Előadást letölteni
1
Fotoszintézis III. The Dark Biochemistry A CO2 asszimilációja:
avagy a sötét szakasz… Fotoszintézis III. A CO2 asszimilációja: fixáció és redukció Dr. Horváth Ferenc

2
2n H2O + n CO2 n (CH2O) + n H2O + n O2
redukált termék redukció oxidáció fény Ha nem fotoszintetikus úton előállított ATP-t és NADPH-t adunk sötétben lévő kloroplasztiszokhoz, akkor ugyanúgy megtörténik a CO2-fixáció, mintha megvilágítottuk volna őket.

3
A C2-es fotorespirációs (karbon oxidációs, glikolát) ciklus
M. Calvin ( ) tisztázta elsőként a CO2-fixáció folyamatát, azóta több alternatív utat is feltártak Melvin Calvin, Nobel díj 1961. A C3-as (primer termék 3-szénatomos molekula) fotoszintetikus szénredukció (Calvin-ciklus) A C2-es fotorespirációs (karbon oxidációs, glikolát) ciklus A CO2 fixáció C4-es útja (CO2 koncentráló mechanizmus) A CO2 fixáció C4-es útja a CAM növényekben A szacharóz és a keményítő szintézise és szabályozási mechanizmusok a CO2 asszimilácójában
 tisztázta elsőként a CO2-fixáció folyamatát, azóta több alternatív utat is feltártak. Melvin Calvin, Nobel díj A C3-as (primer termék 3-szénatomos molekula) fotoszintetikus szénredukció (Calvin-ciklus) A C2-es fotorespirációs (karbon oxidációs, glikolát) ciklus. A CO2 fixáció C4-es útja (CO2 koncentráló mechanizmus) A CO2 fixáció C4-es útja a CAM növényekben. A szacharóz és a keményítő szintézise. és szabályozási mechanizmusok a CO2 asszimilácójában.")
4
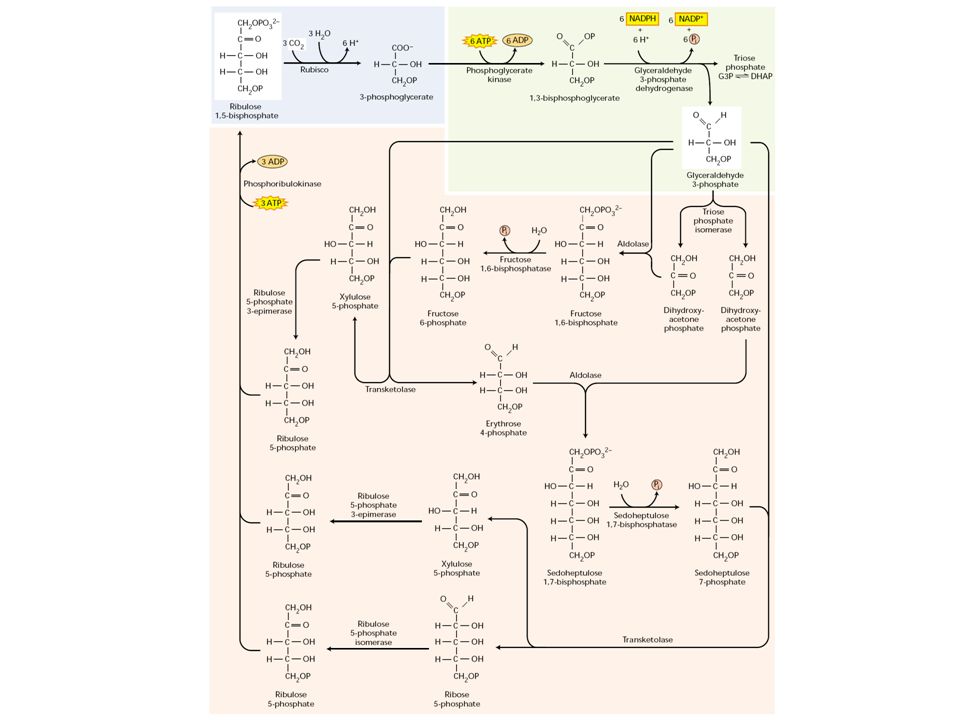
1. A C3-as fotoszintetikus szénredukció (Calvin-ciklus)
5 másodperces megvilágítás után forró metanolban fixálva, és 2D papírkromatográfia PGA- glicerinsav-3-foszfát 30 másodperces megvilágítás után forró metanolban fixálva, és 2D papírkromatográfia Szénben jelzett hidrogénkarbonát ion, H14CO3- adagolásával a széndioxid fixálás elsődleges termékei kimutathatók

