ezért a folyamat csak anaerob körülmények közt mehetne végbe, ha nem létezne több olyan biológiai evolúciós alkalmazkodási mechanizmus, mely mégiscsak lehetővé teszi az aerob körülmények közti N2-kötést. Ezek közül a három, egyidejűleg működő legfontosabb a következő: 1) specializált, vastag falú sejtek; 2) bakteriális fotoszintézis és. 3) szimbionta baktériumok jelenléte.")
33
Nitrogénforgalom Heterocita fénymikroszkópos felvételen
A heterocitát körülvevő aerob szimbionta baktériumok. Gátolják az oxigén átdiffundálását a vékonyabb sejtfalon.

34
Nitrogénforgalom A N2-kötés energiaigényes folyamat, minthogy egyetlen N2 megkötéséhez mintegy 12 ATP-re van szükség: N2 + (kb.) 12 ATP + 6e- + 6H+ → 2 NH3 + (kb.) 12 ADP + (kb.) 12P. Ezért is nem engedheti meg magának a sejt, hogy a fotoszintézisről, mint ATP termelő folyamatról lemondjon, csak azt, hogy annak O2 termeléssel járó formájáról tér át egy olyanra, mely a N2-kötéssel összeegyeztethető. Az energiaigényesség további 2 következménnyel jár: 1) N2-kötés csak napközben zajlik (az energiát termelő fotoszintézis miatt), s mélységi megoszlásának intenzitása arányos a fotoszintézisével (14/9. ábra), valamint 2) mert a N2-kötésre fordított energia elvész a populáció növekedése számára a cianobaktériumok csak akkor kezdenek N2-t kötni, ha a környezet NH4+ tartalma kimerült, ill., ha már folyik N2 kötés, akkor az lassul majd megáll, ha pótlólag NH4+ kerül a vízbe.
 12 ATP + 6e- + 6H+ → 2 NH3 + (kb.) 12 ADP + (kb.) 12P. Ezért is nem engedheti meg magának a sejt, hogy a fotoszintézisről, mint ATP termelő folyamatról lemondjon, csak azt, hogy annak O2 termeléssel járó formájáról tér át egy olyanra, mely a N2-kötéssel összeegyeztethető. Az energiaigényesség további 2 következménnyel jár: 1) N2-kötés csak napközben zajlik (az energiát termelő fotoszintézis miatt), s mélységi megoszlásának intenzitása arányos a fotoszintézisével (14/9. ábra), valamint. 2) mert a N2-kötésre fordított energia elvész a populáció növekedése számára a cianobaktériumok csak akkor kezdenek N2-t kötni, ha a környezet NH4+ tartalma kimerült, ill., ha már folyik N2 kötés, akkor az lassul majd megáll, ha pótlólag NH4+ kerül a vízbe.")
35
A N2-kötés napszakos ritmusa (b) és intenzitása különböző vízmélységekben
 és intenzitása különböző vízmélységekben")
36
Nitrogénforgalom A N2-kötést leginkább a nyíltvízben vizsgálták, de a rendelkezésre álló kevés adat arra utal, hogy a parti régió szerepe (pl. a bevonatban élő heterocitás cianobaktériumok tevékenysége miatt) egyáltalán nem elhanyagolható. Az e területeken folyó denitrifikáció az ökoszisztéma N készletét csökkenti, ezáltal a N/P arányt is csökkenti, s ez stimulálja a N2- kötést. A kiterjedt litorális régióval rendelkező sekély tavakban lehet ennek igan nagy jelentősége.
 egyáltalán nem elhanyagolható. Az e területeken folyó denitrifikáció az ökoszisztéma N készletét csökkenti, ezáltal a N/P arányt is csökkenti, s ez stimulálja a N2- kötést. A kiterjedt litorális régióval rendelkező sekély tavakban lehet ennek igan nagy jelentősége.")
37
Foszforforgalom és eutrofizálódás
Biológiai makroelem -> organizmusok nagy mennyiségben veszik fel Mennyisége a környezetben lényegesen alacsonyabb -> a vizek produktivitását a hozzáférhető P limitálja

38
P - formák Ortofoszfát (PO43-) Szerves foszfát Összes foszfor (TP)
Legfontosabb inorganikus forma Közvetlenül hozzáférhető Mennyisége nagyon alacsony lehet -> gyor asszimiláció miatt Szerves foszfát Édesvizekben a legnagyobb mennyiségben megtalálható P forma Összes foszfor (TP) A P terhelés mérőszáma Szűretlen vízből: oldott P és partikulált P
 A P terhelés mérőszáma. Szűretlen vízből: oldott P és partikulált P.")
39
Partikulált foszfor Élőlényekben található foszfor Talajásványokból
DNS, RNS, foszfortartalmú fehérjék, alacsony molekulatömegű észterek vitaminok Nukleotid foszfátok (ADP, ATP) Légzésben, fotoszintézisben C-forgalmi utakban energia transzfer Talajásványokból Hidroxiapatit P abszorbens ásványok Agyagásványok, vashidroxidok, karbonátok Elhalt szervesanyagban, vagy azon abszorbeálva
 Légzésben, fotoszintézisben C-forgalmi utakban energia transzfer. Talajásványokból. Hidroxiapatit. P abszorbens ásványok. Agyagásványok, vashidroxidok, karbonátok. Elhalt szervesanyagban, vagy azon abszorbeálva.")
40
Oldott P Ortofoszfát (PO43-)
Polifoszfátok (szintetikus detergensekből) Szerves kolloidok Alacsony molekulatömegű foszfátészterek >100 Hipertróf 30-100 Eutróf 10-30 Meso-eutróf 5-10 Oligo-mesotróf <5 Ultra-oligotróf TP (g/l) Produktivitás
 Szerves kolloidok. Alacsony molekulatömegű foszfátészterek. >100. Hipertróf Eutróf Meso-eutróf Oligo-mesotróf. <5. Ultra-oligotróf. TP (g/l) Produktivitás.")
41
P forrás Ülepedés Talajvíz Felszíni lefolyás folyóvizek
Kevésbé jelentős, mint N esetében Elsősorban emberi tevékenységből ered Lakatlan régiókban a P ülepedés : < 3 μg P / l Városi-ipari régiókban : >100 μg P / l Talajvíz P-tartalma alacsony, átlagos konc. ~ 20 μg P / l Kőzetekből való kioldódás csekély Felszíni lefolyás folyóvizek Talaj A felszíni vizek kémiai összetétele függ a talajtipusoktól. Felszíni lefolyás a legfőbb P forrás Emberi tevékenység Detergensek (mosószerek) Urbanizáció (szennyvizek) Mezőgazdaság (műtrágyák) Ipar (élelmiszeripar)
 Urbanizáció (szennyvizek) Mezőgazdaság (műtrágyák) Ipar (élelmiszeripar)")
42
Belső terhelés Külső terhelés Alacsony redoxpotenciál Befolyók légkör
P felszabadulás Külső terhelés Befolyók légkör

43
Vertikális profil Vertikális eloszlása a trofitástól függ
Eutróf tavak hipolimnionjában élő és holt szervesanyag (plankton eső) kalcit koprecipitáció, bakteriális produkció
 kalcit koprecipitáció, bakteriális produkció.")
44
Eutrofizáció Eutrofizálódás: vizek növényi (N, P) tápanyagokban való gazdagodása Eredetileg (Neumann XX. Sz eleje: oligotróf: kevés fitoplankton; eutróf: sok fitoplankton) trofitás limitálás: súly arány: 1 P : 7 N : 40 C szárazsúly gramm Redfield arány (atom): 1 P : 16 N : 106 C típusai Planktonikus eutrofizáció Bentonikus eutrofizáció
 trofitás limitálás: súly arány: 1 P : 7 N : 40 C szárazsúly gramm. Redfield arány (atom): 1 P : 16 N : 106 C. típusai. Planktonikus eutrofizáció. Bentonikus eutrofizáció.")
