Előadást letölteni
Az előadás letöltése folymat van. Kérjük, várjon

1
Centrális dogma és a bioinformatika főbb területei a molekuláris biológiában
DNS Gén transzkripció, RNS szerkesztés RNS transzkriptomika degradáció transzláció, poszttranszlációs módosítás proteomika fehérje degradáció biokémiai aktivitás metabolikus útvonalak metabolomika

2
GENOMIKA A BIOLÓGIAI INFORMÁCIÓ HORDOZÓ MEGFEJTÉSE
A teljes genetikai állomány szekvenciájának meghatározása, A szekvenciákon elhelyezkedő funkcionális régiók számítógépes jóslása: annotálás

3
Funkcionális genomika RNS szinten
TRANSZKIPTOMIKA

4
Egy DNS chip kísérlet folyamatábrája

5
A chipek kiértékelése, eredménye

6
Funkcionális genomika fehérje szinten
PROTEOMIKA

7
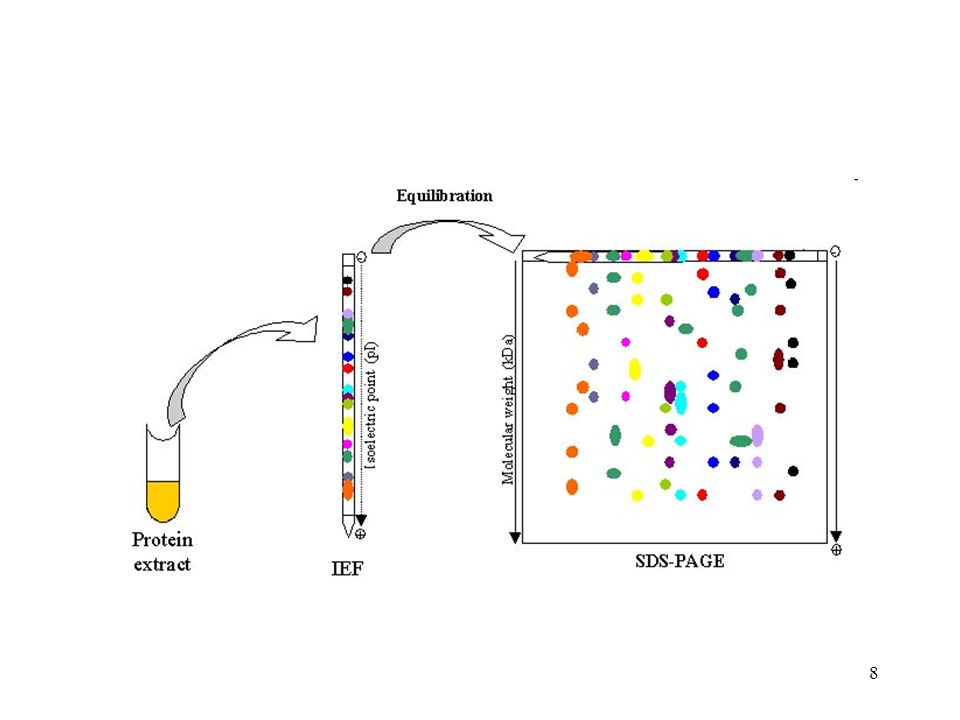
Proteomika Izoelektromos fókuszálás SDS PAGE Minta elő
EgyTipikus protokol Izoelektromos fókuszálás SDS PAGE Minta elő Protein azonosítás tömegspektrometria Láthatóvá tétel Protein pötty kivágás Kép analízis

9
Proteomika: az elválasztástól az azonosításig

10
Oligonukleotid szintézis
Mintegy 50 éves múlt 5 különböző kémia: foszfáttriészter, fosztittriészter, foszfátdiészter Foszforamidit, H-foszfonát Szilárd fázisú szintézis: hordozó: controlled pore glass (CPG), vagy polisztirol Mind DNS mind RNS szintetizálható mind a két módszerrel
, vagy polisztirol. Mind DNS mind RNS szintetizálható mind a két módszerrel.")
11
Oligonukleotid szintézis: monomerek

12
Oligonukleotid szintézis:foszforamidit módszer
B B 1 1 O H hordozó B 1 O O O O di-MeO Tr A c O O 2% DCA/DCM O O di-MeO Tr hordozó hordozó < 1 % Ac 2 O O H hordozó B 1 B 2 O O tetrazol di-MeO Tr O N újabb ciklus O P N N C O > 99 % B 2 B 2 O O O di-MeO Tr O O I 2 / H 2 O B di-MeO Tr O 1 B O 1 O O O O O P O O N C P O O O N C O O hordozó hordozó

13
Néhány automata szintetizátor

14
OLIGONUKLEOTIDOK UTÓKEZELÉSE I.
- Deblokkolás: védőcsoportok eltávolítása, hordozóról való lehasítás A lúg hatására lehasad: - védőcsoportok a bázisokról - -cianoetil csoport a fosztátról - oligo a hordozóról - liofilezés, sómentesítés

15
OLIGONUKLEOTIDOK UTÓKEZELÉSE II.
- méret szerinti elválasztás, PAGE, vagy HPLC (sokszor nem szükséges) Ioncserés kromatográfia Hidrofób kromatográfia a DMT csoporton keresztül - kvantitálás bázis e(µmol * cm/cm3) dA 15.4 dG 11.7 dC 7.5 dT 8.8 átlagosan e = 10 (µmol * cm)/cm3
 Ioncserés kromatográfia. Hidrofób kromatográfia a DMT csoporton keresztül. - kvantitálás. bázis. e(µmol * cm/cm3) dA dG dC dT átlagosan e = 10 (µmol * cm)/cm3.")
16
OLIGONUKLEOTIDOK: FELHASZNÁLÁSI TERÜLETEK
- primerek: DNS, RNS szekvenálás, PCR, mutagenezis - linkerek: mesterséges hasítóhelyek bevitele - adapterek: különböző (nem-kompatibilis) ragadós DNS végek összekötése, esetleg hasítóhely bevitelével - hibridizációs próbák, DNS diagnosztika - antiszensz oligonukleotidok, génterápia - gén darabok - egyéb, pl. DNS affinitásoszlopok készítése, stb.
 ragadós DNS végek összekötése, esetleg hasítóhely bevitelével. - hibridizációs próbák, DNS diagnosztika. - antiszensz oligonukleotidok, génterápia. - gén darabok. - egyéb, pl. DNS affinitásoszlopok készítése, stb.")
17
egyik vég ragadós, a másik tompa.
Linkerek rövid, önkomplemeter oligonukleotidok, amelyek saját magukhoz hibridizálva olyan tompa végű, kettősszálú DNS-t képeznek, amely tartalmazza egy tetszőleges restrikciós endukleáz felismerő helyét: Restrikciós hely bevitelére alkalmas BamHI CGGATCCG EcoRI GGAATTCC PstI GCTGCAGC ADAPTEREK szintetikus kettősszálú oligonukleotidok, amelyek végei kompatibilisek különböző restrikciós endukleázokkal végzett emésztések során képződő végekkel. Esetenként valamilyen restrikciós endonukleáz felismerőhelyét tartalmazhatja. Hosszú adapterek: két ragadós vég között egy harmadik restrikiós endonukleáz felismerõhelye Rövid adapterek: egyik vég ragadós, a másik tompa. EcoRI pl. BamHI (ApaI) SacI EcoRI – SmaI...5'AATTCCCGGG 3‘ ’GGGCCC 5’ 5' GATCCGGGCCCTGTTAGAGCT 3' 3' GCCCGGGACAATC 5' SmaI
 SacI. EcoRI – SmaI...5 AATTCCCGGG 3‘ 3’GGGCCC 5’ 5 GATCCGGGCCCTGTTAGAGCT 3 3 GCCCGGGACAATC 5 SmaI.")
18
Primerek szintetikus oligodeoxinukleotidok, amelyek iniciációs pontjaiként szolgálnak a templát függő DNS polimerázok számára. Univerzális primerek: a gyakorlatban elterjedt vektorok klónozó helyének környékére tervezett primerek. Specifikus primerek: az adott feladatnak megfelelően szintetizált speciális szekvenciájú oligonukleotidok.