5
A Calvin-ciklus három fázisra tagolható
Ribulóz-1,5- biszfoszfát Glicerinsav-3-foszfát Glicerinaldehid- 3-foszfát Szacharóz, keményítő

6
1. A sötétszakasz kezdete: karboxiláció
A Calvin-ciklus folyamán a C3-as úton a széndioxid a ribulóz-1,5-biszfoszfáthoz kötődik és egy feltételezett hat szénatomos intermedier terméken keresztül két molekula glicerinsav-3-foszfáttá alakul át. A glicerinsav-3-foszfát a fotoszintetikus CO2-fixáció első stabil terméke

7
RUBISCO Ribulóz-1,5-biszfoszfát-karboxiláz-oxigenáz a reakciót katalizáló enzim Kettős enzimaktivitású – karboxiláz és oxigenáz (3:1) Kompetíció, ha mindkét szubsztrátum jelen van A kloroplasztisz sztrómájában található

8
A RUBISCO SZERKEZETE L8S8 alegységek (56 kDa, kloroplasztiszban szintetizálódik; 14 kDa, sejtmagban kódolt) small (rbcS) and the large (rbcL) subunit of rubisco A katalitikus hely a nagyobbik alegységen található
 and the large (rbcL) subunit of rubisco. A katalitikus hely a nagyobbik alegységen található.")
9
Rubisco: a levél összes fehérje tartalmának több, mint 50%-a
N-raktár, a Föld népességének minden tagjára 20 kg jut… Évente 200 milliárd tonna CO2-t fixál A fehér és szürke a nagy alegység dimereket, a narancs és kék a kis alegység dimereket mutatja.

10
A Rubisco aktiválása karbamilációval
Az aktiváció CO2 és Mg2+ megkötésével és proton felszabadításával jár, így a megvilágítás hatására bekövetkező sztróma pH emelkedés és a Mg2+ koncentráció növekedés serkenti az aktivációt.

11
A Rubisco-aktiváz működése
A Rubisco-aktiváz eltávolítja a kötött RuBP-ot az inaktív, dekarbamilált Rubisco-ról egy ATP-t igénylő reakcióban. A szabad Rubisco ezután CO2 és Mg2+ kötésével karbamilálódhat. A Rubisco-aktivázt a fény a tioredoxin-rendszeren keresztül aktiválja (később).
.")
12
2. A redukciós fázis kezdete
Foszforilációs lépés 3-foszfoglicerát-kináz (PGA) Ha a glicerinsav-3-foszfát mennyisége nagyobb, mint a glicerinsav-1,3-biszfoszfáté, akkor a reakció a jelzett irányban játszódik le (glikolízisben fordítva) Az ATP mennyiség 2/3-a itt használódik fel.
 Ha a glicerinsav-3-foszfát mennyisége nagyobb, mint a glicerinsav-1,3-biszfoszfáté, akkor a reakció a jelzett irányban játszódik le (glikolízisben fordítva) Az ATP mennyiség 2/3-a itt használódik fel.")
13
Redukciós lépés Glicerinaldehid-3-foszfát dehidrogenáz
A NADPH mennyisége limitálja a reakciót, az enzim aktivitását a fény növeli

14
A regenerációs fázis kezdete
G3P – DHAP izomer átalakulás Trióz-foszfát izomeráz DHAP:G3P = 22:1 A regenerációs fázis kezdete Aldoláz

15
Fruktóz-1,6-biszfoszfatáz
eritróz-4-foszfát xilulóz-5-foszfát Transzketoláz

16
Szedoheptulóz-1,7-biszfoszfatáz
Aldoláz szedoheptulóz-1,7-biszfoszfát Szedoheptulóz-1,7-biszfoszfatáz Transzketoláz ribóz-5-foszfát xilulóz-5-foszfát

17
Ribulóz-5-foszfát epimeráz
izomerizáció Ribóz-foszfát-izomeráz Foszforibulo-kináz v. ribulóz-5-foszfát kináz - fény-aktivált enzim - a regeneráció zárólépése - az ATP 1/3-a itt használódik fel