45
Oligotrofizáció eszközei:
Szennyvíztisztítók P-leválasztása A fő terhelést jelentő folyóviz wetlandeken át való vezetése Művelt területek szegélyezése dús vegetációval Talaj erózió, bemosódás csökkentése Üledék foszfortartalmának immobilizálása Al2(SO3) és FeCl3 bevitelével Meszezés Kotrás Balatonnál nem vált be Biomanipuláció Hipolimnium levegőztetése Fény meghatározta szint P meghatározta szint
 és FeCl3 bevitelével. Meszezés. Kotrás. Balatonnál nem vált be. Biomanipuláció. Hipolimnium levegőztetése. Fény meghatározta szint. P meghatározta szint.")
46
Foszforforgalom A limnológiában egyetlen más elemet sem tanulmányoztak olyan részletesen, mint a foszfort. Ennek az az oka, hogy egyike a biológiai makroelemeknek, az élőlények anyagcseréje nagy mennyiséget igényel (nukleinsavak, ATP/ADP). Ugyanakkor mennyisége a a környezetben lényegesen alacsonyabb, mint a többi biológiai makroelemé. Ezért a vizek produktivitását igen gyakran a hozzáférhető P mennyisége limitálja. Az előző fejezetekben láttuk, hogy a többi biológiai makroelem – s főleg a S és a N – igen sokféle oxidációs állapotban fordul elő a vízben, s átalakulásaikat mikrobiális folyamatok mediálják. A P különbözik: domináns szervetlen formája a foszfát (PO43-) s ugyanilyen oxidációs számmal fordul elő az élőlényekben szervesen kötött foszfor formájában. A szerves frakció tekintélyes: becslések szerint a vizek P készletének 90% fordul elő a szerves frakcióban, mely áll maguknak az élő szervezeteknek a P taralmából, s ehhez számítjuk az elhalt szerves alkotókon adszorbeálódott, vagy bennük kötött foszfort. Bár kémiai szempontból egyéb oxidációs számú P-vegyületek is előfordulhatnak, ezek jelentősége a vízi anyagforgalomban tán a foszfin PH3↑ kivételével (15/1 keret) elhanyagolható. A P a vizekbe a nagyrészt a vízgyűjtőn át, kisebb részben a légköri ülepedéssel jut be. A litoszférában elsősorban az apatitokból (Ca5[PO4]3-) oldódik ki, melyeknek hidroxóniummal (hidroxiapatit), kloriddal és fluoriddal asszociált változatai ismertek, de legalább 205 egyéb, kis mennyiségben előforduló ismert ásvány is tartalmaz P-t (Fischer, 1973). A vulkáni kőzetekből való kioldódását a szénsav segíti. A biológiai úton történő P felhalmozódás jelentős guánó-telepek kialakulásához vezetett.
. Ugyanakkor mennyisége a a környezetben lényegesen alacsonyabb, mint a többi biológiai makroelemé. Ezért a vizek produktivitását igen gyakran a hozzáférhető P mennyisége limitálja. Az előző fejezetekben láttuk, hogy a többi biológiai makroelem – s főleg a S és a N – igen sokféle oxidációs állapotban fordul elő a vízben, s átalakulásaikat mikrobiális folyamatok mediálják. A P különbözik: domináns szervetlen formája a foszfát (PO43-) s ugyanilyen oxidációs számmal fordul elő az élőlényekben szervesen kötött foszfor formájában. A szerves frakció tekintélyes: becslések szerint a vizek P készletének 90% fordul elő a szerves frakcióban, mely áll maguknak az élő szervezeteknek a P taralmából, s ehhez számítjuk az elhalt szerves alkotókon adszorbeálódott, vagy bennük kötött foszfort. Bár kémiai szempontból egyéb oxidációs számú P-vegyületek is előfordulhatnak, ezek jelentősége a vízi anyagforgalomban tán a foszfin PH3↑ kivételével (15/1 keret) elhanyagolható. A P a vizekbe a nagyrészt a vízgyűjtőn át, kisebb részben a légköri ülepedéssel jut be. A litoszférában elsősorban az apatitokból (Ca5[PO4]3-) oldódik ki, melyeknek hidroxóniummal (hidroxiapatit), kloriddal és fluoriddal asszociált változatai ismertek, de legalább 205 egyéb, kis mennyiségben előforduló ismert ásvány is tartalmaz P-t (Fischer, 1973). A vulkáni kőzetekből való kioldódását a szénsav segíti. A biológiai úton történő P felhalmozódás jelentős guánó-telepek kialakulásához vezetett.")
47
Foszforforgalom A víz összes foszfor (TP, total P) tartalma oldott (DP, dissolved P) és partikulált (PP, particulate P) frakcióból áll. A partikulált frakciót a következőképp bonthatjuk tovább: az élőlényekben található P, ami áll relatíve stabil nukleinsavakból (DNS, RNS) és olyan foszfortartalmú fehérjékből, melyek foszfort tartalmaznak, de nem vesznek közvetlenül részt a sejt belső foszforszállítási folyamataiban alacsony molekulasúlyú enzim-észeterből, vitaminokból, stb. és a légzésben, a fotoszintézisben és a CO2 asszimliációban alapvető szerepet játszó nukleotid foszfátokból (ATP, ADP) a talajokból bekerülő ásványi formák, pl. hidroxiapatit, s minden egyéb ásványi anyag, melyhez a P adszorbeálódik (agyagásványok, vashidroxidok, karbonátok; az elhalt szerves anyagban található, vagy azon adszorbeálódott P. Az oldott frakció legfontosabb összetevői az ortofoszfát (PO43-), polifoszfátok, amik gyakran szintetikus detergensekből kerülnek a vízbe, szerves kolloidok és alacsony molekulatömegű foszfátészterek.
 és olyan foszfortartalmú fehérjékből, melyek foszfort tartalmaznak, de nem vesznek közvetlenül részt a sejt belső foszforszállítási folyamataiban. alacsony molekulasúlyú enzim-észeterből, vitaminokból, stb. és. a légzésben, a fotoszintézisben és a CO2 asszimliációban alapvető szerepet játszó nukleotid foszfátokból (ATP, ADP) a talajokból bekerülő ásványi formák, pl. hidroxiapatit, s minden egyéb ásványi anyag, melyhez a P adszorbeálódik (agyagásványok, vashidroxidok, karbonátok; az elhalt szerves anyagban található, vagy azon adszorbeálódott P. Az oldott frakció legfontosabb összetevői az. ortofoszfát (PO43-), polifoszfátok, amik gyakran szintetikus detergensekből kerülnek a vízbe, szerves kolloidok és. alacsony molekulatömegű foszfátészterek.")
48
A foszforforgalom klasszikus modellje: a P-kérdést vízkémiai kérdésnek gondolja
Megérthetjük az előzőek alapján: A tavak foszforforgalmának klasszikus modelljét jórészt a II. világháború idején dolgozta ki Ohle, Eisener és Mortimer (Kalff 2002). Eszerint a tavakba kerülő PO43- aerob körülmények közt igen gyorsan vas-oxi-hidroxid (FeOOH) flokkulumokhoz adszorbeálódik kiválik, s leülepszik. Az aerob üledékfelszínen ezen aggregátumok hatékony csapdát képeznek az anaerob üledékrétegekből származó P számára, ezzel megakadályozzák a víztérbe jutását. Mortimer (1941) 200 mV redoxpotenciálban állapította meg azt a határt, ami P szempontjából kritikus: e felett annak csapadékos (elsősorban Fe[III]-mal asszociált), oldhatatlan formái jellemzőek, s alatta a vas Fe(II) formájú, mely mellett a PO43- oldatban maradhat. Továbbfejlesztés: Az alapvetően Fe(II)-Fe(III)-P rendszert összekötötték a kénciklussal, tekintve, hogy a Fe2+ a szulfátredukció során keletkező szulfiddal oldhatatlan FeS csapadékot képez, ezért alacsony vastartalom esetén nem marad vas a P becsapdázására. Azt a folyamatot, mely a víz-üledék határ redoxpotenciál változásai miatti P-felszabadulást okozza belső foszforterhelésnek nevezzük (ebbe ma beleértjük a belső seice okozta P tartalom növekedést az eufotikus rétegben), megkülönböztetve a külső forszforterheléstől, mely a vizeket befolyóikon vagy a légkörön át éri. Az üledék redoxi állapota és belső foszforterhelés folyamata azért jelentős, mert jellemző módon az üledék intersticiális vizének P-tartalma 5-20x-osan haladja meg a közvetlenül felette lévő vízrétegét, s nem mindegy, hogy ez a hatalmas P készlet az általában P-limitált eufotikus rétegbe juthat vagy sem.