19
Polimeráz láncreakció I.
Termostabil DNS polimeráz, Taq, Pfu, Vent, Pwo stb. Soknak nincs 3’ 5’ exonukleáz aktivitása “A” túlnyúló

20
Polimeráz láncreakció II.

21
A POLIMERÁZ LÁNC REAKCIÓ MÓDOZATAI
- belső PCR, specificitás növelése belső primerek használatával egy elsődleges PCR terméken - egy specifikus primert használó PCR + primer adapter limitált információ esetén - LM PCR, ligálás közvetített PCR, kromoszóma metiláltsági térképezésére - inverz PCR, szegélyező szekvenciák izolálására - “farkazott PCR” ld. cDNS izolálás - RT PCR, reverz transzkripció kapcsolt PCR, ld. cDNS - kvantitatív PCR, mRNS mennyiségének becslésére

22
A POLIMERÁZ LÁNC REAKCIÓ FELHASZNÁLÁSI TERÜLETEI
klónozás, megfelelő DNS szakaszok kinyerése, esetenként degenerált primerekkel DNS szekvenálás, pl. ThermoSequenase mutagenezis szálspecifikus próbák előállítása diagnosztika fertőzések kimutatására mutáns allélek kimutása

23
BELSŐ (NESTED) PCR 1. PCR 2. PCR 2 primer párt használunk nagyobb specificitás, a nemspecifikus termékek eltűnése költségesebb

24
INVERZ PCR A géneket környező régiók kiamplifikálkására
Általában a gén szekvenciája részben vagy teljesen ismert primer pár tervezhető A hasítás 'A' restrikciós enzimmel önligálás PCR 'A'

25
REVERZ TRANSZKRIPCIÓ KAPCSOLT PCR
(RT-PCR) mRNS antiszensz primer reverz transzkripció reverz transzkriptázzal AMV: avian mieloblastosis virus RT MMLV: Moloney murine leukémia vírus RT Tth: Thermus thermophilus, Mn2+ jelenlétében cDNS szensz primer PCR Gének szerveződésének vizsgálatára 250 500 750 1000 RT+ RT- gK bp
 mRNS. antiszensz primer. reverz transzkripció reverz transzkriptázzal. AMV: avian mieloblastosis virus RT. MMLV: Moloney murine leukémia vírus RT. Tth: Thermus thermophilus, Mn2+ jelenlétében. cDNS. szensz primer. PCR. Gének szerveződésének vizsgálatára RT+ RT- gK. bp.")
26
REVERZ TRANSZKRIPCIÓ KAPCSOLT KVANTITATÍV PCR
Az RNS preparátumnak DNS mentesnek kell lennie exponenciális rész lineráris szakasz plató ciklus szám termék Hagyományos PCR-rel igen nehéz a termék mennyiségének a meghatározása

27
Real-time PCR T : SYBRGreenI : fluorescens SYBRGreenI Denaturálás
Hibiridizáció DNA szintézis Real-time PCR : SYBRGreenI : fluorescens SYBRGreenI

28
Real-time PCR reakció analízise
Kalibráció ismert mennyiségű genomiális DNS-sel reprodukálhatóság ismeretlen mennyiségű templát cDNA-sel. 280ng 28ng 2.8ng 0.28ng 0.028ng gDNA:

29
A POLIMERÁZ LÁNC REAKCIÓ FELHASZNÁLÁSI TERÜLETEI: klónozás

30
A POLIMERÁZ LÁNC REAKCIÓ FELHASZNÁLÁSI TERÜLETEI: mutagenezis
random: a Taq polimeráz átírási hűsége rosszabb, hibák épülnek be és amplifikálódnak 1-2 hiba 1 kb hosszon Mn2+ vagy nukleotid analógok (inozin) növelik a mutációs rátát in vitro evolúció irányított: O1 Mf BamHI XhoI Mr O2 O1-Mr 1st PCR-s Mf –O2 O1 BamHI XhoI CO2 2nd PCR O1 – O2 XhoI BamHI digest with XhoI and BamHI ligate into XhoI – BamHI digested vector
 növelik a mutációs rátát. in vitro evolúció. irányított: O1. Mf. BamHI. XhoI. Mr. O2. O1-Mr. 1st PCR-s. Mf –O2. O1. BamHI. XhoI. CO2. 2nd PCR. O1 – O2. XhoI. BamHI. digest with XhoI and BamHI. ligate into XhoI – BamHI digested vector.")
31
A PCR DIAGNOSZTIKAI ALKALMAZÁSAI I.
- FERTŐZÉSEK KIMUTATÁSA minden élőlény genetikai anyaga nukleinsav, ez tartalmaz specifikus szekvenciaelemeket speciálisan tervezett primerek segítségével PCR a termék megjelenése fertőzésre utal

32
A PCR DIAGNOSZTIKAI ALKALMAZÁSAI II. MUTÁCIÓK KIMUTATÁSA
normális C gén mutáns 5' 3' A C 5' 3' PCR egészséges nincs termék beteg

33
Ligálás és a PCR kombinálása

34
GÉNSZINTÉZIS STRATÉGIÁK
1. Szintetikus DNS fragmentek összeállítása átfedő oligomerekből 1. fragment 2. fragment 1+2. fragment 3. fragment 1+2. fragment fragment hátrány: drága, mind a két szálat meg kell szintetizálni

35
2.a. Hajtű módszer 2. Fragment összeállítás primer-templát módszerrel
3’ 5’ 5’ 3’ Klenow, dNTP 5’ 3’ 3’ 5’ Több verzió is erre az enzimatikus feltöltési elvre alapszik. 2.a. Hajtű módszer 5’ 3’ Klenow, dNTP emésztés restrikciós endonuklázokkal ligálás

