Előadást letölteni
Az előadás letöltése folymat van. Kérjük, várjon

1
AZ IMMUNVÁLASZ LEFOLYÁSA IMMUNOLÓGIA INFORMATIKUS HALLGATÓKNAK
DR HOLUB MARCSILLA

2
Barrierek

3
Endogén: intenzív lokális immunreakció
Lokális gyulladás érpálya szövet Szekréció hízósejtek degranulációja génexpresszió Exogén: fertőzés, égés Endogén: intenzív lokális immunreakció

4
Lokális Tünetek: gyors és rövid ideig
Leukocitatoborzás Vazodilatáció Érpermeabilitás növekedés Leukociták érfalhoz kötődése megnőtt vérmennyiség leukociták, szolubilis faktorok szövetekbe jutnak Duzzanat (oedema) Bőrpír (rubor) Lokális melegérzés (calor) Fájdalom (dolor) (bradikinin) Funkciók kiesése (functio laesia) Lokális Tünetek: gyors és rövid ideig
 Bőrpír (rubor) Lokális melegérzés (calor) Fájdalom (dolor) (bradikinin) Funkciók kiesése (functio laesia) Lokális Tünetek: gyors és rövid ideig.")
5
barrier hámsérülés inflamresponse4.jpg” ábra alapján

6
az első percek, óra makrofág vérlemezkék komplement aktiválás NK C3 C3
reaktív oxigéngyökök NK IL-1 NO IL-6 TNF- IL-12 IL-8 vérlemezkék prosztaglandin

7
MAC (membrane attack complex)
Komplement aktiváció Alternatív útvonal MBL útvonal Klasszikus C1q komplex C3b C3bB C3bBb D factor MBL + MASP C4b, C2b C3b anaphylatoxinok C4a, C3a, C5a C5b C6-8 C9n MAC (membrane attack complex)
")
8
az első órák makrofágok komplement aktiválás hízósejt C5a, C3a
NK IL-1 C5a, C3a IL-6 hízósejt IL-8 IL-12 TNF- Prosztaglandin, LTB4 Hisztamin

9
az első órák makrofág A terület vörös, meleg, duzzadt fájdalmas
komplement aktiválás C3 C3 makrofág C3 C5a, C3a IL-1 IL-6 hízósejt NK TNF- IL-8 hisztamin IL-12 Hisztamin, LTB4 Prosztaglandin A terület vörös, meleg, duzzadt fájdalmas oedema

10
az első órák oedema komplement aktiválás makrofágok hízósejt C5a C3a
IL-1 IL-6 NK TNF- IL-8 prosztaglandin IL-12 Hisztamin oedema ér endothel

11
VÉRÉR erős adhézió gördülés kemotaxis diapedesis IL-8, C3a, C5a SZÖVET

12
órákon belül IL1, TNF-alfa IL-6 láz

13
órákon belül IL-6

14
Akutfázis reakció IL-6 CRP, C3 , B faktor, SAA, süllyedés , stb
néhány óra IL-6 Akutfázis reakció CRP, C3 , B faktor, SAA, süllyedés , stb