20
A sötét szakasz legfontosabb biokémiai folyamatai
- karboxilációs fázis ribulóz-1,5-bifoszfát + CO2 = 2 glicerinsav-3-foszfát Katalizálja: a ribulóz-1,5-bifoszfát-karboxiláz, oxigenáz (RUBISCO) - redukció szakasza foszforiláció: az ATP 2/3 részének felhasználása redukció: a redukált NADPH felhasználása a glicerinsav-3- foszfát redukciójára - regeneráció a ribulóz-1,5-biszfoszfát-karboxiláz regenerálása: 6 glicerinsav-3-foszfát 3 ribulóz-1,5-biszfoszfát + 1 trióz-foszfát Nyereség ciklusonként egy trióz-foszfát!
 - redukció szakasza. foszforiláció: az ATP 2/3 részének felhasználása. redukció: a redukált NADPH felhasználása a glicerinsav-3- foszfát redukciójára. - regeneráció. a ribulóz-1,5-biszfoszfát-karboxiláz regenerálása: 6 glicerinsav-3-foszfát 3 ribulóz-1,5-biszfoszfát + 1 trióz-foszfát. Nyereség ciklusonként egy trióz-foszfát!")
21
A Calvin-ciklus jelentősége
Három pentóz-foszfátból 3 molekula széndioxid megkötésével 6 trióz-foszfát keletkezik A három pentóz-foszfát regenerálódik A 3 széndioxid molekulából a nettó eredmény 1 exportálódó trióz-foszfát Ez cukrok (szacharóz), zsírsavak, aminosavak szintézisére szolgál A folyamat során 9 ATP és 6 NADPH használódik fel
, zsírsavak, aminosavak szintézisére szolgál. A folyamat során 9 ATP és 6 NADPH használódik fel.")
22
A Calvin-ciklus regulációja
A fotoszintetikus elektrontranszportlánc működését elsősorban a NADP+, az ADP és Pi mennyisége szabja meg. Sötétben: a redukált NADPH mennyiség 5-20%, ATP/ADP arány 0,1-0,2; sztróma pH = 7 Fényen: a redukált NADPH mennyiség 40-50%, ATP/ADP arány 1-5, sztróma pH = 8-8,5 (enzim optimum) Fényen, CO2 hiányában: a redukált NADPH mennyiség 90% A Calvin-ciklus működését szabályozza: 1, A trióz-foszfát/ortofoszfát transzlokátor (később) 2, A fény Fényre aktiválódó enzimek: Rubisco-aktiváz NADP:gliceraldehid-3-foszfát dehidrogenáz Fruktóz-1,6-biszfoszfát foszfatáz Szedoheptulóz-1,7-biszfoszfát foszfatáz Ribulóz-5-foszfát kináz (foszforibulo-kináz)
 Fényen, CO2 hiányában: a redukált NADPH mennyiség 90% A Calvin-ciklus működését szabályozza: 1, A trióz-foszfát/ortofoszfát transzlokátor (később) 2, A fény. Fényre aktiválódó enzimek: Rubisco-aktiváz. NADP:gliceraldehid-3-foszfát dehidrogenáz. Fruktóz-1,6-biszfoszfát foszfatáz. Szedoheptulóz-1,7-biszfoszfát foszfatáz. Ribulóz-5-foszfát kináz (foszforibulo-kináz)")
23
A Calvin-ciklus számos enzimét a fényfüggő ferredoxin-tioredoxin rendszer aktiválja
Fény PS I a célenzimek diszulfid-csoportjainak redukálása szulfhidrillé a redukált tioredoxin fehérje segítségével. Így kerülnek a célenzimek aktív állapotba

24
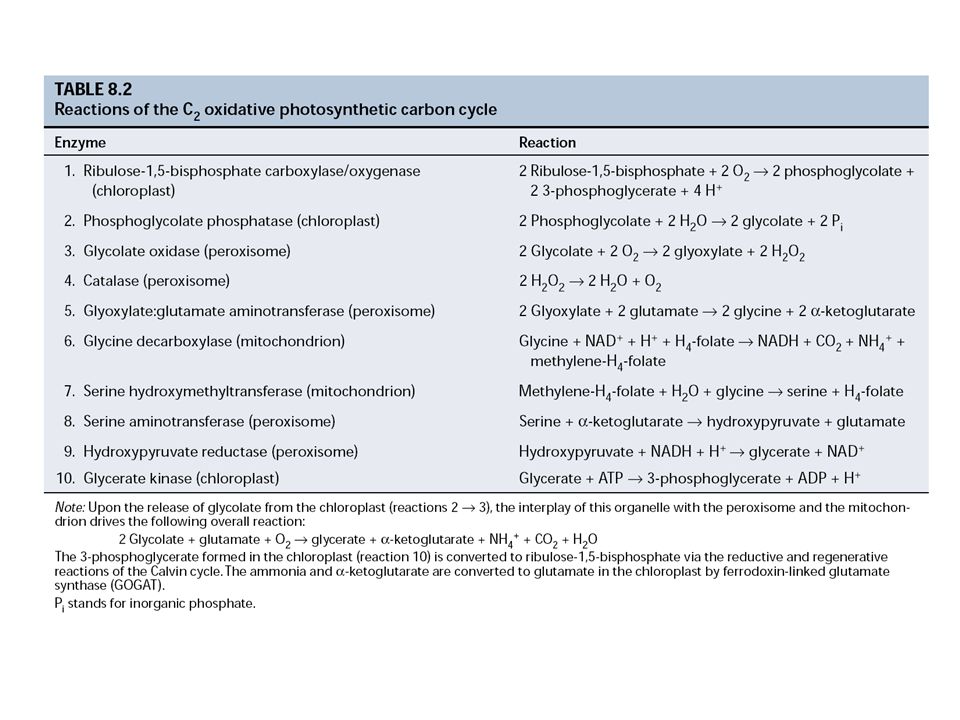
2. A C2-es fotorespirációs (karbon oxidációs, glikolát) ciklus
A Rubisco kettős aktivitásának következménye… Magas hőmérséklet oldott CO2/O2 arány csökken

