Előadást letölteni
1
Fotoszintézis I. Alapfogalmak A fotoszintézis mint redox folyamat
A fotoszintetikus apparátus Pigmentek és pigment-protein komplexek A fény abszorpciója A gerjesztési energia sorsa A két fotokémiai rendszer Az elektrontranszport lánc felépítése

2
A fotoszintézis jelentősége
Az élővilág energia -forrása Szerves anyagok előállítása szervetlenekből (Az ábrán a keményítő szemcsék láthatók) A Föld légkörének oxigéntartalma ebből a folyamatból ered
 A Föld légkörének oxigéntartalma ebből a folyamatból ered.")
3
1) Bevezetés - redox folyamat - a fény és sötét szakasz - fényszakasz: két fotokémiai rendszer, PS II és PS I.

4
A fotoszintézis két fő szakasza: a fényreakciók és a sötét reakció

5
A fotoszintézis mint redox folyamat
H2D + A →fény→ H2A + D Ha víz a donor és CO2 az akceptor: 2nH2O + nCO2 →fény→ n(CH2O) + nH2O + nO2
 + nH2O + nO2.")
7
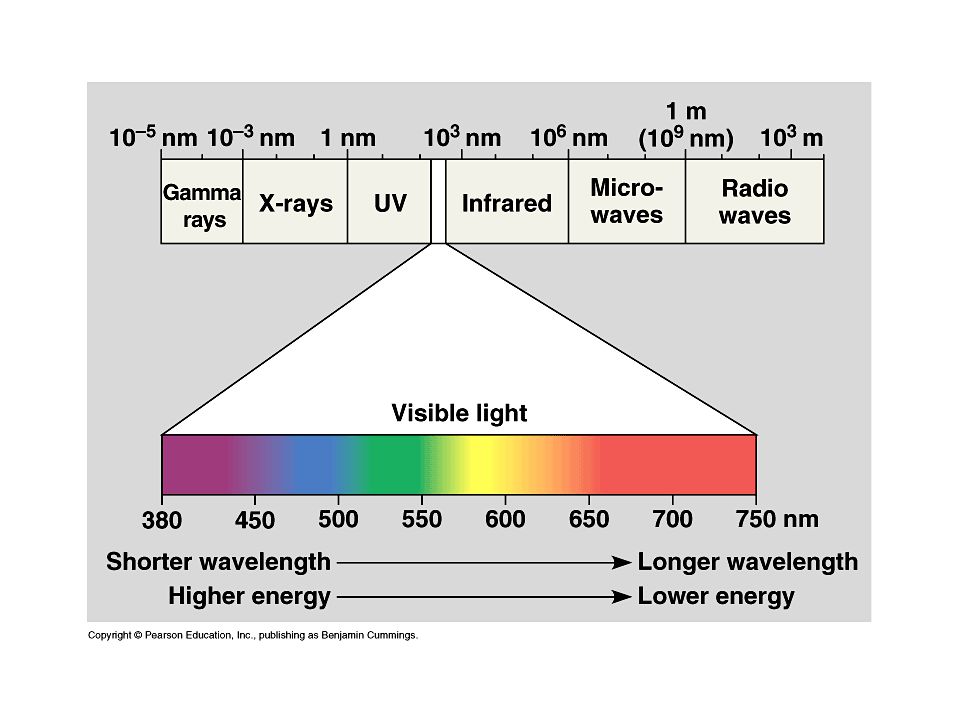
1. Fényszakasz: A fényt fotoszintetikus pigmentek abszorbeálják
A fényt abszorbeáló pigmentek a cianobaktériumokban és magasabbrendű növényekben: Kolorofill a, b Karotinoidok Fikobilinek (cianobaktériumokban és vörösmoszatokban)
")
8
A prizmával alkotóelemeire bontott fehér fény kék és vörös hullámhosszú komponenseivel megvilágított régió köré gyűlnek az aerob baktériumok a fotoszintetizáló Spirogyra körül

9
A levél keresztmetszete

10
14-35b A kloroplasztisz szerkezete
Kettős borítómembrán határolja Folyékony közege a sztróma –benne enzimek, keményítőszemcsék, DNS és riboszómák Membránrendszere tilakoidokból áll– ezek helyenként gránumokba rendeződik A tilakoidok belső üregei egy összefüggő, lumen nevezetű hálózatot alkotnak