15
órákon belül IL-1 IL-6

16
órák - egy-két nap neutrofil-makrofág infiltráció vándorlás
Langerhans sejt (éretlen dendritikus sejt) vándorlás Környéki nyirokcsomó neutrofil-makrofág infiltráció
 vándorlás. Környéki nyirokcsomó. neutrofil-makrofág infiltráció.")
17
Korlátozott specificitás:
A felismerésben három stratégia Patogén eredetű nem saját Hiányzó saját Megváltozott saját Mikroorganizmusokra jellemző markerek, anyagcseretermékek (gazdaszervezetből hiányzik) Normál saját markerek esetén a saját elleni immunválasz blokkolt (mikroorganizmusokból hiányzik) Nem az egészséges sajátra jellemző markerek The concept of missing self was proposed to explain why natural killer (NK) cells preferentially kill target cells that express few or no MHC class I proteins on the cell surface (4). On the basis of these studies, it has been suggested that NK cells recognize the lack of self MHC-I as a signal for target cell lysis. The mechanism of missing-self recognition was worked out in studies that demonstrated that MHC-I is recognized by various inhibitory receptors, that upon binding to their ligands block the lytic activity of NK cells (5, 6). Inhibitory receptors expressed on NK and other cell types are characterized by a sequence motif called ITIM (immunoreceptor tyrosine inhibitory motif ). When the tyrosine in the ITIM is phosphorylated, it recruits protein tyrosine phosphatases SHP-1 and SHP-2, which in turn inhibit cytotoxic activity by dephosphorylating tyrosine residues critical for activating NK cells. Another wellknown example of the use of this strategy is the regulation of the alternative complement pathway (9). A key component of the complement, a serum protein called C3, can covalently and indiscriminately attach itself to the surfaces of autologous and microbial cells. This attachment occurs as a result of a spontaneous hydrolysis of a highly labile thioester bond in C3. However, the formation of an active protease complex of the alternative pathway, the C3 convertase, occurs only on the surfaces of microbial cells and not on autologous cells (9). This is because all self cells ubiquitously express several gene products, including CD46 and CD55, which inhibit formation of the active C3 convertase. Another example of a molecular signal that functions as a marker of normal self is the carbohydrate structures that decorate glycoproteins and glycolipids on the cell surface. In vertebrate animals, these usually terminate with sialic acids, which can be recognized by a variety of receptors involved in intercellular communication (10, 11). One group of receptors that can bind to sialated glycoproteins and glycolipids, but not to their desialated counterparts, comprise a family of proteins known as Siglecs (10). Similar to other inhibitory receptors, Siglecs are transmembrane proteins that contain ITIM motifs in their cytoplasmic regions. monitoring the expression of sialic acids on normal self-tissues and antigens (10). Lack of sialic acid expression on most microorganisms, and in some cases, on virally infected and transformed cells may be recognized as a missing-self signal. Siglecs expressed on macrophages, dendritic cells, and neutrophils may direct these cells to phagocytose microbial nonself (which lacks biosynthetic pathways for sialic acids) and abnormal self (which lost sialic acid expression because of infection, transformation, or senescence). Another Siglec called CD22 is expressed on B cells and presumably helps to prevent unwanted antibody responses against self-antigens by blocking B cell receptor signaling (12). Sialic acids also regulate the activation of the alternative complement cascade by binding to a serum protein called factor H, which upon binding to sialic acids on autologous normal cells inhibits formation of the C3 convertase
 Normál saját markerek esetén a saját elleni immunválasz blokkolt. (mikroorganizmusokból hiányzik) Nem az egészséges sajátra jellemző markerek. The concept of missing self was proposed to. explain why natural killer (NK) cells preferentially. kill target cells that express few or no. MHC class I proteins on the cell surface (4). On the basis of these studies, it has been. suggested that NK cells recognize the lack of. self MHC-I as a signal for target cell lysis. The mechanism. of missing-self recognition was worked. out in studies that demonstrated that MHC-I. is recognized by various inhibitory receptors, that upon binding to their ligands block the. lytic activity of NK cells (5, 6). Inhibitory. receptors expressed on NK and other cell. types are characterized