36
2. c. Fragment összeállítás a javító mechanizmus (gap repair) kihasználásával
Híd ligálás 5’ 3’ 5’ 3’ 3’ 5’ átfedő szintetikus oligonukleotidok hasított vektor hibridizálás, enzimatikus feltöltés hasított vektor 5’ 3’ 3’ 5’ polimerizáció Klenow, dNTP ligálás közvetlen transzformálás transzformálás

37
Génexpresszió szabályozása szintetikus oligonukleotidokkal
sejtmembrán fehérjék mRNS sejtmag Kromoszómális DNS oligonukleotid citolplazma Cél molekula: fehérje mRNS DNS oligonukleotid iránya a leolvasással megegyezõ v. kettõsszálú a leolvasással ellentétes a leolvasással megegyezõ RNS DNS duplex triplex köcsönhatás fehérje-DNS RNS-DNS hibrid/ RNáz H Hoogsteen féle triplex Gátlás transzkripciós szabályozás transzláció transzkripció

38
Triplex képződésének kölcsönhatásai
homopurin homopirimidin szekvenciáknál

39
Antiszensz oligonukleotidok lehetséges célszekvenciái a mRNS-en
exon-intron határhoz, slicing gátlás Az 5’ sapka régióhoz Transzlációs iniciációs régióhoz Génen belüli régióhoz Kapcsolódó oligonukleotidok Elsődleges hatásmechanizmus: Az endogén RNázH leemészti a DNS:RNS hibrid RNS szálát

40
Oligonukleotid szintézis:foszforamidit módszer
B B 1 1 O H hordozó B 1 O O O O di-MeO Tr A c O O 2% DCA/DCM O O di-MeO Tr hordozó hordozó < 1 % Ac 2 O O H hordozó B 1 B 2 O O tetrazol di-MeO Tr O N újabb ciklus O P N N C O > 99 % B 2 B 2 O O O di-MeO Tr O O I 2 / CS2 B di-MeO Tr O 1 B O 1 O O O O O P O S N C P O O O N C O O hordozó hordozó

41
PEPTID NUKLEINSAVAK (PNA)
Nincs cukorfoszfát gerinc, ehelyett peptid kötéssel összekapcsolt lánc van Specifikus Stabil PNA T10-Lys TTTTTTTTTT-Lys PNA SV40-Lys ATTTTCTTCATTTTTTCTTC-Lys Ma már külön könyv van a PNA- alkalmazásairól

42
MUTAGENEZIS - in vivo gének elrontására,vagy módosítására
Random mutagenezis találomra létrehozott mutációk mutagén anyagok: UV, kemikáliák, radioaktív sugárzás PCR: a hőstabil polimerázok átírási hűsége rosszabb mutagén törzsek transzpozon mutagenezis Irányított mutagenezis Adott helyen - deléciók inszerciók léterhozása - pont mutációk létrehozása

43
Gének irányított szétroncsolása: interpozon mutagenezis
Ar1 oriV oriT oriV: szűk gazdaspecificitás Ar2 Ar: antibiotikum rezisztencia Ar2 oriT ori Ar1 Ar2 vad típus mutáns poláris hatás

44
Deléciós mutagenezis a leolvasási keret sértése nélül
Ar1 oriV oriT oriV: szűk gazdaspecificitás Ar: antibiotikum rezisztencia oriT ori Ar1 vad típus mutáns poláris hatás ?

45
QUICK CHANGE MUTAGENEZIS (STRATAGENE)
")
46
FEHÉRJE TERMELTETÉS VAN GÉNÜNK: Legyen az prokarióta vagy eukarióta
természetes vagy szintetikus vad típusú vagy mutáns

47
FEHÉRJE TERMELTETŐ RENDSZEREK
PROKARIÓTÁK E. coli ismert, Bármilyen fehérje, feltéve, ha - nem túl nagy - nem túl kicsi - nem túl hidrofób - nincs túl sok cisztein benne szekréciós rendszere minimális Bacillus subtilis jól ismert, ha nem is annyira, mint az E. coli van szekréciós rendszere EUKARIÓTÁK emlős sejtvonalak minden fajta fehérje termeltethető bennük tranziens expressziós rendszerek viszonylag gyors, de korlátozott lehetőségek, stabil expressziós rendszerek hosszú, fáradságos optimalizálást igényel élesztő gyorsan szaporodó eukarióta sejtvonal, sok ismeretanyag viszonylag könnyű kezelhetőség poszttranszlációs modifikációk lehetősége

48
Escherichia coli ELŐNYÖK óriási mennyiségű ismeretanyag
könnyű kezelhetőség, gyors növekedési sebesség, viszonylag olcsó médium nagy mennyiségű biomassza gazdaságos előállítása ismert szelekciós rendszerek legtöbb expressziós rendszer HÁTRÁNYOK általában nem szekretál a sejten belül redukáló atmoszféra diszulfid hidak kialakulása gátolt az eukarióta poszttranszlációs módosítások általában nem megvalósíthatóak nem alakul ki a megfelelő aktiv szerkezet, rossz folding

49
STRATÉGIÁK TÚLTERMELTETÉSRE
KONSTITUTÍV PROMÓTER INDUKÁLT TERMELTETÉS a fehérje a teljes növekedési fázis alatt expresszálódik nagy sejttömeg elérése után a fehérje visszanyerése nem igazán használatos, toxicitási problémák miatt a promoter represszált állapotban van a növekedés egy bizonyos fázisáig valamilyen indukcióval derepresszió további növesztés fehérje visszanyerése legáltalánosabban használt stratégia toxicitás sok esetben megoldható

50
PROKARIÓTA EXPRESSZIÓS VEKTOR ELEMEI
SZF TSZE GS Pr SD kódoló szekvencia TT -35 -10 STOP kodon UAAU UGA UAG TTGACAN17TATAAT START kodon AUG GUG UUG 5’ UAAGGAGGN(3-11) AR ORI Pr: promóter TT: transzkripciós terminációs szignál, SZF: szabályozó fehérje,, TSZE: transzlációt szabályozó elemek, SD: Shine-Dalgarno szekvencia, AR: antibiotikum rezisztencia, ORI: replikációs origo GS: gazdasejt
 AR. ORI. Pr: promóter. TT: transzkripciós terminációs szignál, SZF: szabályozó fehérje,, TSZE: transzlációt szabályozó elemek, SD: Shine-Dalgarno szekvencia, AR: antibiotikum rezisztencia, ORI: replikációs origo. GS: gazdasejt.")
51
A génexpresszió szabályozása
transzkripció transzláció DNS RNS FEHÉRJE transzkripciós szinten poszt-transzkripciós szinten (mRNS degradáció, protein stabilitás, stb.)
")
52
Különböző faktorok relatív hatása a fehérje termeltetésre E. coli-ban
Promóter indukálhatóság kb x Gazdasejt kb x A mRNS 5’ struktúrája kb. 100 x Transzláció kb. 100 x növekedési sebesség kb. 50 x (mRNS stabilitás)
")
53
TRANSZKRIPCIÓ Iniciáció fehérje termeltetés során a legfontosabb
Elongáció mivel a gén belsejében van, nem manipulálható Termináció a transzkripció befejezése, a transzkripciós apparátus túlterhelésének elkerülésére