12
A fotoszintetikus apparátus szerkezete
kloroplasztisz; tilakoid membránok gránum és sztróma kapcsolt és nem-kapcsolt felszínek a proteinkomplexek szerveződése a tilakoid membránban

13
A pigment-protein komplexek rendezetten helyezkednek a tilakoidmembránban

14
Honnan származik a kloroplasztisz?

15
14-41 A fénybegyűjtő pigmentek: Klorofill
Porfirin gyűrű – négy pirrolgyűrűből áll A négy pirrolgyűrű mellett egy öttagú ciklopentanon gyűrű található A konjugációs rendszer kékkel jelölve Magnézium ion – Mg++ - narancs Észter kötéssel csatlakozó fitol – zöld Ez a klorofilla – a többi klorofill kicsit külőnbözik.

16
Klorofillok

17
A karotinoidok járulékos pigmentek

18
Fikobilinek: cianobaktériumokban, vörös algákban
A pirrol gyűrűk nem záródnak porfirin gyűrűvé

19
Pigment-protein komplexek:
Klorofill - protein komplexek: a) CC I. (P-700), I. reakciócentrum b) LHC I. I. fénybegyűjtő komplex apoprotein kód a sejtmagban c) CC II. (P-680), II. reakciócentrum (6 fehérje) d) LHC II. II. fénybegyűjtő komplex Fikobiliproteinek allofikocianin, fikocianin, fikoeritrin = fikobiliszóma
 CC I. (P-700), I. reakciócentrum. b) LHC I. I. fénybegyűjtő komplex. apoprotein kód a sejtmagban. c) CC II. (P-680), II. reakciócentrum (6 fehérje) d) LHC II. II. fénybegyűjtő komplex. Fikobiliproteinek. allofikocianin, fikocianin, fikoeritrin = fikobiliszóma.")
21
A fényenergia abszorpciója

22
A fény abszorpciója és a gerjesztési energia sorsa
Fotoszintetikusan aktív fény ~ nm-ig Klorofill molekula: alapállapot (S0), Egy p- elektron kötő pályáról lazító pályára lép. szinglet (S1 és S2) és triplet (T1) állapotok vörös fény elnyelése: S állapot kék fény elnyelése: S2 állapot A gerjesztési energia sorsa: hővé alakul, fluoreszcencia, fotokémiai reakciók (redox folyamatok) energia migráció vagy energia transzfer (azonos vagy kémiailag eltérő pigmenteknek adódik át az energia)
, Egy p- elektron kötő pályáról lazító pályára lép. szinglet (S1 és S2) és. triplet (T1) állapotok. vörös fény elnyelése: S1 állapot. kék fény elnyelése: S2 állapot. A gerjesztési energia sorsa: hővé alakul, fluoreszcencia, fotokémiai reakciók (redox folyamatok) energia migráció vagy energia transzfer. (azonos vagy kémiailag eltérő pigmenteknek adódik át az energia)")
23
A klorofill fluoreszcenciája akkor is a vörös hullámhossz tartományba esik, ha kék fényt nyel el
Ok: az S2-S1 átmenet mindig hő formájában disszipálódik

24
A fény abszorpciója és a gerjesztési energia sorsa Kvantumhatásfok és kvantum szükséglet
Kvantumhatásfok (Φ): azon excitált molekulák hányada, amelyek egy bizonyos módon veszítik el energiájukat. Értéke 0-1. Pl. fotokémiai reakciók száma/összes abszorbeált kvantumok száma (ált. 0.95) Kvantum szükséglet: az egy bizonyos reakciótermék (pl. O2) létrejöttéhez szükséges kvantumok száma, azaz 1/ Φ.
: azon excitált molekulák hányada, amelyek egy bizonyos módon veszítik el energiájukat. Értéke 0-1. Pl. fotokémiai reakciók száma/összes abszorbeált kvantumok száma (ált. 0.95) Kvantum szükséglet: az egy bizonyos reakciótermék (pl. O2) létrejöttéhez szükséges kvantumok száma, azaz 1/ Φ.")
25
A fluoreszcencia imázs analízis a lézerrel egy ponton gerjesztett fluoreszcencia terjedését mutatja a levél felszínén (a megvilágított minták effektív kvantumhatékonyságának a képe)
")
26
A növényzet fluoreszcenciája műholdas felvételeken is detektálható
Kaliforniai öböl, algavirágzás, október 6., számítógépes színek, a kék az alacsony, a vörös a nagy intenzitású fluoreszcencia