by a sequence motif. called ITIM (immunoreceptor tyrosine inhibitory. motif ). When the tyrosine in the ITIM is. phosphorylated, it recruits protein tyrosine. phosphatases SHP-1 and SHP-2, which in. turn inhibit cytotoxic activity by dephosphorylating. tyrosine residues critical for activating. NK cells. Another wellknown. example of the use of this strategy is. the regulation of the alternative complement. pathway (9). A key component of the complement, a serum protein called C3, can covalently. and indiscriminately attach itself to. the surfaces of autologous and microbial. cells. This attachment occurs as a result of a. spontaneous hydrolysis of a highly labile. thioester bond in C3. However, the formation. of an active protease complex of the alternative. pathway, the C3 convertase, occurs only. on the surfaces of microbial cells and not on. autologous cells (9). This is because all self. cells ubiquitously express several gene products, including CD46 and CD55, which inhibit. formation of the active C3 convertase. Another example of a molecular signal. that functions as a marker of normal self is. the carbohydrate structures that decorate glycoproteins. and glycolipids on the cell surface. In vertebrate animals, these usually terminate. with sialic acids, which can be recognized by. a variety of receptors involved in intercellular. communication (10, 11). One group of receptors. that can bind to sialated glycoproteins. and glycolipids, but not to their desialated. counterparts, comprise a family of proteins. known as Siglecs (10). Similar to other inhibitory. receptors, Siglecs are transmembrane. proteins that contain ITIM motifs in their. cytoplasmic regions. monitoring the expression of sialic. acids on normal self-tissues and antigens. (10). Lack of sialic acid expression on most. microorganisms, and in some cases, on virally. infected and transformed cells may be. recognized as a missing-self signal. Siglecs. expressed on macrophages, dendritic cells, and neutrophils may direct these cells to. phagocytose microbial nonself (which lacks. biosynthetic pathways for sialic acids) and. abnormal self (which lost sialic acid expression. because of infection, transformation, or. senescence). Another Siglec called CD22 is. expressed on B cells and presumably helps to. prevent unwanted antibody responses against. self-antigens by blocking B cell receptor signaling. (12). Sialic acids also regulate the. activation of the alternative complement cascade. by binding to a serum protein called. factor H, which upon binding to sialic acids. on autologous normal cells inhibits formation. of the C3 convertase.")
18
A sejtfelszínen elhelyezkedő TLR-ek olyan ligandokat ismernek fel, amilyen egyáltalán nincs a gazdaszervezetben: pl. LPS, flagellin Az intracellulárisan elhelyezkedő TLR-ek nukleinsav alapú PAMP-ot ismernek fel. TLR3: dupla szálú RNS-t (dsRNS) TLR 7 és 8: (G + U)-gazdag egyszálú RNS-t (ssRNS) TLR9: virális és bakteriális nem metilált CpG DNS-t Compared with lipid or protein TLR ligands, nucleic acid TLR ligands, including CpG DNA, have distinct features in several aspects. Notably, nucleic acid TLR ligands show ambiguous distinction between the host and microorganisms. For example, LPS and flagellin are apparently exogenous and are not found at all in the host. However, unmethylated CpG motifs can be found in mammals, although the frequency is much lower than in bacteria or viruses. Furthermore, host-derived singlestranded RNAs and dsRNAs can also function as TLR7 and TLR3 stimulators, respectively. Thus the structural and functional differences between pathogen and host nucleic acids are not very prominent. Host-derived nucleic acids are quite unstable but can be accessible to recognition by TLRs. For example, host cells are incorporated by certainAPCsafter they die, and nucleic acids from ingested cells are then released into endosomes or lysosomes. This can cause potential danger for autoimmunity. Such danger, however, is evaded through several mechanisms. First, nucleic acid TLR ligands are recognized in different cellular compartments from lipid or protein TLR ligands. Lipid or protein TLR ligands are recognized on the plasma membrane, whereas nucleic acids are recognized by TLRs in the endosome. If manipulated to express on the plasma membrane, TLR9 can be stimulated by self DNA.29 Thus endosomal expression of TLR7/9 can prevent self DNA from being recognized, although it is beneficial for recognizing viral DNA. Second, TLR7 expression is very low in cells with high phagocytic activity.30 Third, the