25
2. A C2-es fotorespirációs (karbon oxidációs, glikolát) ciklus
- a Rubisco oxigenáz aktivitása következtében ("parazita") - CO2/O2 koncentrációk, hőmérséklet-függő - a kloroplasztisz, a peroxiszóma és a mitokondrium közreműködésével jön létre a ciklus
 - CO2/O2 koncentrációk, hőmérséklet-függő. - a kloroplasztisz, a peroxiszóma és a mitokondrium közreműködésével jön létre a ciklus.")
26
A) A kloroplasztiszban:
Ru-1,5-BP + O2 glikolsav-foszfát + glicerinsav-3-foszfát glikolsav-foszfát + H2O glikolsav + Pi B) A peroxiszómában (1): glikolsav + O2 glioxálsav + H2O2 (glikolsav oxidáz) (a glioxálsav visszajuthat a kloroplasztiszba, ahol NADPH felhasználásával újra glikolsavvá alakulhat, vagy glioxálsav + glutaminsav glicin + α-ketoglutársav (transzamináció) C) A mitokondriumban: glicin + NAD+ + H4-folát NADH + CO2 + NH4+ + metilén H4-folát metilén H4-folát + H2O + glicin szerin + H4-folát D) A szerin visszajut a peroxiszómába: szerin + α-ketoglutársav hidroxi-piruvát + glutaminsav hidroxi-piruvát + NADH + H+ glicerinsav + NAD+ E) A kloroplasztiszban: glicerinsav + ATP glicerinsav-3-foszfát + ADP + H+
 A peroxiszómában (1): glikolsav + O2 glioxálsav + H2O2 (glikolsav oxidáz) (a glioxálsav visszajuthat a kloroplasztiszba, ahol NADPH felhasználásával újra glikolsavvá alakulhat, vagy. glioxálsav + glutaminsav glicin + α-ketoglutársav (transzamináció) C) A mitokondriumban: glicin + NAD+ + H4-folát NADH + CO2 + NH4+ + metilén H4-folát. metilén H4-folát + H2O + glicin szerin + H4-folát. D) A szerin visszajut a peroxiszómába: szerin + α-ketoglutársav hidroxi-piruvát + glutaminsav. hidroxi-piruvát + NADH + H+ glicerinsav + NAD+ E) A kloroplasztiszban: glicerinsav + ATP glicerinsav-3-foszfát + ADP + H+")
27
GOGAT ATP