54
A transzkripció iniciációja prokariótákban
sigma faktor holoenzim ? /` NTD CTD RNS polimeráz RNS polimeráz alegységek: , `, , , promóter 5’ 3’ 3’ 5’ k1 k-1 Ha k1 >> k-1, akkor a polimeráz holoenzim nagyon erősen kötődik a promóter szekvenciához, a polimeráz nehezen tudja elindítani a polimerizációs reakciót. Tehát a transzkript mennyisége csökken, habár in vitro a promóter és a holoenzim közötti kapcsolat erős. 5’ 3’ k2 k-2 5’ 3’ 3’ 5’ 5’

55
70 promoter szekvencia: -10 and -35 régiók

56
Bakteriális faktorok

57
Különböző gének expresszióját különböző szigma faktorok befolyásolják
tápelvonás, Stressz, Vasra éheztetés, sejtsűrűség, Felületi adházió Transzkripciós faktorok (Szigma) gén expresszió változás Morfológiai & fiziológiás változások Adaptáció / Védekezés Upon entry into stationary phase, bacteria undergo morphological and physiological changes in order to adapt and prevent cell damaging. These adaptations are accompanied by a switch in gene expression resulting form the action of different transcription factors. Among these factors involved in transcription regulation in stationary phase, there is sigmaS, an alternative subunit of the bacterial RNAP which accumulates in stressed cells or at the onset of stationary phase.
 gén expresszió változás. Morfológiai & fiziológiás változások. Adaptáció / Védekezés. Upon entry into stationary phase, bacteria undergo morphological and physiological changes in order to adapt and prevent cell damaging. These adaptations are accompanied by a switch in gene expression resulting form the action of different transcription factors. Among these factors involved in transcription regulation in stationary phase, there is sigmaS, an alternative subunit of the bacterial RNAP which accumulates in stressed cells or at the onset of stationary phase.")
58
Két faktor felépítése

59
riporter fehérje aktivitás Western blot fehérje detektálása
A génexpresszió tanulmányozása különböző módszerekkel különböző szinteken fehérje érése transzláció transzkripció Enzim aktivitás mérés mRNS detektálása Riportergén riporter fehérje aktivitás promóter aktivitás Western blot fehérje detektálása A vizsgált operon promóter

60
nem specifikusan kötődik
ELECTRO MOBILITY SHIFT ASSAY (EMSA) I. más néven gél retardációs assay a DNS-t, amely feltételezhetően fehérjét köt, megjelöljük jelölés lehet végen, szálon belül, egyszálon, kétszálon, kötési kísérletben a DNS kétszálú! a DNS ne legyen től hosszú bp, minél hosszabb a DNS, annál több nem specifikus reackió történik lehet tiszta fehérje vagy fehérje populáció Natív poliakrilamid gél -: fehérje nélkül, +: fehérjével összekeverve DNS jelölés DNS kötő fehérje nem kötődik kötődik összekeverés nem kötődik kötődik nem specifikusan kötődik
 I. más néven gél retardációs assay. a DNS-t, amely feltételezhetően fehérjét köt, megjelöljük. jelölés lehet végen, szálon belül, egyszálon, kétszálon, kötési kísérletben a DNS kétszálú! a DNS ne legyen től hosszú bp, minél hosszabb a DNS, annál több nem specifikus reackió történik. lehet tiszta fehérje vagy fehérje populáció. Natív poliakrilamid gél -: fehérje nélkül, +: fehérjével összekeverve. DNS jelölés. DNS kötő fehérje. nem kötődik. kötődik. összekeverés. - + nem kötődik. kötődik. nem specifikusan kötődik.")
61
ELECTRO MOBILITY SHIFT ASSAY (EMSA) II.
a mobilitás eltolódás inkább a komplex méretét jellemzi, a kölcsönhatás errősségét nem a kölcsönhatás erősségét a nem kötöt DNS, és a kötésben levő DNS jelének intenzitása jellemzi minél kevesebb fehérje okoz erős jelet az eltolódott sávban annál erősebb a kölcsönhatás SUPERSHIFT ASSAY titrálás fehérje mennyiségre ha van tippünk, hogy milyen fehérje kötődik és van rá specifikus ellenanyagunk hozzáadott fehérje mennyiség nő nem kötődik kötődik ellenanyag is kötődik a DNS - fehérje kölcsönhatás erősségéről ad felvilágosítást, nem a képződött mRNS mennyiségéről -: fehérje nélkül, +: csak fehérje hozzáadása, ++: fehérje és ellenanyag hozzáadása

62
DNázI FOOT PRINT I. DNS hossz 100 – 200 bp
DNS jelölés szigorúan a végén, szálspecifikusan mind a két jelölt szállal meg kell csinálni a kötési kísérletben a DNS kétszálú denaturáló poliakrilamid gélelektroforézis Maxam-Gilbert létra DNázI hasítás DNázI * * G A+G C+T C D *

63
hiperszenzitív hely (HSH)
DNázI FOOT PRINT II. DNázI hasítás -: fehérje nélkül +: fehérje jelenlétében Maxam-Gilbert létra * * G A+G C+T C D- D+ hiperszenzitív hely (HSH) DNázI HSH * *
 DNázI. HSH. * *")
64
UV KERESZTKÖTÉS (CROSS LINKING)
Ismeretlen DNS kötő fehérjék molekulatömegének becslésére 1. A DNS szálat szálon belül jelöljük, és Br-dUMP-t építünk be (a Br-dUMP nem zavarja a kötődést) minta marker : Br MS? SDS-PAGE jelölt fehérje DNázI kezelés EMSA UV fény kivágás, izolálás a fehérje DNS komplex kötés kovalens lesz
 minta marker. : Br. MS SDS-PAGE. jelölt fehérje. DNázI kezelés. EMSA. UV fény. kivágás, izolálás. a fehérje DNS komplex kötés kovalens lesz.")
65
A mRNS mennyiségének meghatározása
Northern-blot és hibridizáció reverz-transzkripció kapcsolt kvantitatív PCR korrekt, de belső összehasonlító standardokat igényel