host possesses certain DNA sequences that can inhibit TLR9 signaling, for example in the telomere.31 Fourth, compared with bacterial RNA, host-derived RNA carries abundantly modified nucleosides. These modified RNAs are much less potent in dendritic cell (DC) activation than unmodified RNA.32 Furthermore, TLR7/9 expression and released nucleic acids can be localized in different cell compartments, although there is no formal proof of this. Nucleic acid TLR ligands can induce both IFN-a and IFN-b. TLR activation leading to type I IFNs contributes to the pathogenesis of autoimmune diseases, such as systemic lupus erythematosus (SLE). Protein–nucleic acid complexes can activate PDCs to produce IFN-a by coengaging TLR and FcR. Furthermore, the number of activated PDCs is increased in skin lesions in patients with SLE. As a result, serum IFN-a levels are increased in patients with SLE and correlate with the severity of the disease. This hypothesis can also explain deterioration of SLE manifestations after viral infection, which can trigger PDC activation. Autoreactive B cells can also be activated by protein– nucleic acid complexes (Fig 3).34 Rheumatoid factor (RF) is an Ig that can bind to certain types of IgG and that is often found in the sera of patients with autoimmune disorders. DNA–anti-DNA complexes are recognized by RF on the B-cell surface. RF BCR (B-cell receptor) mediates internalization and transport of these complexes into the endosome, where TLR9 is expressed. Then CpG DNA in the complex stimulates TLR9 and, in turn, activates B cells to produce RF. Dual engagement of bothBCRand TLR9 is mandatory for this type of autoreactive B-cell activation. Thus TLR signaling is critically involved in autoimmune pathogenesis.
 TLR 7 és 8: (G + U)-gazdag egyszálú RNS-t (ssRNS) TLR9: virális és bakteriális nem metilált CpG DNS-t. Compared with lipid or protein TLR ligands, nucleic. acid TLR ligands, including CpG DNA, have distinct. features in several aspects. Notably, nucleic acid TLR. ligands show ambiguous distinction between the host. and microorganisms. For example, LPS and flagellin are. apparently exogenous and are not found at all in the. host. However, unmethylated CpG motifs can be found. in mammals, although the frequency is much lower than. in bacteria or viruses. Furthermore, host-derived singlestranded. RNAs and dsRNAs can also function as TLR7. and TLR3 stimulators, respectively. Thus the structural. and functional differences between pathogen and host. nucleic acids are not very prominent. Host-derived nucleic. acids are quite unstable but can be accessible to recognition. by TLRs. For example, host cells are incorporated by. certainAPCsafter they die, and nucleic acids from ingested. cells are then released into endosomes or lysosomes. This. can cause potential danger for autoimmunity. Such danger, however, is evaded through several mechanisms. First, nucleic acid TLR ligands are recognized in. different cellular compartments from lipid or protein. TLR ligands. Lipid or protein TLR ligands are recognized. on the plasma membrane, whereas nucleic acids are. recognized by TLRs in the endosome. If manipulated to. express on the plasma membrane, TLR9 can be stimulated. by self DNA.29 Thus endosomal expression of TLR7/9. can prevent self DNA from being recognized, although. it is beneficial for recognizing viral DNA. Second, TLR7 expression is very low in cells with high phagocytic. activity.30 Third, the host possesses certain DNA. sequences that can inhibit TLR9 signaling, for example. in the telomere.31 Fourth, compared with bacterial RNA, host-derived RNA carries abundantly modified nucleosides. These modified RNAs are much less potent in dendritic. cell (DC) activation than unmodified RNA.32. Furthermore, TLR7/9 expression and released nucleic. acids can be localized in different cell compartments, although there is no formal proof of this. Nucleic acid TLR ligands. can induce both IFN-a and IFN-b. TLR activation leading to. type I IFNs contributes to the pathogenesis of autoimmune. diseases, such as systemic lupus erythematosus (SLE). Protein–nucleic acid complexes. can activate PDCs to produce IFN-a by coengaging TLR. and FcR. Furthermore, the number of activated PDCs is increased. in skin lesions in patients with SLE. As a result, serum. IFN-a levels are increased in patients with SLE and. correlate with the severity of the disease. This hypothesis. can also explain deterioration of SLE manifestations after. viral infection, which can trigger PDC activation. Autoreactive B cells can also be activated by protein– nucleic acid complexes (Fig 3).34 Rheumatoid factor (RF) is an Ig that can bind to certain types of IgG and that is. often found in the sera of patients with autoimmune disorders. DNA–anti-DNA complexes are recognized by RF. on the B-cell surface. RF BCR (B-cell receptor) mediates. internalization and transport of these complexes into the. endosome, where TLR9 is expressed. Then CpG DNA in. the complex stimulates TLR9 and, in turn, activates B cells. to produce RF. Dual engagement of bothBCRand TLR9 is. mandatory for this type of autoreactive B-cell activation. Thus TLR signaling is critically involved in autoimmune. pathogenesis.")