28
2. A C2-es fotorespirációs (karbon oxidációs, glikolát) ciklus
Összegezve: Minden 2 molekula glikolsav-foszfát (2 x 2 = 4 C atom), mely a Calvin-ciklusból a RuBP oxigenálódása miatt vész el, 1 molekula glicerinsav-3-foszfáttá (C3) + 1 molekula CO2-dá alakul. Más szóval, az oxigenálódás miatt elveszett szén 75%-át a fotorespirációs ciklus visszavezeti a Calvin-ciklusba. A fotorespirációs ciklussal viszont a CO2 fixáció teljes energiaigénye megnő. A C2-ciklusban 3 mól O2 felvétele 1 mól CO2 felszabadulást eredményez 2 mól ATP + 2 mól redukálószer felhasználása mellett.
, mely a Calvin-ciklusból a RuBP oxigenálódása miatt vész el, 1 molekula glicerinsav-3-foszfáttá (C3) + 1 molekula CO2-dá alakul. Más szóval, az oxigenálódás miatt elveszett szén 75%-át a fotorespirációs ciklus visszavezeti a Calvin-ciklusba. A fotorespirációs ciklussal viszont a CO2 fixáció teljes energiaigénye megnő. A C2-ciklusban 3 mól O2 felvétele 1 mól CO2 felszabadulást eredményez 2 mól ATP + 2 mól redukálószer felhasználása mellett.")
30
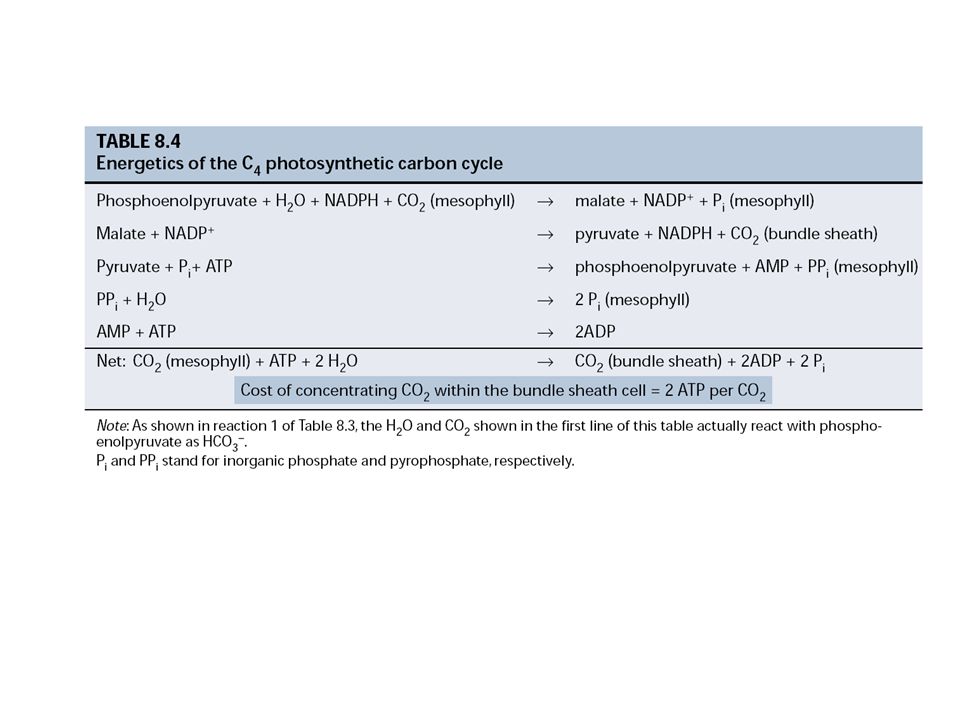
A CO2 fixáció C4-es útja (CO2 koncentráló mechanizmus)
Probléma: a hőmérséklet emelésével a CO2 és O2 oldhatósága megváltozik úgy, hogy a CO2/O2 arány csökken. Ezért a Rubisco oxigenáz aktivitása erősebbé válik. A sztómák is záródnak a vízvesztés megakadályozása miatt, így tovább csökken a rendelkezésre álló CO2 mennyisége. C4 stratégia C4-es növények jellegzetességei: A primer fixációs termékek 4 szénatomosak, pl. oxálecetsav, almasav és aszparaginsav A CO2 fixáció fényen történik A négyszénatomos molekulákból 1 szénatom adódik a C3-as ciklus felé Kétféle fotoszintetikus sejttípus van bennük (mezofill és hüvelyparenchima sejtek)
")
31
C3 és C4 levélszerkezet A C4-es növények anatómiája szembeszökően különbözik a C3-as növényekétől. Csak a C4-es növényekben található a hüvelyparenchima (bundle sheath) vagy Kranz sejtek. A széndioxid elsődleges kötését a PEP karboxiláz enzim katalizálja a mezofill sejtek citoplazmájában.
 vagy Kranz sejtek. A széndioxid elsődleges kötését a PEP karboxiláz enzim katalizálja a mezofill sejtek citoplazmájában.")
32
A kukoricalevél tipikus Kranz-anatómiája