66
In vitro transzkripció
Regulátor G mentes kazetta régió (3-400 bp) Promóter transzkripcionálisan aktív sejt vagy sejtmagi extraktum + ATP, CTP, 32P-UTP, a képződött radioaktív RNS analizise denaturáló PAGE-n. csak a fenti bp-os fragmentum fog látszani, a többi tartalmaz G-t ezek transzkirpciója leáll mennyiségi becslés: denzitometriásan, vagy PhosphorImager-rel elsősorban transz szabályozó elemek hatásának vizsgálatára alkalmas
 Promóter. transzkripcionálisan aktív sejt. vagy sejtmagi extraktum. + ATP, CTP, 32P-UTP, a képződött radioaktív RNS analizise denaturáló PAGE-n. csak a fenti bp-os fragmentum fog látszani, a többi tartalmaz G-t ezek transzkirpciója leáll. mennyiségi becslés: denzitometriásan, vagy. PhosphorImager-rel. elsősorban transz szabályozó elemek hatásának vizsgálatára alkalmas.")
67
Riporter gének alkalmazása
Regulátor régió riporter gén (pl. lacZ) Promóter SEJT -galaktozidáz (LacZ) -galaktozidáz aktivitás mérés feltételezi, hogy nincs poszt-transzkripciós szabályozás, és az enzim specifikus aktivitása is minden mintára egyforma aktivitás mérés kiküszöbölése GFP: Green Fluorescent Protein, gerjesztve floureszkál a kapott aktivitást illetve szignált normálni kell a sejt vagy protein mennyiségére
 Promóter. SEJT. -galaktozidáz (LacZ) -galaktozidáz aktivitás mérés. feltételezi, hogy nincs poszt-transzkripciós szabályozás, és az enzim specifikus aktivitása is minden mintára egyforma. aktivitás mérés kiküszöbölése. GFP: Green Fluorescent Protein, gerjesztve floureszkál. a kapott aktivitást illetve szignált normálni kell a sejt vagy protein mennyiségére.")
68
Gének koordinált regulációjának két fő mechanizmusa baktériumokban
operon: gének egy lókuszban, közös transzkript, közös regulátor policisztronos elrendezés regulon: gének szétszórva a genomban, közös regulátor operon regulon

69
A TRANSZKRIPCIÓS SZABÁLYOZÁS FŐBB GLOBÁLIS STRATÉGIÁI PROKARIÓTÁKBAN
indukálószer represszor inaktív represszor derepresszált KATABOLIKUS aktív represszált BIOSZINTETIKUS indukált aktív aktivátor inaktív aktivátor RNS polimeráz negatív szabályozás pozitív szabályozás indukció represszió inaktív aktivátor inaktív aktív aktivátor indukálószer

70
A transzkripciós faktorok és aDNS közötti specifikus kölcsönhatás
Hélix-turn-hélix (HTH) Motívumok: csgD: NNEIARSLFISENTVKTH LY merR: IGEVALLCDINPVTLRAWQR luxR: SWDISKILGCSERTVTFHLT lehet a faktor N vagy C terminálisán, a másik végen szokott lenni a ligand, kofaktor kötő régió
 Motívumok: csgD: NNEIARSLFISENTVKTH LY. merR: IGEVALLCDINPVTLRAWQR. luxR: SWDISKILGCSERTVTFHLT. lehet a faktor N vagy C terminálisán, a másik végen szokott lenni a ligand, kofaktor kötő régió.")
71
BAKTERIÁLIS TRANSZKRIPCIÓS FAKTOROK FŐBB CSALÁDJAI
Faktor család Tagok AraC család AraC, MelR, RhaS, RhaR, SoxS LysR család LysR, OxyR, MetR, CysB Crp család Crp, Fnr MerR család SoxR Két komponensű NarL, OmpR, Arc szabályozó család Lac represszor család LacI, GalR MetJ család MetJ

72
Aktiváció a gén expresszióban I.
Kölcsönhatás: - CTD-nel (CRP) - 70 4-es régiójával ( cI aktivátor) - NTD-nel (CRP) - alegységgel (DnaA) - ’ alegységgel (N4 single-stranded DNA kötő fehérje) - CTD-nel és 70 4-es régiójával (FNR) Positive activation of gene expression Virgil A Rhodius, Stephen JW Busby Current Opinion in Microbiology 1998, 1:
 - 70 4-es régiójával ( cI aktivátor) - NTD-nel (CRP) - alegységgel (DnaA) - ’ alegységgel. (N4 single-stranded DNA kötő fehérje) - CTD-nel és 70 4-es régiójával (FNR) Positive activation of gene expression. Virgil A Rhodius, Stephen JW Busby. Current Opinion in Microbiology 1998, 1:")
73
Aktiváció a gén expresszióban II.
Promóter konformáció megváltoztatása: - “-35” és “-10” régió azonos oldalra kerül (MerR, SoxR) - DNS visszahajlik és az aktiváló cis szekvencia RNAP fölé kerül - DNS konformáció változást indukál (FIS, IHF) Positive activation of gene expression Virgil A Rhodius, Stephen JW Busby Current Opinion in Microbiology 1998, 1:
 - DNS visszahajlik és az aktiváló cis szekvencia RNAP fölé kerül. - DNS konformáció változást indukál (FIS, IHF) Positive activation of gene expression. Virgil A Rhodius, Stephen JW Busby. Current Opinion in Microbiology 1998, 1:")
74
Távoli aktivátor helyek segítséget igényelnek
DNS-hajlító fehérje (pl. IHF) Specifius kötőhely
 Specifius kötőhely.")
75
Transzkripció repressziója baktériumokban

76
FNR O2 - fumarát nitrát reduktáz regulátor
- citoplazmatikus szenzor-regulátor - dimer[4Fe-4S]2+ DNS-t köt - monomer[2Fe-2S]2+ inaktív - aenaerob respirációra (+) vagy (-) hatás - pO2 1 mbar alatt - TTGAT-N4-ATCAA konszenzus szekvencia - [2Fe-2S] [4Fe-4S]2+ (in vitro) Cys, Fe, DTT, NifS - Pseudomonas: ANR; Bacillus: FNR Rhodobacter sphaeroides: FnrL O2 FNRred FNRox
 vagy (-) hatás. - pO2 1 mbar alatt. - TTGAT-N4-ATCAA konszenzus szekvencia. - [2Fe-2S]2+ [4Fe-4S]2+ (in vitro) Cys, Fe, DTT, NifS. - Pseudomonas: ANR; Bacillus: FNR. Rhodobacter sphaeroides: FnrL. O2. FNRred. FNRox.")
77
Két komponensű szabályozó rendszerek
Komponensek - egy szenzor kináz és egy DNS kötő regulátor - E. coli genom 2% - kb 50 különböző 2 komponensű rendszer - 3 alcsalád: OmpR, FixJ és NtrC

78
A bakteriális kétkomponensű szabályozó rendszerek működése elve
szenzor kináz fehérje DNS kötő fehérje Érzékelő Foszforilációs Felvevő DNS kötő szignál transzfoszforiláció DNS