19
Felismerés - Több ponton
Szecernált PRR opszonizálás Endoszomális PRR Figure 4 | Dectin-1-mediated cellular responses to fungal β-glucans. Recognition of β-glucans on fungal particles induces several dectin-1-mediated cellular responses, which might contribute to anti-fungal immunity in vivo. These include fungal uptake and killing51,70 and the production of pro-inflammatory cytokines and chemokines, such as tumour-necrosis factor (TNF) and CXC-chemokine ligand 2 (CXCL2), in collaboration with the Toll-like receptors (TLRs)11,19, which is likely to lead to cellular recruitment and activation. Dectin-1-mediated recognition also stimulates the production of interleukin-12 (IL-12)11,31, which might result in a protective T-helper 1 (TH1)-cell response and the production of interferon-γ (IFN-γ), thereby activating the fungicidal activities of phagocytes. In dendritic cells, β-glucan recognition by dectin-1 can also induce the production of IL-10 and IL-2 (REF. 31), which could potentially contribute to the development of regulatory T cells, thereby limiting inflammatory pathology and promoting fungal persistence and long-term immunity, as proposed previously68. IL-10 would also inhibit the production of pro-inflammatory cytokines and chemokines. Fungi might also mask their β-glucan, by conversion from yeast to hyphal forms70. This could result in the induction of non-protective TH2-cell immune responses, mediated by IL-4 (REF. 68); this could be the result of preventing recognition by dectin-1 although the pathways leading to this response are unknown. Although dectin-1 is described here as having a central role in the generation of protective immune responses, it should be noted that many other opsonic and non-opsonic receptors (such as the mannose receptor, complement receptor 3 (CR3), dendritic-cell-specific ICAM3-grabbing non-integrin (DC-SIGN) and TLRs also contribute to this process. Brown GD. Dectin-1: a signalling non-TLR pattern-recognition receptor. Nat Rev Immunol Jan;6(1):33-43.
 and CXC-chemokine ligand 2 (CXCL2), in collaboration. with the Toll-like receptors (TLRs)11,19, which is likely to lead to cellular recruitment. and activation. Dectin-1-mediated recognition also stimulates the production of. interleukin-12 (IL-12)11,31, which might result in a protective T-helper 1 (TH1)-cell response. and the production of interferon-γ (IFN-γ), thereby activating the fungicidal activities of. phagocytes. In dendritic cells, β-glucan recognition by dectin-1 can also induce the. production of IL-10 and IL-2 (REF. 31), which could potentially contribute to the. development of regulatory T cells, thereby limiting inflammatory pathology and. promoting fungal persistence and long-term immunity, as proposed previously68. IL-10. would also inhibit the production of pro-inflammatory cytokines and chemokines. Fungi. might also mask their β-glucan, by conversion from yeast to hyphal forms70. This could. result in the induction of non-protective TH2-cell immune responses, mediated by IL-4. (REF. 68); this could be the result of preventing recognition by dectin-1 although the. pathways leading to this response are unknown. Although dectin-1 is described here. as having a central role in the generation of protective immune responses, it should. be noted that many other opsonic and non-opsonic receptors (such as the mannose. receptor, complement receptor 3 (CR3), dendritic-cell-specific ICAM3-grabbing. non-integrin (DC-SIGN) and TLRs also contribute to this process. Brown GD. Dectin-1: a signalling non-TLR pattern-recognition receptor. Nat Rev Immunol Jan;6(1):")
20
A makrofágon többféle receptor különféle bakteriális részeket ismer fel
A baktériumok makrofágokhoz kötődése citokinek és kis gyulladásos lipid mediátorok kibocsájtását indukálja A makrofágon többféle receptor különféle bakteriális részeket ismer fel A makrofágok bekebelezik és lebontják a megkötött baktériumokat