. Eszerint a tavakba kerülő PO43- aerob körülmények közt igen gyorsan vas-oxi-hidroxid (FeOOH) flokkulumokhoz adszorbeálódik kiválik, s leülepszik. Az aerob üledékfelszínen ezen aggregátumok hatékony csapdát képeznek az anaerob üledékrétegekből származó P számára, ezzel megakadályozzák a víztérbe jutását. Mortimer (1941) 200 mV redoxpotenciálban állapította meg azt a határt, ami P szempontjából kritikus: e felett annak csapadékos (elsősorban Fe[III]-mal asszociált), oldhatatlan formái jellemzőek, s alatta a vas Fe(II) formájú, mely mellett a PO43- oldatban maradhat. Továbbfejlesztés: Az alapvetően Fe(II)-Fe(III)-P rendszert összekötötték a kénciklussal, tekintve, hogy a Fe2+ a szulfátredukció során keletkező szulfiddal oldhatatlan FeS csapadékot képez, ezért alacsony vastartalom esetén nem marad vas a P becsapdázására. Azt a folyamatot, mely a víz-üledék határ redoxpotenciál változásai miatti P-felszabadulást okozza belső foszforterhelésnek nevezzük (ebbe ma beleértjük a belső seice okozta P tartalom növekedést az eufotikus rétegben), megkülönböztetve a külső forszforterheléstől, mely a vizeket befolyóikon vagy a légkörön át éri. Az üledék redoxi állapota és belső foszforterhelés folyamata azért jelentős, mert jellemző módon az üledék intersticiális vizének P-tartalma 5-20x-osan haladja meg a közvetlenül felette lévő vízrétegét, s nem mindegy, hogy ez a hatalmas P készlet az általában P-limitált eufotikus rétegbe juthat vagy sem.")
49
A foszforforgalom modern modellje: a P-kérdést biológiai kérdésnek gondolja
A 20. század utolsó évtizedeiben végzett kutatások mutattak rá arra, hogy a P kibocsátást inkább biológiai, mint kémiai folyamatok határozzák meg. Az aerob, heterotróf baktériumok a szerves anyagok lebontása során keletkező oldható reaktív foszfort (SRP = soluble reacive phosphorus) közvetlenül a vízbe adják le. A Fe(III) redukciójához az oldott oxigén hiánya nem elégséges feltétel, mert az ezt tartalmazó flokkulumokat a hozzájuk asszociált szerves anyag stabilizálja. Ezért a vasoxidáló baktériumok (ezek a Fe[III]-at a szerves anyag oxidációja során keletkező elektronok akceptorának használják) jelenléte elengedhetetlen. Ha a klasszikus modell igaz lenne, akkor az üledék Fe2+ és PO43- kibocsátásának egyidejűen kéne bekövetkeznie. Ezzel szemben a kettő közt fáziseltolódás tapasztalható, s az a tény, hogy az antibiotikumokkal kezelt üledék P-kibocsátása lényegesen kisebb, mint a kezeletlené szintén a mikrobiális folyamatok fontosságára utal (Gächter és mtsi. 1988), sőt ez elő sem fordulhatna, ha a folyamatot csak fizikai vagy kémiai egyensúlyok szabályoznák.
 közvetlenül a vízbe adják le. A Fe(III) redukciójához az oldott oxigén hiánya nem elégséges feltétel, mert az ezt tartalmazó flokkulumokat a hozzájuk asszociált szerves anyag stabilizálja. Ezért a vasoxidáló baktériumok (ezek a Fe[III]-at a szerves anyag oxidációja során keletkező elektronok akceptorának használják) jelenléte elengedhetetlen. Ha a klasszikus modell igaz lenne, akkor az üledék Fe2+ és PO43- kibocsátásának egyidejűen kéne bekövetkeznie. Ezzel szemben a kettő közt fáziseltolódás tapasztalható, s az a tény, hogy az antibiotikumokkal kezelt üledék P-kibocsátása lényegesen kisebb, mint a kezeletlené szintén a mikrobiális folyamatok fontosságára utal (Gächter és mtsi. 1988), sőt ez elő sem fordulhatna, ha a folyamatot csak fizikai vagy kémiai egyensúlyok szabályoznák.")
50
A foszforforgalom modern modellje: a P-kérdést biológiai kérdésnek gondolja
A 20. század utolsó évtizedeiben végzett kutatások mutattak rá arra, hogy a P kibocsátást inkább biológiai, mint kémiai folyamatok határozzák meg. Az üledék aerob körülmények mellett történő foszforkibocsátásában több tényező játszik szerepet, s ezek relatív fontossága térben és időben is változik. 1) A hőmérséklet gyorsítja a biológiai folyamatokat. A fotoszintézis intenzitásának növekedése pH emelkedéshez vezet, s a FeOOH flokkulumokban koprecipitált P ilyenkor OH--ra cserélődhet, ezáltal felszabadul (Søndergaard 1989). 2) Bioturbáció: Az üledéklakó gerinctelenek szűrésükkel, anyagcseretermékeik kibocsátásával, azzal, hogy folytonosan túrják az üledéket (ezeket a tevékenységeket nevezzük összefoglaló néven bioturbációnak) jelentősen befolyásolják annak kemizmusát, s a folyamat során P szabadulhat fel. A jelenség megítélése nem egyértelmű, mert az üledéklakó állatok légzése csökkenti, a bioturbáció viszont növeli az üledék oxigéntartalmát, s ezzel a P-felszabadulást meghatározó redoxi-viszonyokra ellentétes tényezők hatnak, melyek eredője térben és időben is változó. 3) Bevonatlakó szervezetek, elsősorban algák vékony, de igen aktív bevonatot képeznek az üledékfelszínen, mely a P-kibocsátás növekedéséhez és csökkenéséhez is vezethet időben változó módon. 4) A szél keltette hullámzás az üledék felső rétegét a nyíltvízbe keveri, s ezáltal a lényegesen nagyobb SRP tartalmú intersticiális víz is felszabadul. A Balaton belső foszforterhelésének ez igen lényeges összetevője, s a viharok során kiszabaduló foszfor vízvirágzásokat eredményezhet (Istvánovics és mtsi. 2004). 5) Az üledékben a baktériumok tevékenysége redukált gázokat termel, az üledék felszínén lévő algabevonat pedig oxigént. Mindkettő buborékok formájában akkumulálódhat, s amikor a légnyomás csökken e gázok a felszínre törnek. Az üledék felső rétegéből vagy felszínéről történő kibuborékolásuk a bioturbációhoz hasonló hatású lehet.