79
ArcA/B O2 - aerobic respiratory control
- ArcB (szenzor kináz): sejt redox és metabolikus helyzet (elektron transzport változást érzékel) - ArcA(citoplazmatikus regulátor): ArcB foszforilálja aktív - pO mbar között - TATTTaa konszenzus szekvencia - Haemophilus: ArcA - E. coli homológ gén: OmpR ArcA ArcB P O2 P ArcAP
: sejt redox és metabolikus helyzet. (elektron transzport változást érzékel) - ArcA(citoplazmatikus regulátor): ArcB foszforilálja aktív. - pO2 1-5 mbar között. - TATTTaa konszenzus szekvencia. - Haemophilus: ArcA. - E. coli homológ gén: OmpR. ArcA. ArcB. P. O2. P. ArcAP.")
80
Kétkomponensű rendszerek vége
NarX/L és NarP/Q - nitrát regulator - NarX és NarQ: membrán szenzor kináz - NarL és NarP: citoplazmatikus regulátor - szignál: nitrát és nitrit - nitrát metabolizmusra hat - NarL és NarP különböző génekre különböző atás a génexpresszió finom hangolása NO3 NarPP NarX NarQ NarLP P Kétkomponensű rendszerek vége

81
A lac operon kettős szabályozása
laktóz (allolaktóz) indukál glükóz gátol, cAMP/CAP-n keresztül glükóz/egyéb cukor kiiktatása tápból nem célszerű glükóz szabályozás kikapcsolása
 indukál. glükóz gátol, cAMP/CAP-n. keresztül. glükóz/egyéb cukor kiiktatása tápból nem célszerű. glükóz szabályozás kikapcsolása.")
82
lac (és trp) alapú promóterek
erősség “-35” “-10” trp AATGAGCTGTTGACAATTAATCATCGAACTAGTTAACTAGTACGCA tac AATGAGCTGTTGACAATTAATCATCCGGCTCGTATAATGTGTGGA lacUV5 CCCCAGGCTTTACACTTTATGCTTCCGGCTCGTATAATGTGTGGA lac CCCCAGGCTTTACACTTTATGCTTCCGGCTCGTATGGTGTGTGGA lacUV5 lac promóter UV

83
A transzkripciós faktorok sokoldalúak….
Ara C Ara PBAD represszió +Ara PBAD indukció RNS polimeráz PBAD AraC

84
Transzkripció termináció baktériumokban
Rho független Rho függő a) hurok képződés indul b) hurok képződés indul hibrid destabilizálódik c) termináció a) Rho kötődik és üldözi a polimerázt b) hurok képződés, polimeráz megáll c) Rho helikáz felszabadítja a transzkriptet, termináció
 hurok képződés indul. b) hurok képződés indul hibrid destabilizálódik. c) termináció. a) Rho kötődik és üldözi a polimerázt. b) hurok képződés, polimeráz megáll. c) Rho helikáz felszabadítja a transzkriptet, termináció.")
85
A transzkripció és a transzláció párhuzamosan megy baktériumokban

86
A triptofán operon szerkezete
protein antranilát szintáz indol-glicerin szintáz triptofán szintáz A triptofán operon szerkezete

87
A triptofán operon szabályozása

88
Attenuáció-termináció – trp operon

89
E. coli-ban fehérje túltermeltetésre használt promóterek

90
mRNS degradáció baktériumokban
mRNS stabilitás prokariótákban néhány perc, eukariótákban órás nagyságrend előbb utóbb minden RNS lebomlik mRNS stabilitását meghatározó faktorok: - belső, saját szerkezet - a környezet hatására bekövetkezett változás a degradációs apparátusban puf operon (a fotoszintetikus komplex komponensei) Rhodobacter capsulatus degradációja O2 hatására felgyorsul policisztronos rendszerek esetén az alegységek arányának szabályozása a mRNS régióinak eltérő stabilitásával
 Rhodobacter capsulatus degradációja O2 hatására felgyorsul. policisztronos rendszerek esetén az alegységek arányának szabályozása. a mRNS régióinak eltérő stabilitásával.")
91
A R. capsulatus puf mRNS régióinak stabilitása

92
mRNS degradáció baktériumokban,
vizsgálati módszerek - transzkripció gátlása (pl. rifampicin) t=0 időpontban, majd időközönként mintavétel és RNS analízis (Northern..) a degradációs mechanizmusban szerepet játszó gének deléciója, hőmérséklet érzékeny expressziós változatának kialakítása - in vitro transzkripció jelölt nukleotidokkal, a kapott termék inkubációja a sejtextraktummal különböző ideig, majd analízis gélelektroforézissel, kvantitálás
 t=0 időpontban, majd időközönként mintavétel és RNS analízis (Northern..) a degradációs mechanizmusban szerepet játszó gének deléciója, hőmérséklet érzékeny expressziós változatának kialakítása. - in vitro transzkripció jelölt nukleotidokkal, a kapott termék inkubációja a sejtextraktummal különböző ideig, majd analízis gélelektroforézissel, kvantitálás.")
93
mRNS-t stabilizáló-destabilizáló tényezők

94
Az 5’ végi struktúra stabilizáló hatása
a stabillizálódás a mRNS hurok struktúrájában van nem csak a riboszóma véd, a stabilizáló effektus átvihető más génekre

95
mRNS-eket stabilizáló (védő) tényezők
5’ végi trifoszfát RNS struktúra riboszóma

96
A degradoszóma felépítése

97
Az RNaseE elsődleges felépítése
Érdekes módon sok bakteriális genomban nincs meg

98
A degradoszóma komponensei I.
Endoribonuclease E (RNáz E) 1061 aminosav 118 kDa fehérje, virtuálisan 180 kDa (oka prolin gazdag régió) felismerő hely: (A/G)AUU(A/T) vagy egy komplex másodlagos struktúra 5' monofoszfátot preferál 5' trifoszfát stabilizál N-terminális régió (50 kDa): endoribonukleáz funkció C-terminális: a degradoszóma egyéb komponenseire megfelelő kötő domének
 1061 aminosav 118 kDa fehérje, virtuálisan 180 kDa (oka prolin gazdag régió) felismerő hely: (A/G)AUU(A/T) vagy egy komplex másodlagos struktúra. 5 monofoszfátot preferál 5 trifoszfát stabilizál. N-terminális régió (50 kDa): endoribonukleáz funkció. C-terminális: a degradoszóma egyéb komponenseire megfelelő kötő domének.")
99
A degradoszóma komponensei II.
PNPase (polynucleotide phosphorylase) 78 kDa alegységek, homotrimer 3' 5' foszfát függő processzív exonukleáz, ribonukleotid difoszfátok képződnek poliadenilációs aktivitás Polyphosphate Kinase (PPK) funkció: ATP regeneráció, polifoszfát (inhibiálja a degradációt) eltávolítás ppk mínusz törzs : megnövekedett mRNS stabilitás 80 kDa alegységek, homotetramer, sok van E. coli-ban
 78 kDa alegységek, homotrimer. 3 5 foszfát függő processzív exonukleáz, ribonukleotid difoszfátok képződnek. poliadenilációs aktivitás. Polyphosphate Kinase (PPK) funkció: ATP regeneráció, polifoszfát (inhibiálja a degradációt) eltávolítás. ppk mínusz törzs : megnövekedett mRNS stabilitás. 80 kDa alegységek, homotetramer, sok van E. coli-ban.")
100
ATP hiányában a hurokstruktúra stabil marad
A degradoszóma komponensei III. Helikáz ATP függő DEAD (Asp-Glu-Ala-Asp) helikáz 50 kDa RhlB a másodlagos struktúrák kinyitása szétroncsolása ATP hiányában a hurokstruktúra stabil marad
 helikáz. 50 kDa RhlB. a másodlagos struktúrák kinyitása szétroncsolása. ATP hiányában a hurokstruktúra stabil marad.")
101
Egyéb – mRNS degradációjában résztvevő – enzimek
RNáz II 70 kDa monomer, a sejt 3' 5' exoribonukláz aktivitásának 90%-a ribonukleotid monofoszfátok képződnek a PNPáz-zal együttes deléciója letális !!! PolyA polimerázok PAPI kDa PAPII kDa poliadeniláció, bázis hosszú mRNS instabilitás