27
Az antennapigmentek az elnyelt fényenergiát a reakciócentrum klorofilla felé továbbítják
A fényenergia vándorlása: antenna pigmentek között: külső és belső antennák reakciócentrumok klorofill a molekulájához: II. fotokémiai rendszer: P680, I. fotokémiai rendszer: P700

28
Energiaátadás két pigment molekula között akkor történik, ha a donor molekula abszorpciós vagy fluoreszcencia spektruma részben fedi egymást

29
Energia migráció, energia transzfer és fotokémiai folyamatok
14-43 Energia migráció, energia transzfer és fotokémiai folyamatok A fénygyűjtő komplex (LHC II. és I.) a kísérőpigmentek által elnyelt fényenergiát a reakciócentrum klorofillokhoz közvetíti. A fénybegyűjtő komplexek pigmentjei magasabbrendű növényekben klorofill a és b, karotin és xantofill molekulák, gyakran fehérjékhez kötődve. A gerjesztett pigmentek energiája rezonancia által átadódik a reakciócentrumba. A fényenergia kémiai energiává alakulása: töltésszeparációval Donor Kla Akceptor; Donor Kla Akceptor Donor Kla Akceptor; Donor Kla Akceptor
 a kísérőpigmentek által elnyelt fényenergiát a reakciócentrum klorofillokhoz közvetíti. A fénybegyűjtő komplexek pigmentjei magasabbrendű növényekben klorofill a és b, karotin és xantofill molekulák, gyakran fehérjékhez kötődve. A gerjesztett pigmentek energiája rezonancia által átadódik a reakciócentrumba. A fényenergia kémiai energiává alakulása: töltésszeparációval. Donor Kla Akceptor; Donor Kla Akceptor. Donor Kla Akceptor; Donor Kla Akceptor")
30
A két fotokémiai rendszer Korai bizonyítékok
A „vörös esés” A „kromatikus átmenetek”

31
A két fotokémiai rendszer Korai bizonyítékok
Az „antagonisztikus effektus” Az „erősítési effektus”

32
1. A PS II felépítése - reakciócentrum: D1 és D2 fehérjék: ehhez kapcsolódnak a feofitin (primer e- akceptor), a QA és QB kinon akceptorok, D1 fehérje tirozin aminosava (primer e- donor) Mn2+, Mn3+ ionok (vízbontás) - reguláló sapka - proximális antenna - disztális antenna 2. Funkciói vízbontás: 2H2O O2 +4H+ + 4e- proton keletkezik a lumenben, elektron lép az e- transzportláncba a plasztokinon redukciója
 Mn2+, Mn3+ ionok (vízbontás) - reguláló sapka. - proximális antenna. - disztális antenna. 2. Funkciói. vízbontás: 2H2O O2 +4H+ + 4e- proton keletkezik a lumenben, elektron lép az e- transzportláncba. a plasztokinon redukciója.")
33
A PS II felépítése

34
A PS I felépítése Az I. fotokémiai rendszer Reakciócentrum:
heterodimer kDa fehérje P-700; A0 (10 ps);, A1 (50 ps): elsődleges, másodlagos e-akceptorok (kla; ill. K-vitamin- fillokinon) Vas-kén centrumok: FX, FB, FA (4Fe-4S)
;, A1 (50 ps): elsődleges, másodlagos e-akceptorok. (kla; ill. K-vitamin- fillokinon) Vas-kén centrumok: FX, FB, FA (4Fe-4S)")
35
A PS I felépítése

36
Az elektrontranszportlánc felépítése
- membránhoz kötött komponensek PS II, citokróm b6/f komplex, PS I, ferredoxin-NADP+ reduktáz - mobilis komponensek plasztokinon pool, PQH2 (membránban, PS II. és citokróm b6/f komplex között) plasztocianin, PC (lumen, citokróm b6/f komplex és PS I. közt) ferredoxin (PS I. és ferredoxin-NADP+ reduktáz között)
 plasztocianin, PC (lumen, citokróm b6/f komplex és PS I. közt) ferredoxin (PS I. és ferredoxin-NADP+ reduktáz között)")
37
Energiaszint változások a fotoszintézis folyamán