 A hőmérséklet gyorsítja a biológiai folyamatokat. A fotoszintézis intenzitásának növekedése pH emelkedéshez vezet, s a FeOOH flokkulumokban koprecipitált P ilyenkor OH--ra cserélődhet, ezáltal felszabadul (Søndergaard 1989). 2) Bioturbáció: Az üledéklakó gerinctelenek szűrésükkel, anyagcseretermékeik kibocsátásával, azzal, hogy folytonosan túrják az üledéket (ezeket a tevékenységeket nevezzük összefoglaló néven bioturbációnak) jelentősen befolyásolják annak kemizmusát, s a folyamat során P szabadulhat fel. A jelenség megítélése nem egyértelmű, mert az üledéklakó állatok légzése csökkenti, a bioturbáció viszont növeli az üledék oxigéntartalmát, s ezzel a P-felszabadulást meghatározó redoxi-viszonyokra ellentétes tényezők hatnak, melyek eredője térben és időben is változó. 3) Bevonatlakó szervezetek, elsősorban algák vékony, de igen aktív bevonatot képeznek az üledékfelszínen, mely a P-kibocsátás növekedéséhez és csökkenéséhez is vezethet időben változó módon. 4) A szél keltette hullámzás az üledék felső rétegét a nyíltvízbe keveri, s ezáltal a lényegesen nagyobb SRP tartalmú intersticiális víz is felszabadul. A Balaton belső foszforterhelésének ez igen lényeges összetevője, s a viharok során kiszabaduló foszfor vízvirágzásokat eredményezhet (Istvánovics és mtsi. 2004). 5) Az üledékben a baktériumok tevékenysége redukált gázokat termel, az üledék felszínén lévő algabevonat pedig oxigént. Mindkettő buborékok formájában akkumulálódhat, s amikor a légnyomás csökken e gázok a felszínre törnek. Az üledék felső rétegéből vagy felszínéről történő kibuborékolásuk a bioturbációhoz hasonló hatású lehet.")
51
Morfometriai hatások Az aerob hipolimnionnal rendelkező tavak számos közös morfometriai sajátsággal rendelkeznek: mélyek (emiatt vastag hipolimnionjuk igen sok oxigént tartalmaz a tavaszi átkeveredés után), área-koefficiensük kicsi (emiatt egyrészt kicsi a külső terhelés, másrészt a kis bejövő vízmennyiség miatt hosszú a retenciós idő). Az anyagmérleg vizsgálatok azt mutatták, hogy az ilyen tavakban a bejövő P-t inkább visszatartja (R) a tó üledéke semmint átmosódna rajta (15/1. táblázat), s a visszatartás mértéke a retenciós idővel fordítottan arányos. Az olyan tavak, melyek retenciós ideje > 10 év, jellemzően a bejutó P 70-90%-át visszatartják, s a retenciós idő csökkenésével a visszatartás is csökken. Sekélyebb tavaknál a bejövő nagyobb vízmennyiség (nagyobb áreakoefficiens) és emiatt rövidebb retenciós idő következménye, hogy az epilimnionban (ha rétegzettek egyáltalán) marad a bejövő P nagy része, ez itt nagyobb fitoplankton produkciót tesz lehetővé, ennek a tápanyaghálózatra gyakorolt hatásaival együtt. Az olyan rétegzett tavakban, melyek alacsony külső P és N terhelést kapnak a primer produkció alacsony, emiatt a szedimentáció is alacsony, a hipolimnion oxigénkészletét az ülepedő szerves anyag nem csökkenti jelentősen, üledékük ezért aerob marad. Az ilyen tavakban az üledékből eredő belső terhelés elhanyagolhatóan kicsi. E tavakra különösen igaz, hogy a globális foszforforgalom csapdáiként viselkednek. Az anoxikus hipolimnionnal rendelkező tavak vízmélysége tipikusan kisebb, (átlagmélyság < 10 m), retenciós ideje rövidebb, külső foszforterhelése nagyobb és a fitoplankton biomassza magasabb (huminanyagokkal terhelt tavak más tárgyalást igényelnek). Ennek megfelelően a kiülepedő anyagmennyiség is nagyobb, s arra elég, hogy a hipolimnion oxigénkészletét kimerítse, de arra nem, hogy a P az üledékben stabilizálódjon egyrészt, mert a rövid retenciós idő miatt kimosódik, másrészt mert az üledék is anaerob s ez nem kedvez a stabilizálódásnak. A 15/1-es táblázatban az aerob és anaerob tavak foszformérlegére vonatkozó becsült adatokat találhatunk.
, área-koefficiensük kicsi (emiatt egyrészt kicsi a külső terhelés, másrészt a kis bejövő vízmennyiség miatt hosszú a retenciós idő). Az anyagmérleg vizsgálatok azt mutatták, hogy az ilyen tavakban a bejövő P-t inkább visszatartja (R) a tó üledéke semmint átmosódna rajta (15/1. táblázat), s a visszatartás mértéke a retenciós idővel fordítottan arányos. Az olyan tavak, melyek retenciós ideje > 10 év, jellemzően a bejutó P 70-90%-át visszatartják, s a retenciós idő csökkenésével a visszatartás is csökken. Sekélyebb tavaknál a bejövő nagyobb vízmennyiség (nagyobb áreakoefficiens) és emiatt rövidebb retenciós idő következménye, hogy az epilimnionban (ha rétegzettek egyáltalán) marad a bejövő P nagy része, ez itt nagyobb fitoplankton produkciót tesz lehetővé, ennek a tápanyaghálózatra gyakorolt hatásaival együtt. Az olyan rétegzett tavakban, melyek alacsony külső P és N terhelést kapnak a primer produkció alacsony, emiatt a szedimentáció is alacsony, a hipolimnion oxigénkészletét az ülepedő szerves anyag nem csökkenti jelentősen, üledékük ezért aerob marad. Az ilyen tavakban az üledékből eredő belső terhelés elhanyagolhatóan kicsi. E tavakra különösen igaz, hogy a globális foszforforgalom csapdáiként viselkednek. Az anoxikus hipolimnionnal rendelkező tavak vízmélysége tipikusan kisebb, (átlagmélyság < 10 m), retenciós ideje rövidebb, külső foszforterhelése nagyobb és a fitoplankton biomassza magasabb (huminanyagokkal terhelt tavak más tárgyalást igényelnek). Ennek megfelelően a kiülepedő anyagmennyiség is nagyobb, s arra elég, hogy a hipolimnion oxigénkészletét kimerítse, de arra nem, hogy a P az üledékben stabilizálódjon egyrészt, mert a rövid retenciós idő miatt kimosódik, másrészt mert az üledék is anaerob s ez nem kedvez a stabilizálódásnak. A 15/1-es táblázatban az aerob és anaerob tavak foszformérlegére vonatkozó becsült adatokat találhatunk.")
52
Foszforforgalom tó Lext Lint %int RI zátl OXIKUS HIPOLIMNION
Cayuga, US 550 58 10 9 55 Aegeri, CH 160 -16 49 Turler, CH 300 -22 2 14 Superior, CA/US 30 -4 189 148 Michigan, US 100 -11 31 84 Huron, CA/US 70 48 59 Ontario, CA/US 680 -53 8 89 Zürich, CH 1320 -288 5 50 Washington, US 460 -6 3 33 Maggiore, CH/IT 3390 -1353 4 177 Clear, CA 40 1 13 Tahoe, US 42 714 303 [1] A Bergudasjöen szennyvíztisztítását 1974-re oldották meg, tehát ettől kezdődően az adatok a rekonstrukciót mutatják.

53
Foszforforgalom tó Lext Lint %int RI zátl ANOXIKUS HIPOLIMNION
Chautauqua, US 340 164 33 1,4 7 Saratoga, US 1600 -205 0,4 8 Chemung, US 220 26 11 4,2 9 Shagawa 1967, US 653 58 0,7 6 Harriett, US 710 355 0,8 Hallwiler, CH 550 69 4,0 28 Baldegger, CH 1750 485 22 4,5 34 Esrøm 1973, DK 600 78 12 17,0 Berundasjön, SE 1973[1] 8810 2954 25 1,0 2 1974 2110 3239 61 1975 410 1281 76 1976 240 1746 88 [1] A Bergudasjöen szennyvíztisztítását 1974-re oldották meg, tehát ettől kezdődően az adatok a rekonstrukciót mutatják.