102
A mRNS degradáció mechanizmusa

103
Transzláció prokariótákban

104
A transzláció hatékonyságát meghatározó elemek
30S* + mRNA PK k1 k-1 k2 k-2 IK TK k3 ( 30S + IF1, IF2, IF3, fMet-tRNSMet) ( preiniciációs komplex) ( iniciációs komplex) ( transzlációs komplex) K1 nagy, ha az SD és a rRNS közti kölcsönhatás erős. k2 a SD és a transzlációs start kodon közti távolság, és a régió másodlagos szerkezetének függvénye.
 ( preiniciációs komplex) ( iniciációs komplex) ( transzlációs komplex) K1 nagy, ha az SD és a rRNS közti kölcsönhatás erős. k2 a SD és a transzlációs start kodon közti távolság, és a régió másodlagos szerkezetének függvénye.")
105
Shine - Dalgarno szekvencia
GGAGG a tökéletes komplementer a 16S rRNS 3'végéhez, pl. GAAG GGAG mutáció (maltóz operon) 10 x növekedés a termék mennyiségében a SD szekvencia hosszának növelése nem feltétlenül növeli a transzlációt Hasonlóan a promóterekhez, túl erős kötődés a SD szekvencia és a 16S rRNS 3’ vége között a transzlációt gátolhatja. Transzlációs start kodon kodon gyakoriság AUG 91 % GUG 8 % UUG 1 %
 10 x növekedés a termék mennyiségében. a SD szekvencia hosszának növelése nem feltétlenül növeli a transzlációt. Hasonlóan a promóterekhez, túl erős kötődés a SD szekvencia. és a 16S rRNS 3’ vége között a transzlációt gátolhatja. Transzlációs start kodon. kodon. gyakoriság. AUG. 91 % GUG. 8 % UUG. 1 %")
106
Az AUG előtti szekvenciák
20x-os különbség UAU, CUU >> UUC, UCA, AGG ("-1 triplet) G-k kerülendők Az AUG utáni szekvenciák 30x -oskülönbségek lehetnek AAA(Lys) > AAG(Lys) > UUU(Phe), AUC/A(Val), GUA(Ala)... A természetes gének között is az AAA(Lys) és GCU(Ala) transzlációs szabályozás a 2. kodonnal +10 régió AT gazdag
 G-k kerülendők. Az AUG utáni szekvenciák. 30x -oskülönbségek lehetnek. AAA(Lys) > AAG(Lys) > UUU(Phe), AUC/A(Val), GUA(Ala)... A természetes gének között is az AAA(Lys) és GCU(Ala) transzlációs szabályozás a 2. kodonnal. +10 régió AT gazdag.")
107
Távolság a start kodon és a SD között
- meglehetősen érzékeny, általban bázispár megengedett. - Génfüggő, egy gén esetén általában csak kicsit változhat. A köztes szekvencia összetétele “A” jó, ha van (2x-es stimulus), “C” mindegy “U” a "-4"-es pozícióban hatékony “G” gátol (3x-os gátlás) Upstream nemtranszlálódó szekvencia Nem nagyon vizsgált, de kell, mert eltávolítása megszünteti a transzlációt (ld lac). Minimum 10 bp kell. Esetleg valamilyen faktorok kötődnek hozzá.
, C mindegy. U a -4 -es pozícióban hatékony G gátol (3x-os gátlás) Upstream nemtranszlálódó szekvencia. Nem nagyon vizsgált, de kell, mert eltávolítása megszünteti a transzlációt (ld lac). Minimum 10 bp kell. Esetleg valamilyen faktorok kötődnek hozzá.")
108
Kodon felhasználási preferencia
Bár a kodonok gyakorlatilag univerzálisak, a különböző sejtféleségek különböző gyakorisággal használják az azonos aminosavakat kódoló tripleteket. Ha a termeltetendő fehérje génjének kodon használata nagyon eltér a gazdasejtétől, akkor meg kell szintetizáltatni a gént a gazda sejt preferenciájának megfelelően. Alternatív megoldás: a gazdasejt kodon preferenciájának megváltoztatása, ritka kodonokhoz tartozó tRNS gének bevitelével

109
Kodon felhasználási preferencia – táblázat
Sphingomonas paucimobilis AmAcid Codon Number / Fraction Gly GGG Gly GGA Gly GGT Gly GGC Glu GAG Glu GAA Asp GAT Asp GAC Val GTG Val GTA Val GTT Val GTC Ala GCG Ala GCA Ala GCT Ala GCC Arg AGG Arg AGA Ser AGT Ser AGC Lys AAG Lys AAA Asn AAT Asn AAC Met ATG Ile ATA Ile ATT Ile ATC Thr ACG Thr ACA Thr ACT Thr A CC Trp TGG End TGA Cys TGT Cys TGC End TAG 0.10 End TAA Tyr TAT Tyr TAC Leu TTG Leu TTA Phe TTT Phe TTC Ser TCG [gbbct]: 50 CDS's (13879 codons) egy adott gén esetén a kodon preferencia alapján a gén E. coli-ban, vagy élesztőben való expresszálhatósága in silico becsülhető.
 egy adott gén esetén a kodon preferencia alapján a gén E. coli-ban, vagy élesztőben való expresszálhatósága in silico becsülhető.")
110
A transzláció terminációja
STOP kodon UAA UGA UAG kötődő faktor RF1, RF2 RF1 RF2 UAA >> UGA, UAG Az esetek 80 %-ában: UAAU