33
C4 levél szerkezet és a C4 út
CO2 Mezofill sejt Hüvely- parenchima sejt Levélér (szállítószövet) A C4 növény levél fotoszintetizáló sejtek Sztóma C4 levél felépítés PEP karboxiláz Oxálecetsav (4 C) PEP (3 C) Almasav (4 C) ADP ATP Piruvát (3 C) CALVIN CIKLUS Cukor Szállítószövet primér fixációs termék C4 sav (oxálecetsav) foszfoenol-piruvát (PEP) karboxilálódik a C4-es és a C3 ciklus térben elválasztódik: speciális anatómia (mezofill és hüvelyparenchima) trópusi, szubtrópusi növényekben
 A C4 növény levél fotoszintetizáló sejtek. Sztóma. C4 levél felépítés. PEP karboxiláz. Oxálecetsav (4 C) PEP (3 C) Almasav (4 C) ADP. ATP. Piruvát (3 C) CALVIN. CIKLUS. Cukor. Szállítószövet. primér fixációs termék C4 sav (oxálecetsav) foszfoenol-piruvát (PEP) karboxilálódik. a C4-es és a C3 ciklus térben elválasztódik: speciális anatómia (mezofill és hüvelyparenchima) trópusi, szubtrópusi növényekben.")
34
A foszfoenol-piruvát karboxiláz (PEP)… rendkívül nagy affinitással végzi a karboxilációs reakciót
… rendkívül nagy affinitással végzi a karboxilációs reakciót")
35
A C4 alapciklus lépései:
1) PEP karboxiláció a MEZOFILL sejtek citoplazmájában oxálecetsav malát, aszpartát 2) A C4 sav transzportja a HÜVELYPARENCHIMA-ba 3) A C4 sav (almasav, aszparaginsav) dekarboxilációja a CO2 fixációja a C3-as ciklusban 4) A C3 sav (piruvát, alanin) transzportja MEZOFILL
 PEP karboxiláció a MEZOFILL sejtek citoplazmájában. oxálecetsav malát, aszpartát. 2) A C4 sav transzportja a HÜVELYPARENCHIMA-ba. 3) A C4 sav (almasav, aszparaginsav) dekarboxilációja. a CO2 fixációja a C3-as ciklusban. 4) A C3 sav (piruvát, alanin) transzportja MEZOFILL.")
36
NADP-almasav-enzim (NAPD-ME) típus
 típus")
39
A CO2 fixáció C4-es útja (CO2 koncentráló mechanizmus)
A CO2 koncentrálódása 8-10-szeres a C3-as levélhez képest. Energiamérleg: ellentétben a C3-as növényekkel, nem 3 ATP és 2 NADPH, hanem 5 ATP és 2 NADPH / 1 CO2 a CO2 koncentráláshoz több energia szükséges, melynek nyeresége, hogy nincs fotorespiráció Fény-regulált kulcsenzimek: PEP karboxiláz NADP-almasav dehidrogenáz (tioredoxin) Piruvát-ortofoszfát-dikináz
 Piruvát-ortofoszfát-dikináz.")
40
A dekarboxilációs mechanizmus alapján 3 típus
i) NADP-almasav enzim típus (NADP-ME, kloroplasztisz) ii) Foszfoenol-piruvát-karboxikináz típus (PEP-CK, citoplazma) iii) NAD-almasav enzim típus (NAD-ME, mitokondrium)
 NADP-almasav enzim típus. (NADP-ME, kloroplasztisz) ii) Foszfoenol-piruvát-karboxikináz típus (PEP-CK, citoplazma) iii) NAD-almasav enzim típus (NAD-ME, mitokondrium)")
41
A kloroplasztiszok szerkezete és elhelyezkedése a különböző fixációs típusú hüvelyparenchima sejtekben Klp centrifugálisan, nincsenek gránumok (nincs lineáris e-trp), mert az almasav a CO2-on kívül redukáló erőt (NADPH-t) is biztosít Az ATP előállításához elegendő a ciklikus fotofoszforilációs aktivitás Pl. Zea mays - kukorica Vannak gránumok, klp centrifugálisan Pl. Panicum maximum - pázsitkóró Vannak gránumok, klp centripetálisan Pl. Amaranthus retroflexus – szőrös disznóparéj, Panicum miliaceum - köles
, mert az almasav a CO2-on kívül redukáló erőt (NADPH-t) is biztosít. Az ATP előállításához elegendő a ciklikus fotofoszforilációs aktivitás. Pl. Zea mays - kukorica. Vannak gránumok, klp centrifugálisan. Pl. Panicum maximum - pázsitkóró. Vannak gránumok, klp centripetálisan. Pl. Amaranthus retroflexus – szőrös disznóparéj, Panicum miliaceum - köles.")
42
4. A CO2 fixáció C4-es útja a CAM növényekben
Crassulacean Acid Metabolism Crassulaceae Cactaceae Euphorbiaceae Liliaceae Bromeliaceae…