54
Az oldott (DP) és a partikulált (PP) foszfor idealizált megoszlása oligotróf és eutróf tóban
 és a partikulált (PP) foszfor idealizált megoszlása oligotróf és eutróf tóban")
55
Foszforforgalom kooprecipitáció
A tavak foszforkészletének csökkenésében ill. a foszfor visszatartásában – mint láttuk – alapvető szerepe van a kimosódásnak és az ülepedésnek. Ez utóbbi egyik alapfolyamata a kemény vizű (sok Ca-ot és karbonát-bikarbonátot tartalmazó, jól pufferolt vizek) tavakban a biológiai kalcitprecipitáció (az alapfolyamat a fejezetben található). Ennek intenzitása a fotoszintézissel arányos, s miután az intenzív fotoszintézis pH növekedéshez vezet, ez utóbbi tovább gyorsítja a kristályképződést. A növekvő kristályok a foszfort adszorbeálják, kisebb mértékben beépülhet magába a kristályba. A CaCO3 kristályok növekedését gátló anyagok (pl. a magnézium és a vas) csökkentik a P koprecipitációját is. A CaCO3 túltelítettségének mértékében változik a kristályok morfológiája: alacsony túltelítettségnél romboéderes kristályok keletkeznek, ha nő a túltelítettség, akkor a kristályfprma komplexitása nő, dendritikus és oszlopszerű formák keletkeznek (Koschel 1990, 1997). Ez utóbbiak adszorpciós felülete nagyobb, emiatt több P megkötésére képesek A koprecipitáció jelentős mennyiségű P-t vonhat ki időlegesen vagy véglegesen az anyagforgalomból, a mérések szerint 25-35%-ban hozzájárulhat az epilitikus foszforkészlet csökkenéséhez (Kleiner 1988, Jäger és Röhrs 1990). A leülepedett kalcit kristályokban kötött P egy része a hipolimnionon való áthaladás során, az üledék felszínén vagy a legközelebbi cirkuláció során mobilizálódhat, más része azonban kötött marad. E folyamatok mértékére átlagot megállapítani nehéz lenne, mert erősen függenek az egyes tavak individuális sajátságaitól.
 tavakban a biológiai kalcitprecipitáció (az alapfolyamat a fejezetben található). Ennek intenzitása a fotoszintézissel arányos, s miután az intenzív fotoszintézis pH növekedéshez vezet, ez utóbbi tovább gyorsítja a kristályképződést. A növekvő kristályok a foszfort adszorbeálják, kisebb mértékben beépülhet magába a kristályba. A CaCO3 kristályok növekedését gátló anyagok (pl. a magnézium és a vas) csökkentik a P koprecipitációját is. A CaCO3 túltelítettségének mértékében változik a kristályok morfológiája: alacsony túltelítettségnél romboéderes kristályok keletkeznek, ha nő a túltelítettség, akkor a kristályfprma komplexitása nő, dendritikus és oszlopszerű formák keletkeznek (Koschel 1990, 1997). Ez utóbbiak adszorpciós felülete nagyobb, emiatt több P megkötésére képesek. A koprecipitáció jelentős mennyiségű P-t vonhat ki időlegesen vagy véglegesen az anyagforgalomból, a mérések szerint 25-35%-ban hozzájárulhat az epilitikus foszforkészlet csökkenéséhez (Kleiner 1988, Jäger és Röhrs 1990). A leülepedett kalcit kristályokban kötött P egy része a hipolimnionon való áthaladás során, az üledék felszínén vagy a legközelebbi cirkuláció során mobilizálódhat, más része azonban kötött marad. E folyamatok mértékére átlagot megállapítani nehéz lenne, mert erősen függenek az egyes tavak individuális sajátságaitól.")
56
Foszforforgalom kooprecipitáció

57
Eutrofizálódás Az eutrofizálódás definíciója Az eutrofizálódás a vizek növényi tápanyagokban – elsősorban foszforban és nitrogénben - való gazdagodását jelenti, melynek eredményeképp fokozott növényesedés jelentkezik, a víz zöldesre színeződik a planktonikus algáktól, felszíni vízvirágzások következnek be (planktonikus eutrofizálódás). Más esetekben az üledékfelszínt borítják sűrűn az alámerült hinarak, állományaik között vastag algaszőnyeg alakulhat ki, mely időnként a felszínre kerül s ott bomló csomókat képez (bentonikus eutrofizálódás). A keletkezett növényi tömeg – akár planktonikus, akár bentonikus - lebomlása oxigént fogyaszt, mely egy sor másodlagos hatást eredményez, mint pl. halpusztulások, korróziót okozó gázok vagy toxikus anyagok keletkezése, stb. (Vollenweider, 1989). Az eutrofizálódást nem annak oka (a növényi tápanyagok mennyiségének növekedése) vagy közvetlen következménye (algásodás vagy hinarasodás) teszik veszélyes és megakadályozandó jelenséggé, hanem az ezek kapcsán bekövetkező másodlagos hatások, melyek szinte minden humán vízhasználatot befolyásolnak (rekreáció, halászat, horgászat, közlekedés, ivóvíz nyerése, stb.) és közben az eredeti flóra és fauna is megváltozik.
. Más esetekben az üledékfelszínt borítják sűrűn az alámerült hinarak, állományaik között vastag algaszőnyeg alakulhat ki, mely időnként a felszínre kerül s ott bomló csomókat képez (bentonikus eutrofizálódás). A keletkezett növényi tömeg – akár planktonikus, akár bentonikus - lebomlása oxigént fogyaszt, mely egy sor másodlagos hatást eredményez, mint pl. halpusztulások, korróziót okozó gázok vagy toxikus anyagok keletkezése, stb. (Vollenweider, 1989). Az eutrofizálódást nem annak oka (a növényi tápanyagok mennyiségének növekedése) vagy közvetlen következménye (algásodás vagy hinarasodás) teszik veszélyes és megakadályozandó jelenséggé, hanem az ezek kapcsán bekövetkező másodlagos hatások, melyek szinte minden humán vízhasználatot befolyásolnak (rekreáció, halászat, horgászat, közlekedés, ivóvíz nyerése, stb.) és közben az eredeti flóra és fauna is megváltozik.")