38
14-47 Az elektrontranszportlánc két formája
A nem ciklusos elektrontranszport (Z séma) A ciklikus variációban a ferredoxin a citokróm b6/f komplexnek adja át az elektront. A fény ily módon a PSI egyedüli részvételével körbe hajtja az elektronokat és így hoz létre H+ koncentráció-különbséget a sztróma és a luminális tér között. Ez ATP szintézisre használódik fel.
 A ciklikus variációban a ferredoxin a citokróm b6/f komplexnek adja át az elektront. A fény ily módon a PSI egyedüli részvételével körbe hajtja az elektronokat és így hoz létre H+ koncentráció-különbséget a sztróma és a luminális tér között. Ez ATP szintézisre használódik fel.")
39
A PS II energia szint változásai és kinetikai paraméterei

40
Az LHC II szabályozza az energiamegoszlást a PS II és PS I között

41
Kautsky effektus II. A fluoreszcencia indukció és kioltás szakaszai
F0 = minimális fluoreszcencia Fm = maximális fluoreszcencia Fv = változó fluoreszcencia Fv = Fm - F0 ; Fv/Fm max. 0,84 D → P; PSII → PSI e- átmenet Fluoreszcencia kioltás: fotokémiai és nem-fotokémiai P → M → T szakaszok

42
A vízbontó enzim állapotváltozásai
S0 : S1 : S2 : S3 = 0,25 : 0,75 : 0 : 0

43
A plasztokinon mint mobilis komponens

44
A plasztokinon mint mobilis komponens

45
A citokróm b6/f komplex működése oxidálja a PQH2-t, redukálja a PC-t
a ciklusos elektrontranszportban oxidálja a ferredoxint proton átadást közvetít a sztrómából a lumenbe Fd Fd -50 mV -150 mV -50 mV -150 mV

46
A PS I. felépítése és működése
Reakciócentrum: 2 db centrális fehérje P700 klorofilla elsődleges (Kla) és másodlagos (K-vitamin) akceptorok vas-kén centrumok PC és ferredoxin kötő fehérjék Nem ciklusos elektrontranszport: az e- a NADP+ felé, Ciklusos elektrontranszport: e- a citokróm b6/f komplex felé Ferredoxin, ferredoxin-NADP+ reduktáz, NADP+ Eredmény: NADPH + H+
 és másodlagos (K-vitamin) akceptorok. vas-kén centrumok. PC és ferredoxin kötő fehérjék. Nem ciklusos elektrontranszport: az e- a NADP+ felé, Ciklusos elektrontranszport: e- a citokróm b6/f komplex felé. Ferredoxin, ferredoxin-NADP+ reduktáz, NADP+ Eredmény: NADPH + H+")
47
Ferredoxin-NADP reduktáz
LUMEN SZTRÓMA

48
A kemiozmotikus mechanizmus. Fotofoszforiláció.
Peter Mitchell 1963; Nobel díj 1978: Kapcsolódás a H+ elektrokémiai potenciálgradiens és a sejt munkavégzése között szelektíven permeábilis membránon keresztül: μH = p = ψ – 2.3RTpH /F elektrokémiai proton membrán pH gradiens H+ gradiens mozgató potenciál p = ψ – 59pH (mV) ATP szintáz (F0-F1 típusú ATPáz) – fotofoszforiláció
 ATP szintáz (F0-F1 típusú ATPáz) – fotofoszforiláció.")
49
8. A fotoszintetikus foszforiláció: a Mitchell-f
8. A fotoszintetikus foszforiláció: a Mitchell-f. kemiozmotikus mechanizmus Jagendorf: az ATP képződés mechanizmusa, bizonyítás CF0/CF1 ATP szintáz felépítése ATP szintézis

50
A fotoszintetikus foszforiláció: kemiozmotikus modell
proton elektrokémiai potenciálgrádiens a lumen és a sztróma között: ezt használja fel az ATP- szintáz enzim az ATP szintézisére ADP-ből és anorganikus foszfátból

51
Az elektontranszportlánc és a foszforiláció
A két folyamat között olyan szoros a kapcsolat hogy ATP-képződés nélkül az elektrontanszportlánc leáll és fordítva. Szétkapcsoló anyagok megszüntetik ezt a szoros kapcsolatot. Ilyen szétkapcsoló anyagok a következők: CCCP, valinomicin, gramicidin D, DCPIP stb. CCCP = karbonilcianid m-klorofenil hidrazon DCPIP = 2,6-diklorofenol indofenol

52
ÖSSZEFOGLALÁS













 gáz, vagy gázelegy Lézerátmenet: elektronszintek között (UV és látható lézerek) rezgési szintek.>")