21
Neutrofil granulocita patogén elleni működése a gyulladás során
baktériumsejt Komplement receptor Fc-receptor

22
órák - egy-két nap neutrofil-makrofág infiltráció vándorlás
Langerhans sejt (éretlen dendritikus sejt) vándorlás Környéki nyirokcsomó neutrofil-makrofág infiltráció
 vándorlás. Környéki nyirokcsomó. neutrofil-makrofág infiltráció.")
23
nekrotikus vagy fertőzött sejt
kórokozó PAMP receptor dendritikus sejt Dendritikus sejt endogén aktivátorok nekrotikus vagy fertőzött sejt naív T sejt nyirokcsomó aktivált T sejt

24
kostimulációs molekula expresszió TLR
naív T-sejt immunológiai szinapszis érett dendritikus sejt

25
Ag bemutatása a T-sejteknek Ag feldolgozás Ag felvétel
Peptidek megjelenése MHC molekulán a felszínen Ag feldolgozás natív fehérjékből peptidek keletkeznek Ag felvétel

26
SPECIFIKUS ANTIGÉN FELISMERÉS
T-sejt B-sejt T-sejt receptor B-sejt receptor Feldolgozott peptid (lineáris epitóp) Teljes molekula (konformációs epitóp) Antigén Bemutató Sejt MHC
 Teljes molekula. (konformációs epitóp) Antigén Bemutató Sejt. MHC.")
27
MHC-I (HLA-A,B,C) MHC-II (HLA DR, DP, DQ)
 MHC-II (HLA DR, DP, DQ)")
28
MHC I + endogén peptid citotoxikus T- limfocita
elpusztít Vírus peptiddarabok MHC I

29
MHC II + exogén peptid helper T- limfocita
aktivál MHC II baci bakteriális peptiddarabok

30
Természetes immunitás
Adaptív immunitás Sejt-közvetített Humorális Dendritikus sejt

31
Patogének PRR-hez (TLR) kötődése egyidejűleg kiváltja a természetes és az adaptív immunválaszt, és ugyanakkor meghatározza az adaptív válasz irányát (humorális-celluláris) is. Magyar Tudomány, 2004/ o.