58
Trofitási kategóriák TPéá: az összes P éves átlaga (μg L-1); TPtá: Az összes P mennyiságe a tavaszi átkeveredés időszakában (μg L-1); Chléá: a klorofilltartalom éves átlaga (μg L-1); Chlmax: a klorofill tartalom éves maximua (μg L-1); Secéá: a Secchi-átlátszóság éves átlaga (m); Secmin: a Secchi átlátszóság éves minimuma; Osat,min: a %-os oxigéntelítettség évei minimuma az üledék feletti cm-es rétegben; Npico: a pikofitoplankton egyedszámának éves átlaga (sejt mL-1); %piko: a pikofitoplankton százalékos részesedése az éves összbiomasszából és/vagy összprodukcióból. *: sekély, gyakran felkeveredő tavakra és folyókra nem használható; **: csak rétegzett tavakban használható; ***: savas vizekben nem használható ultra-oligotróf oligotróf mezotróf eutróf hipertróf 1TPéá < 4 4-10 10-35 35-100 > 300 2TPtá < 5 5-10 10-30 30-100 > 100 1Chléá < 1 1-2,5 2,5-8 8-25 > 25 1Chlmax < 2,5 25-75 > 75 1Secéá* > 12 12-6 6-3 3-1,5 < 1,5 1Secmin* > 6 1,5-7 < 0,7 1Osat,min** 90% 80% 90-40% 40-0% 10-0% 3Npico*** < 2x104 2x104 -2x105 2x105 -2x106 ? 108 -ig 3%piko*** > 80% 80-55% 55-25% > 25%
; TPtá: Az összes P mennyiságe a tavaszi átkeveredés időszakában (μg L-1); Chléá: a klorofilltartalom éves átlaga (μg L-1); Chlmax: a klorofill tartalom éves maximua (μg L-1); Secéá: a Secchi-átlátszóság éves átlaga (m); Secmin: a Secchi átlátszóság éves minimuma; Osat,min: a %-os oxigéntelítettség évei minimuma az üledék feletti cm-es rétegben; Npico: a pikofitoplankton egyedszámának éves átlaga (sejt mL-1); %piko: a pikofitoplankton százalékos részesedése az éves összbiomasszából és/vagy összprodukcióból. *: sekély, gyakran felkeveredő tavakra és folyókra nem használható; **: csak rétegzett tavakban használható; ***: savas vizekben nem használható. ultra-oligotróf. oligotróf. mezotróf. eutróf. hipertróf. 1TPéá. < > TPtá. < > Chléá. < ,5. 2, > 25. 1Chlmax. < 2, > 75. 1Secéá* > ,5. < 1,5. 1Secmin* > 6. 1,5-7. < 0,7. 1Osat,min** 90% 80% 90-40% 40-0% 10-0% 3Npico*** < 2x104. 2x104 -2x105. 2x105 -2x ig. 3%piko*** > 80% 80-55% 55-25% > 25%")
59
Egyéb trofitási kategóriák
Típus Jellemzők Atróf Terméketlen víz (pl. a csapvíz ennek nevezhető) Acidotróf Alacsony produktivitás, alacsony N és P szint, alacsony pH (< 5,5); e tavakban gyakran a C limitál Alkalitróf Magas produktivitás, magas Ca tartalom Argillotróf Alacsony produktivitás, melyet az agyagásványok jelenléte miatti rendkívül nagy zavarosság jellemez, fénylimitáltak Sziderotróf Alacsony produktivitás magas Fe koncentráció mellett Disztróf Alacsony produktivitás, sötét szín a nagy mennyiségű oldott huminanyag miatt, gyakran savasak is Politróf Az eu- és hipertróf típus közti átmeneti kategória egyes trofitási rendszerekben
 Acidotróf. Alacsony produktivitás, alacsony N és P szint, alacsony pH (< 5,5); e tavakban gyakran a C limitál. Alkalitróf. Magas produktivitás, magas Ca tartalom. Argillotróf. Alacsony produktivitás, melyet az agyagásványok jelenléte miatti rendkívül nagy zavarosság jellemez, fénylimitáltak. Sziderotróf. Alacsony produktivitás magas Fe koncentráció mellett. Disztróf. Alacsony produktivitás, sötét szín a nagy mennyiségű oldott huminanyag miatt, gyakran savasak is. Politróf. Az eu- és hipertróf típus közti átmeneti kategória egyes trofitási rendszerekben.")
60
A Liebig-féle minimumtörvény és a Redfield-arány
A tápanyaglimitáció megértéséhez legegyszerűbb a Liebig-féle minimumtörvényből kiindulni, mely kimondja, hogy a növények a számukra szükséges tápanyagokat mindig a legkisebb mennyiségben rendelkezésükre álló tápelem arányában veszik fel. Ha tehát egy adott tápelemtaralmú oldatról meg kívánjuk állapítani, hogy melyik van az igényekhez képest a legkisebb arányban jelen, akkor elsősorban az igényeket kell ismernünk. A tengeri fitoplankton vizsgálata alapján Redfield (Redfield és mtsi. 1963) írta le az ún. Redfield-arányt, mely atomos arányként a következő: 106 C : 16 N : 1 P : ~ 0,01 (Fe, Zn, Mn) : ~ 0,001 (Cu, Mo, Co, etc.)[1]. A tavak növényi tápanyagkészlete egyrészt kívülről bekerülő komponensekből áll, másrészt a tóban magában keletkező anyagok térülnek vissza a biológiai reciklizáció folyamán. E források mindegyike rendelkezik bizonyos tápelem aránnyal, mely sokszor eltér a Redfield aránytól. Minhogy a C általában nem limitáló elem, s azok sem, melyekre az igény < 0.01 atom, elegendő a N:P arányt részletesebben megvizsgálni. [1] A Redfield arány élőlénycsoportonként, de fajonként is jelentős eltéréseket mutathat. Kovaalgák esetén pl. a Si-ot is figyelembe kell venni, hisz növekedésüket ez alapvetően meghatározza. Másrészről éppen szilíciumvázuk adszorpciós kapacitása miatt a P-t igen hatékonyan kötik meg a vízből, s a sejtből való kiszivárgást is csökkenti a kovavázon való adszorpció. Lényegi eltérést okozhat az is, ha az adott populációt a fény, vagy valamely egyéb tápelem limitálja.
 írta le az ún. Redfield-arányt, mely atomos arányként a következő: 106 C : 16 N : 1 P : ~ 0,01 (Fe, Zn, Mn) : ~ 0,001 (Cu, Mo, Co, etc.)[1]. A tavak növényi tápanyagkészlete egyrészt kívülről bekerülő komponensekből áll, másrészt a tóban magában keletkező anyagok térülnek vissza a biológiai reciklizáció folyamán. E források mindegyike rendelkezik bizonyos tápelem aránnyal, mely sokszor eltér a Redfield aránytól. Minhogy a C általában nem limitáló elem, s azok sem, melyekre az igény < 0.01 atom, elegendő a N:P arányt részletesebben megvizsgálni. [1] A Redfield arány élőlénycsoportonként, de fajonként is jelentős eltéréseket mutathat. Kovaalgák esetén pl. a Si-ot is figyelembe kell venni, hisz növekedésüket ez alapvetően meghatározza. Másrészről éppen szilíciumvázuk adszorpciós kapacitása miatt a P-t igen hatékonyan kötik meg a vízből, s a sejtből való kiszivárgást is csökkenti a kovavázon való adszorpció. Lényegi eltérést okozhat az is, ha az adott populációt a fény, vagy valamely egyéb tápelem limitálja.")
61
log (chl-ay) = 0,91 log (Ply) – 0, 435
Az OECD-Vollenweider eutrofizációs modell Ha a tavak P ellátottsága és pl. alga biomasszája között keresünk összefüggést, soha nem a víz oldott foszfor tartalmát kell az algák mennyiségével összevetni. Ennek oka, hogy éppen azért mert az algák mennyiségi növekedése többnyire P-limitált, a hozzáférhető P-t azonnal beépítik szervezetükbe, s ezáltal annak alacsony koncentrációja megmarad. A Balatonban a leghipertófabb időszakban is szinte állandóan az analitikai méréshatár alatt volt az ortofoszfát koncentráció. A folyamat leírásához ezért az adott víz P-terhelése és növényi anyagmennyisége között kell összefüggést keresni. Sok tó éves P-terhelésének és klorofill-a tartalom formájában kifejezett alga biomasszájának analízise után írta le (15/3. ábra) Vollenweider és Kerekes (1980), ahol az éves klorofill-a koncentráció (chl-ay, mg m-3) és a foszforterhelés közt a következő az összefüggés: log (chl-ay) = 0,91 log (Ply) – 0, 435 ahol (Ply) = [L(P)/qs]/[1+(z/qs)1/2] és L(P) az éves P-terhelés (mg m-2), q (m3) az évente kicserélődő víz relatív mennyisége és z (m) a víztest átlagmélysége.
 Vollenweider és Kerekes (1980), ahol az éves klorofill-a koncentráció (chl-ay, mg m-3) és a foszforterhelés közt a következő az összefüggés: log (chl-ay) = 0,91 log (Ply) – 0, 435. ahol. (Ply) = [L(P)/qs]/[1+(z/qs)1/2] és L(P) az éves P-terhelés (mg m-2), q (m3) az évente kicserélődő víz relatív mennyisége és z (m) a víztest átlagmélysége.")