43
A CAM útvonal hasonló a C4 úthoz
A lépések térbeli elkülönülése. A C4 növényekben, a szén fixálása és a Calvin ciklus eltérő sejtekben zajlik. (a) A lépések időbeli elkülönülése. A CAM növényekben, a szén fixálása és a Calvin ciklus ugyanabban a sejtben de eltérő időben zajlik. (b) Ananász Cukornád Hüvelyparenchima sejt Mezofill sejt Szerves sav CALVIN CIKLUS Cukor CO2 C4 CAM CO2 négy szénatomos savakba épül (szén fixálás) Éjszaka Nappal 1 2 A szerves savak- ból CO2 szabadul fel a Calvin ciklus számára
 A lépések időbeli elkülönülése. A CAM növényekben, a szén fixálása és a Calvin ciklus ugyanabban a sejtben de eltérő időben zajlik. (b) Ananász. Cukornád. Hüvelyparenchima sejt. Mezofill sejt. Szerves sav. CALVIN CIKLUS. Cukor. CO2. C4. CAM. CO2 négy szénatomos savakba épül. (szén fixálás) Éjszaka. Nappal A szerves savak- ból CO2 szabadul fel a Calvin ciklus számára.")
44
4. A CO2 fixáció C4-es útja a CAM növényekben
karboxiláció /dekarboxiláció időben elválasztva - sivatagi növények Éjjel: sztómák nyitva PEP + CO2 oxálecetsav almasav vakuólum ("sötét savanyodás") Nappal: zárt sztómák dekarboxiláció, CO2 Calvin ciklus C3 savak trióz-P keményítő, szacharóz CAM növényekben a PEP-karboxiláz kettős funkciójú éjjel: karboxiláz (foszforilált forma) nappal: dekarboxiláz (defoszforilált, almasav-gátolt)
 Nappal: zárt sztómák. dekarboxiláció, CO2 Calvin ciklus. C3 savak trióz-P keményítő, szacharóz. CAM növényekben a PEP-karboxiláz kettős funkciójú. éjjel: karboxiláz (foszforilált forma) nappal: dekarboxiláz (defoszforilált, almasav-gátolt)")
45
A CO2 fixáció C4-es útja a CAM növényekben