32
Az autoreaktív T-sejtek elleni védelmi mechanizmusok
Ignorancia Funkcionálisan elszigetelt antigének Immunprivilégizált hely Pl. szem Fizikailag gátolt felismerés Pl. agy-ér gát T-sejt deléció apoptózissal Potenciálisan autoreaktív T-sejt Anergia Szupresszió Ignorancia: tudatlanság Mechanisms that prevent potentially autoreactive T cells from reacting inappropriately to autoantigens. (a) Antigens that are sequestered from T cells (such as in the brain) cannot provoke an immune response; thus, the T cells are ‘ignorant’ of the autoantigen. (b) Similarly, some antigens, although not separated from T cells by a physical barrier, remain functionally separated in ‘immune-privileged sites’ where local expression of cytokines such as interleukin 10 (IL-10) and transforming growth factor b (TGF-b), or expression of the apoptotic Fas ligand (FasL), prevents the induction of an immune response. Other mechanisms of peripheral tolerance range from (c) removal of the T cell by active deletion, (d) conversion to ‘unresponsiveness’ (anergy), (e) development of the T cell into a more ‘tolerogenic’ phenotype such as T helper 2 (Th2) or (f) active suppression of the cell by a (regulatory) suppressor cell (fig001mhb). Regulátoros T-sejt által Tolerogén T-sejtté válás Az autoreaktív T-sejtek elleni védelmi mechanizmusok
 Antigens that are sequestered from T cells (such as in the brain) cannot provoke an immune response; thus, the T cells are ‘ignorant’ of the autoantigen. (b) Similarly, some antigens, although not separated from T cells by a physical barrier, remain functionally separated in ‘immune-privileged sites’ where local expression of cytokines such as interleukin 10 (IL-10) and transforming growth factor b (TGF-b), or expression of the apoptotic Fas ligand (FasL), prevents the induction of an immune response. Other mechanisms of peripheral tolerance range from (c) removal of the T cell by active deletion, (d) conversion to ‘unresponsiveness’ (anergy), (e) development of the T cell into a more ‘tolerogenic’ phenotype such as T helper 2 (Th2) or (f) active suppression of the cell by a (regulatory) suppressor cell (fig001mhb). Regulátoros. T-sejt által. Tolerogén T-sejtté válás. Az autoreaktív T-sejtek elleni védelmi mechanizmusok.")
33
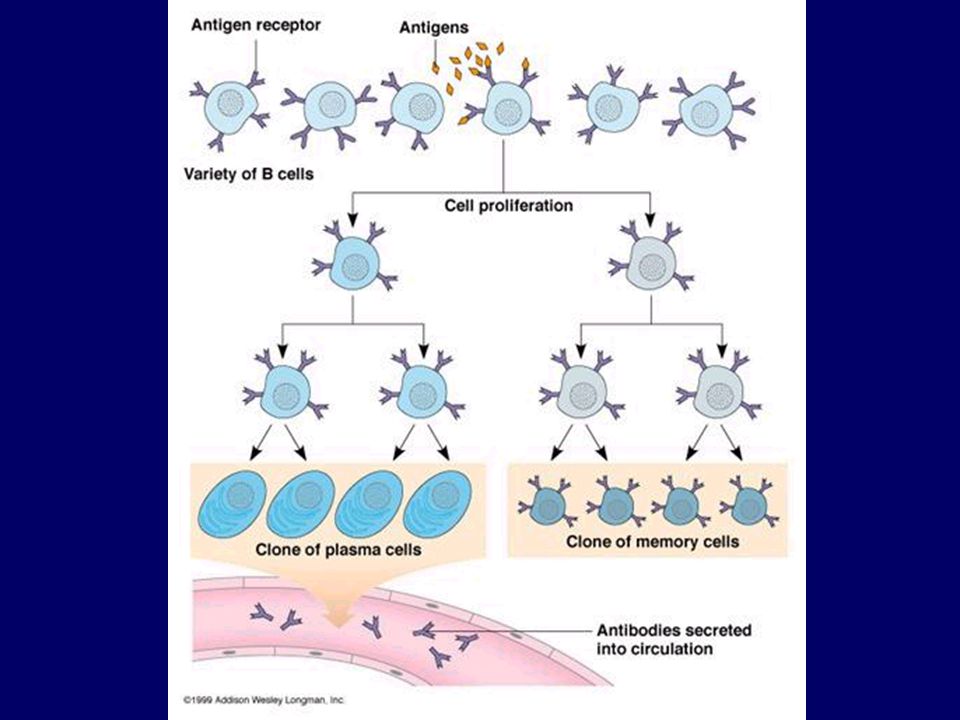
B sejt aktiváció: antigén + Th2
Antitest termelő plazma sejt

34
B-sejtek proliferációja plazmasejtté differenciálódása
Nyirokcsomó Centrum germinativum B-sejtek proliferációja affinitás érése, plazmasejtté differenciálódása nyirokcsomó duzzanat B sejtek proliferá-ciója

36
Az effektor T- és plazmasejtek a gyulladás színhelyére vándorolnak.

37
egy-két hét plazmasejtek effektor T sejtek antitestek

38
Antitest függő effektor mechanizmusok
neutralizálás opszonizálás Antitest függő celluláris citotoxicitás Komplement aktiválás Az antitest Fc régiója által mediált Fc

39
APC T SEJT MHC B7 B7 CD4 TCR CTLA 4 CD28 IL-2 mRNS CD3
POZITÍV KOSTIMULÁCIÓ NEGATÍV KOSTIMULÁCIÓ CTLA4 T CD28-nál nagyobb affinitással kapcsolódik B7-hez Több jelátviteli útvonal Transzkripciós faktorok aktiválódnak sejtmag IL-2 mRNS T SEJT

40
NEGATÍV KOSTIMULÁCIÓ antigén IgG FcR B-sejt Osztódás megáll
Fc-receptor Antitest-feedback

41
hetek-hónapok múlva A lokális gyulladás megszűnik, a helyi erek visszaalakulnak HEV-ből hagyományos postcapilláris venulákká

42
Hetek-hónapok múlva már csak a recirkuláló memória sejtek és antitestek emlékeztetnek a lezajlott fertőzésre B memória sejt T memória sejt

43
IMMUNVÁLASZ antitest primer szekunder IgG IgM

Hasonló előadás
















>")
>")