62
Tápanyagforrás vagy szervezet/produktum
N:P atomos arány N/P tömegarány Nem termékeny területekről érkező felszíni befolyás 547 247,51 Közepesen termékeny területekről érkező felszíni befolyás 166 75,11 Erdős területekről érkező felszíni befolyás 157 71,04 Településekről vagy mg.- területekről érkező felszíni befolyás 135 61,09 Termékeny területekről érkező felszíni befolyás 74 33,48 Nem szennyezett talajvíz 63 28,51 Csapadék 54 24,43 Erdős területekről érkező felszíni befolyás a trópusokon 52 23,53 Befolyás mezőgazdasági területekről 44 19,91 Folyóvíz (Missisipi) 27 12,22 Vízi makrofiton, magasabbrendű növények általában 24 10,86 Nem P limitált körülmények közt tartott algák 22 9,95 Átlagos műtrágyaösszetétel 17 7,69 Kiegyensúlyozottan növekvő fitoplankton (Redfield arány) 16 7,24 Állattartó telepek szennyvize 14 6,33 Háztartási szennyvíz (nem városi) A makrozooplankton által kiválasztott salakanyagok 11 4,98 Legeltetett területekről lefolyó vizek és városi szennyvíz 10 4,52 Baterioplankton protoplazma Szennyvíztisztító telepek kifolyó vize 6 2,71 Eutróf tó üledéke Libaürülék 2 0,90 Üledékes kőzetek Földkéreg < 0,2 < 0,09
 ,22. Vízi makrofiton, magasabbrendű növények általában ,86. Nem P limitált körülmények közt tartott algák ,95. Átlagos műtrágyaösszetétel ,69. Kiegyensúlyozottan növekvő fitoplankton (Redfield arány) 16. 7,24. Állattartó telepek szennyvize ,33. Háztartási szennyvíz (nem városi) A makrozooplankton által kiválasztott salakanyagok ,98. Legeltetett területekről lefolyó vizek és városi szennyvíz ,52. Baterioplankton protoplazma. Szennyvíztisztító telepek kifolyó vize. 6. 2,71. Eutróf tó üledéke. Libaürülék. 2. 0,90. Üledékes kőzetek. Földkéreg. < 0,2. < 0,09.")
63
Az OECD-Vollenweider eutrofizációs modell: eltérések a széleken

64
Az OECD-Vollenweider eutrofizációs modell: eltérések a széleken
nitrogén fény

65
A N- vagy fény limitálta szint beállása
Ha a terhelés folytatódik, s meghaladja egy bizonyos szintet (2. állapot), akkor lép abba a lineáris szakaszba, amit az OECD-Vollenweider összefüggés ír le. A lineáris növekedés azonban többféle okból és szinten is telítődésbe mehet. A Liebig-féle minimumtörvény szerint hiába növeljük a P mennyiségét, ha közben egy másik tápelem kerül a relatíve legkevesebb pozíciójába. Akkor ez az elem fog limitálni, s nem a P. A legelső „gyanúsított” természetesen a N. Ha tehát a tó N-forrásai limitálóvá válnak, akkor a chl-a/P összefüggés telítődésbe megy át az ún. N-meghatározta szinten, mert ekkortól nem a P limitálja a fitoplankton növekedését. A legtöbb esetben a fitoplankton biomassza növekedése azért nem áll be a N-meghatározta szinten, mert a fitoplankton összetételében nagy arányban jelennek meg azok a Nostocales fajok, melyek képesek a légköri N2 megkötésére, s ezáltal a rendszer N-limitáltságát kivédik. Vannak azonban esetek, amikor a fitoplankton mennyisége mégiscsak megáll a N-meghatározta szinten, s ennek okai is többfélék: A tó enyhén savas, és a planktonikus algák (köztük a N2-kötők) ilyen vizekben nem élnek meg (többek között azért, mert csak alkalikus foszfatázuk van, ezért savas közegben a P-t nem tudják felvenni). A víz súlyosan molibdénhiányos, e mikroelem hiányában pedig a nitrogenáz emzimkomplex – tekintve, hogy egyik alegysége Mo-t tartalmaz – létrejötte lehetetlen. Elsősorban trópusi vizek magas hőmérsékletű, anaerob hipolimnionjában fordul elő, hogy a denitrifikáció (14/4. ábra), melynek során gázállapotú N-vegyületek keletkeznek olyan mértékben fogyasztja a tó N-készletét, hogy azzal a N2-kötés nem tud lépést tartani. Fiatal krátertavakra geológiai okok miatt jellemző az alacsony N/P arány, ezért ezekben kialakulhat N-limitáltság. A fenti, kivételes esetektől eltekintve azonban a fitoplankton biomassza növekedése nem áll meg a N-meghatározta szinten, hanem a N2-kötésnek köszönhetően folytatódik. Növekedése nem lehet végtelen, mert előbb-utóbb annyi alga lesz már a vízben, hogy egymás elől árnyékolják el a fényt (3. állapot), s a fellépő fénylimitáltság beállítja az elvileg lehetséges maximális fitoplankton biomasszát, amit fény-meghatárizta szintnek tekinthetünk.
, akkor lép abba a lineáris szakaszba, amit az OECD-Vollenweider összefüggés ír le. A lineáris növekedés azonban többféle okból és szinten is telítődésbe mehet. A Liebig-féle minimumtörvény szerint hiába növeljük a P mennyiségét, ha közben egy másik tápelem kerül a relatíve legkevesebb pozíciójába. Akkor ez az elem fog limitálni, s nem a P. A legelső „gyanúsított természetesen a N. Ha tehát a tó N-forrásai limitálóvá válnak, akkor a chl-a/P összefüggés telítődésbe megy át az ún. N-meghatározta szinten, mert ekkortól nem a P limitálja a fitoplankton növekedését. A legtöbb esetben a fitoplankton biomassza növekedése azért nem áll be a N-meghatározta szinten, mert a fitoplankton összetételében nagy arányban jelennek meg azok a Nostocales fajok, melyek képesek a légköri N2 megkötésére, s ezáltal a rendszer N-limitáltságát kivédik. Vannak azonban esetek, amikor a fitoplankton mennyisége mégiscsak megáll a N-meghatározta szinten, s ennek okai is többfélék: A tó enyhén savas, és a planktonikus algák (köztük a N2-kötők) ilyen vizekben nem élnek meg (többek között azért, mert csak alkalikus foszfatázuk van, ezért savas közegben a P-t nem tudják felvenni). A víz súlyosan molibdénhiányos, e mikroelem hiányában pedig a nitrogenáz emzimkomplex – tekintve, hogy egyik alegysége Mo-t tartalmaz – létrejötte lehetetlen. Elsősorban trópusi vizek magas hőmérsékletű, anaerob hipolimnionjában fordul elő, hogy a denitrifikáció (14/4. ábra), melynek során gázállapotú N-vegyületek keletkeznek olyan mértékben fogyasztja a tó N-készletét, hogy azzal a N2-kötés nem tud lépést tartani. Fiatal krátertavakra geológiai okok miatt jellemző az alacsony N/P arány, ezért ezekben kialakulhat N-limitáltság. A fenti, kivételes esetektől eltekintve azonban a fitoplankton biomassza növekedése nem áll meg a N-meghatározta szinten, hanem a N2-kötésnek köszönhetően folytatódik. Növekedése nem lehet végtelen, mert előbb-utóbb annyi alga lesz már a vízben, hogy egymás elől árnyékolják el a fényt (3. állapot), s a fellépő fénylimitáltság beállítja az elvileg lehetséges maximális fitoplankton biomasszát, amit fény-meghatárizta szintnek tekinthetünk.")