46
A PEP-karboxiláz nappali inaktív, és éjszakai aktív formája

47
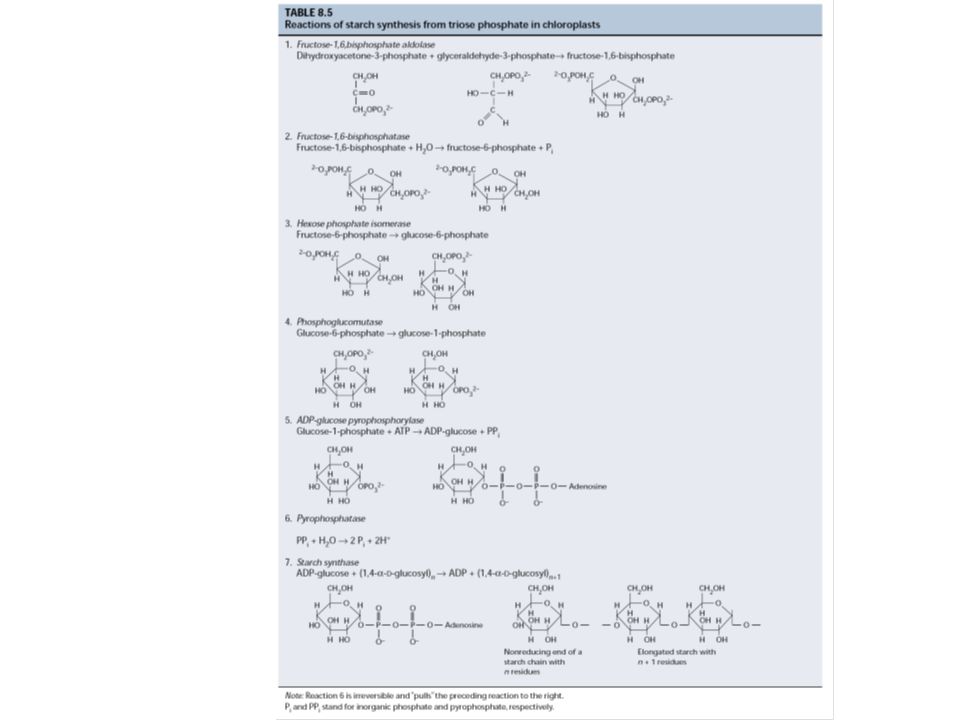
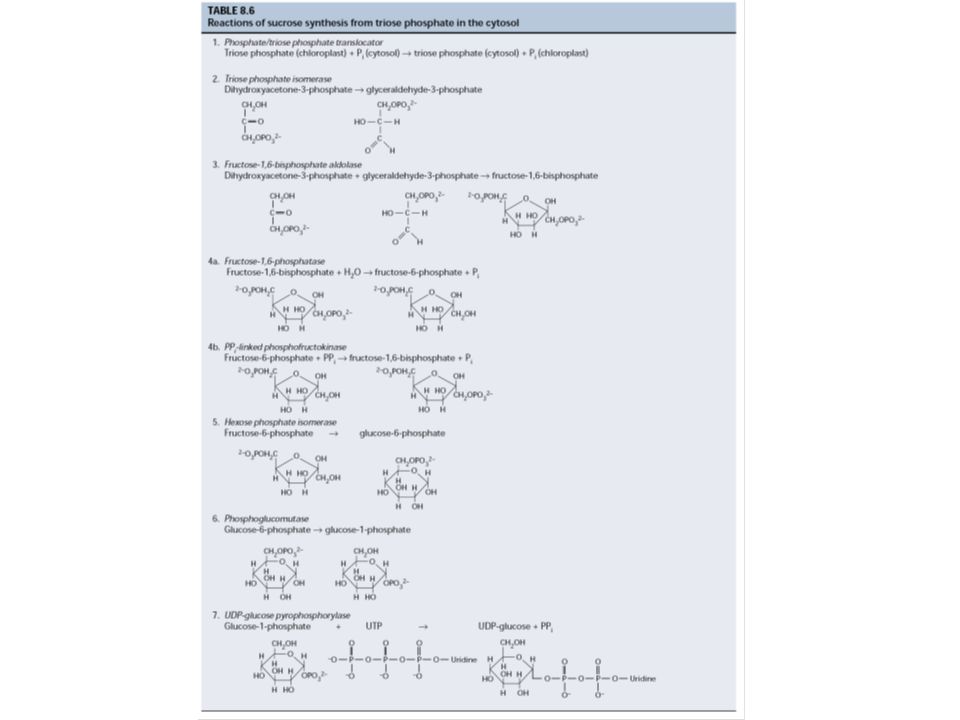
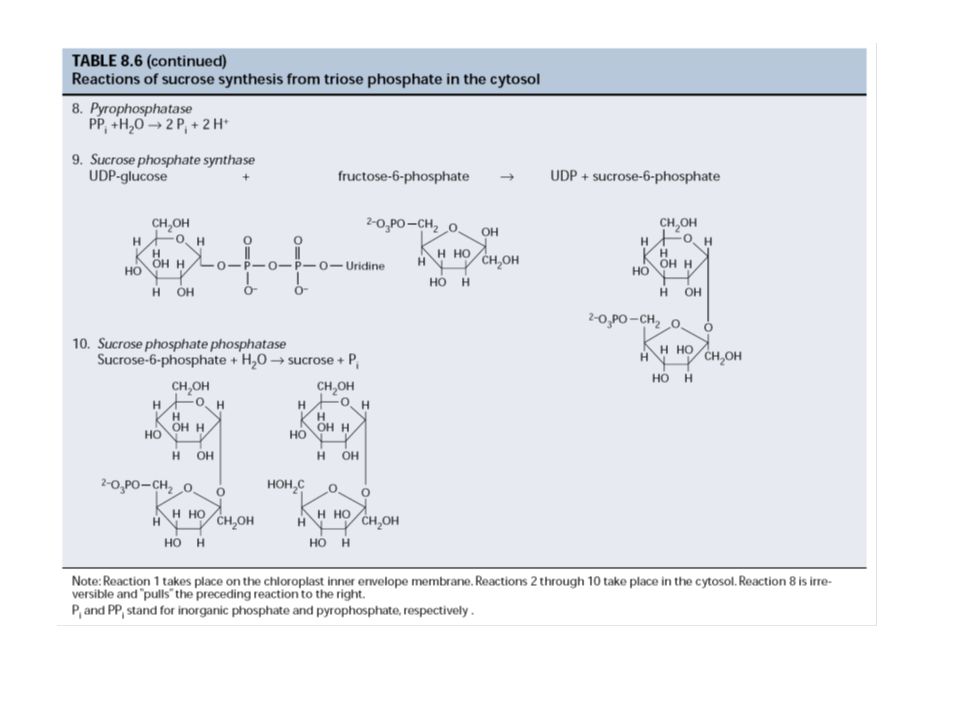
5. A szaharóz és a keményítő szintézise

48
Fr2,6-biszfoszfát gátlás

49
5. A szaharóz és a keményítő szintézise
- keményítő: a kloroplasztiszban szaharóz: a citoplazmában közös lépések a trióz-P-tól a glukóz-1-P-ig izoenzimek: pl. fruktóz-1,6-biszfoszfát foszfatáz kloroplasztiszban: tioredoxin citoszolban: fruktóz-2,6-biszfoszfát keményítő: ADP-glükóz szaharóz: UDP-glükóz Trióz-P megoszlás: trióz-P/Pi transzlokátor (antiport) Pi reguláló szerep Regulátor a citoszolban: fruktóz-2,6-biszfoszfát Fr-1,6-BP Fr-6-P átalakulásnál (, inhibitor; , aktivátor)
 Pi reguláló szerep. Regulátor a citoszolban: fruktóz-2,6-biszfoszfát. Fr-1,6-BP Fr-6-P átalakulásnál (, inhibitor; , aktivátor)")
50
A szaharóz és a keményítő szintézise egymással versengő folyamatok

51
A cukorszintézis regulátora: a fruktóz-2,6-biszfoszfát