66
Rekonstrukciós eljárások
A szennyvíztisztítók olyan tisztítási utólépéssel való felszerelése, melynek során a szennyvízből kicsapják a foszfort és csak ezek után engedik az így kezelt vizet az élővizekbe (a Balaton rekonstrukciója során ilyen rendszerrel szerelték fel a zalaegerszegi szennyvíztisztítót). 2) A szennyvíz elterelése, körcsatornázás (a Balaton keleti karéja körüli szennyvíz körcsatorna összeszedi a szennyvizeket, központi tisztítókba szállítja, majd az ott keletkezett tisztított szennyvizet más vízgyűjtőre emelik át). 3) Természetes vagy létesített vizenyős területeken vezetik át a terhelést adó folyóvizet. Azok vegetációja a P-t felveszi, a denitrifikáció pedig a N-eltávolítást szolgálja (pl. a Zala vize ma a Kis-Balaton tározón át jut a Balaton keszthelyi medencéjébe). 4) Tarvágott vagy erősen trágyázott mezőgazdasági területekről a lefolyó vizeket bizonyos szakaszokon dús parti vegetációval rendelkező „csíkokon” vezetik át, ahol azok a 3) pontban leírthoz hasonló módon veszítenek P és N készletükből, s csak ezután jutnak a védendő vízbázisba. 5) Az üledék foszforkészletének immobilizálása Al2(SO4)3 és FeCl3 mesterséges bevitelével, melynek hatására oldhatatlan AlPO4, FePO4, esetleg Fe(OOH)PO4 flokkulumok válnak ki (ehhez hasonló, az itt leírt 6-ossal kombinált módszerrel rehabilitáltak egy Bécs területén fekvő Duna holtágat, az Alte Donau-t; Donabaum & Dokulil 2001). 6) A P-gazdag szerves üledéket Ca(OH)2 hozzáadásával oxidálják, melynek eredményeképp oldhatatlan apatit [Ca10(PO4)5(OH)2] keletkezik. A mész adagolása pH növelő hatású, ekkor a vas flokkulumok formájában csapódik ki, mely szorpciós felületet biztosít a foszfornak, legalábbis amíg az üledék nem válik anaerobbá. Ez utóbbi megakadályozására szokás folyékony Ca(NO3)2 adagolása az üledékbe, mely a denitrifikációt segíti, ezzel növeli a víz és az üledék redoxpotenciálját, s lehetővé teszi, hogy a Fe(III) flokkulumok továbbra is csapdázzák a PO43- -t. Minedmellett a nitrát, mint elektronakceptor fokozza a szerves anyagok lebontását, s ezzel növeli a P mennyiségét. E példából láthatjuk, hogy a kémiai kezelések igencsak kétélű fegyverek.
. 2) A szennyvíz elterelése, körcsatornázás (a Balaton keleti karéja körüli szennyvíz körcsatorna összeszedi a szennyvizeket, központi tisztítókba szállítja, majd az ott keletkezett tisztított szennyvizet más vízgyűjtőre emelik át). 3) Természetes vagy létesített vizenyős területeken vezetik át a terhelést adó folyóvizet. Azok vegetációja a P-t felveszi, a denitrifikáció pedig a N-eltávolítást szolgálja (pl. a Zala vize ma a Kis-Balaton tározón át jut a Balaton keszthelyi medencéjébe). 4) Tarvágott vagy erősen trágyázott mezőgazdasági területekről a lefolyó vizeket bizonyos szakaszokon dús parti vegetációval rendelkező „csíkokon vezetik át, ahol azok a 3) pontban leírthoz hasonló módon veszítenek P és N készletükből, s csak ezután jutnak a védendő vízbázisba. 5) Az üledék foszforkészletének immobilizálása Al2(SO4)3 és FeCl3 mesterséges bevitelével, melynek hatására oldhatatlan AlPO4, FePO4, esetleg Fe(OOH)PO4 flokkulumok válnak ki (ehhez hasonló, az itt leírt 6-ossal kombinált módszerrel rehabilitáltak egy Bécs területén fekvő Duna holtágat, az Alte Donau-t; Donabaum & Dokulil 2001). 6) A P-gazdag szerves üledéket Ca(OH)2 hozzáadásával oxidálják, melynek eredményeképp oldhatatlan apatit [Ca10(PO4)5(OH)2] keletkezik. A mész adagolása pH növelő hatású, ekkor a vas flokkulumok formájában csapódik ki, mely szorpciós felületet biztosít a foszfornak, legalábbis amíg az üledék nem válik anaerobbá. Ez utóbbi megakadályozására szokás folyékony Ca(NO3)2 adagolása az üledékbe, mely a denitrifikációt segíti, ezzel növeli a víz és az üledék redoxpotenciálját, s lehetővé teszi, hogy a Fe(III) flokkulumok továbbra is csapdázzák a PO43- -t. Minedmellett a nitrát, mint elektronakceptor fokozza a szerves anyagok lebontását, s ezzel növeli a P mennyiségét. E példából láthatjuk, hogy a kémiai kezelések igencsak kétélű fegyverek.")
67
Rekonstrukciós eljárások
7) Az üledék felső rétegének kotrása. Ez drága eljárás, hatékonysága kétséges (a Balatonon pl. nem vált be, ezért abba is hagyták), mindemellett az üledék bolygatása olyan, már eltemetődött ciszták aktiválódásához vezethet, melyek vízvirágzást eredményeznek. Így alakult ki strandtisztító üledékkotrás következtében Dinobryon sociale vízvirágzás a Balaton keleti medencéjében 1993 májusában (Reynolds és mtsi., 1993). A lekotort üledékréteg elhelyezése mindig igen sok gondot jelent. 8) A hínárnövényzet aratása és eltávolítása (ezzel eleinte próbálkoztak a Kis-Balaton tározón); 9) Biomanipuláció, melynek során a planktonevő halak eltávolításával növelik a szűrő zooplankton mennyiségét, s ezek az algákat nagy mennyiségben kiszűrve a víz átlátszóságát növelik (rendszerint csak rövid távon hatékony megoldás). 10) Az anaerob hipolimnion levegővel történő buborékoltatása (a budapesti Feneketlen tavon működött ilyen berendezés). Ennek során a termoklin destabilizálódik, a tó rétegzettből polimiktikussá válhat, ezáltal a szél teljesen fel tudja keverni a nyári időszakban is, és ez segíti az atmoszferikus oxigéncserét.
 Az üledék felső rétegének kotrása. Ez drága eljárás, hatékonysága kétséges (a Balatonon pl. nem vált be, ezért abba is hagyták), mindemellett az üledék bolygatása olyan, már eltemetődött ciszták aktiválódásához vezethet, melyek vízvirágzást eredményeznek. Így alakult ki strandtisztító üledékkotrás következtében Dinobryon sociale vízvirágzás a Balaton keleti medencéjében 1993 májusában (Reynolds és mtsi., 1993). A lekotort üledékréteg elhelyezése mindig igen sok gondot jelent. 8) A hínárnövényzet aratása és eltávolítása (ezzel eleinte próbálkoztak a Kis-Balaton tározón); 9) Biomanipuláció, melynek során a planktonevő halak eltávolításával növelik a szűrő zooplankton mennyiségét, s ezek az algákat nagy mennyiségben kiszűrve a víz átlátszóságát növelik (rendszerint csak rövid távon hatékony megoldás). 10) Az anaerob hipolimnion levegővel történő buborékoltatása (a budapesti Feneketlen tavon működött ilyen berendezés). Ennek során a termoklin destabilizálódik, a tó rétegzettből polimiktikussá válhat, ezáltal a szél teljesen fel tudja keverni a nyári időszakban is, és ez segíti az atmoszferikus oxigéncserét.")
Hasonló előadás














 és a tápanyagterhelés számítása.>")