111
PROTEOLÍZIS Abnormális fehérjék proteolitikus lebontási mechanizmusa
E. coli-ban Abnormális fehérjék, sejten belüli aggregátumok ATP függő endoproteázok polipeptidek < 1500 Da endoproteázok peptidek aminosavak oldékony mono-, di- és tripeptidázok

112
La (Lon) a fő ATP függő proteáz
87 kDa egységekből felépülő homotetramer, ATP és Mg2+ függő, az ATP-áz aktivitás nem a polipeptid kötés felbontásához, hanem a degradálandó fehérjén való végiglovagláshoz kell, in vitro 2 ATP/ peptidkötés, in vivo ez változhat, és ennél nagyobb. hasonló fehérjék előfordulnak magasabbrendűekben is, pl. egy máj mitondriális proteáz immunológiailag keresztreaktív abnormális fehérjék, és rövid élettartamú (pl. szabályozó fehérjék bontása) szerin proteáz, hidrofób jellegű szekvenciáknál hasít alapállapotban inaktív, a szubsztrát fehérje hozzákapcsolódása aktiválja, a lon gén terméke, és a sokk fehérjékhez kapcsolódó szabályozás alatt áll: 32 faktor, ami a htpR (rpoH) gén terméke
 szerin proteáz, hidrofób jellegű szekvenciáknál hasít. alapállapotban inaktív, a szubsztrát fehérje hozzákapcsolódása aktiválja, a lon gén terméke, és a sokk fehérjékhez kapcsolódó szabályozás alatt áll: 32 faktor, ami a htpR (rpoH) gén terméke.")
113
A proteolitikus ciklus
inaktív proteáz (4 ADP) protein szubsztrát 4 ADP 4 PO43- 4 Mg 2+ 4 ATP - Mg2+ alloszterikus aktiválás aktív proteáz (4 ATP)
 protein. szubsztrát. 4 ADP. 4 PO43- 4 Mg 2+ 4 ATP - Mg2+ alloszterikus aktiválás. aktív proteáz. (4 ATP)")
114
Genetikai vonatkozások
léteznek inszerciós, deléciós lon mutánsok A lon- mutáció másodlagos hatásai Poliszaharid burok képzés (főleg alacsonyabb hőmérsékleten, mint 34oC) Megoldás: 37oC-on növeszteni galE struktúrgének inaktiválása cps (capsule) struktúrgének inaktiválása rcsB, rcsA szabályozó régió mutánsok A lon mutánsok UV érzékenyek DNS sérülés filamentáció Gazdag médiumon akár letális is lehet ok: SOS indukálható sejtosztódás inhibitor SulA. Megoldás: - ne használjunk yeast extraktot, hanem tryptont - sulA mutánsok
 Megoldás: 37oC-on növeszteni. galE struktúrgének inaktiválása. cps (capsule) struktúrgének inaktiválása. rcsB, rcsA szabályozó régió mutánsok. A lon mutánsok UV érzékenyek. DNS sérülés filamentáció. Gazdag médiumon akár letális is lehet. ok: SOS indukálható sejtosztódás inhibitor SulA. Megoldás: - ne használjunk yeast extraktot, hanem tryptont. - sulA mutánsok.")
115
ATP hidrolízis mellett
Ti (Clp) a másik ATP függő proteáz 81 (ClpA) és 21 kDa (ClpP) alegységekből álló heterodimer, ez multimerizálódik, a natív enzim kb 700 kDa móltömegű ATP és Mg2+ faktorok szükségesek hidrofób részeknél hasít, de a specificitás eltér a La-tól az alegységek külön multimerizálódnak HtpR szabályozás alatt áll Tulajdonság Ti (Clp) proteáz ClpP ClpA aktivitás fehérje degradáció ATP hidrolízis mellett peptidáz ATP független ATP-áz fehérje aktivált Kofaktor Mg2+ - Inhibítor MalNEt, iPr2P-F iPr2P-F MalNEt Stabilizálás ATP, glicerin hőstabil natív méret 700 kDa 260 kDa 160 kDa alegységek 6 ClpA + 12 ClpP 21 kDa 81 kDa
 a másik ATP függő proteáz. 81 (ClpA) és 21 kDa (ClpP) alegységekből álló heterodimer, ez multimerizálódik, a natív enzim kb 700 kDa móltömegű. ATP és Mg2+ faktorok szükségesek. hidrofób részeknél hasít, de a specificitás eltér a La-tól. az alegységek külön multimerizálódnak. HtpR szabályozás alatt áll. Tulajdonság. Ti (Clp) proteáz. ClpP. ClpA. aktivitás. fehérje degradáció. ATP hidrolízis mellett. peptidáz. ATP független. ATP-áz. fehérje aktivált. Kofaktor. Mg2+ - Inhibítor. MalNEt, iPr2P-F. iPr2P-F. MalNEt. Stabilizálás. ATP, glicerin. hőstabil. natív méret. 700 kDa. 260 kDa. 160 kDa. alegységek. 6 ClpA + 12 ClpP. 21 kDa. 81 kDa.")
116
Genetikai vonatkozások
Rendelkezésre állnak clp mutánsok a mutáns egyedül nem, de a lon- nal együtt hatékony Egyéb proteázok OmpT külső membránhoz kapcsolódó proteáz DegP periplazmatikus proteáz T4 fág fertőzés stabilizálja a fehérjéket pin gén terméke: La inhibitor

117
Példák proteázaikban mutáns E. coli törzsekre
genotípus Kiindulási törzs Megjegyzés lon mutánsok SG1117 Dgal Dlac lon-146::DTn10 leu sup+ rec+ HB101 Tetraciklin rezisztens, jól transzformálható SG12041 Dgal, Dlon-510 sulA recA C600 nincs filamentáció Rec- lon htpR mutánsok SG21163 Dlon-510 supFts htpRam165 MC4100 hőmérséklet érzékeny lon clp mutánsok SG12044 Dgal Dlon-510 sulA clpA319::Dkan kanamycin rezisztens lon clp htpR mutánsok SG21173 DclpA319::Dkan Dlac kanamycin rezisztens,

118
FÚZIÓS FEHÉRJÉK A fúziót gén szinten hozzuk létre úgy, hogy a fúziós partnerek génjeit a megfelelő leolvasási keretben úgy klónozzuk össze, hogy az 5’ vég felőli génhez tartozó transzlációs stop kodon ne szerepeljen. Így egy polipeptidláncot kapunk, amelyben a kiinduási fehérjék egymást követő régiókként helyezkednek el. ELŐNYEI 1. Kis peptidek esetén a fúzió proteolitikus stabilitást jelenthet 2. Óriási előny a tisztítás során, affinitás oszlopok 3. szignálpeptiddel való fúzió, szekretált fehérje (limitált) 4. Az in vitro hasítás sokszor pontosabb, intaktabb N-terminális szekvenciát eredményez
 4. Az in vitro hasítás sokszor pontosabb, intaktabb N-terminális szekvenciát eredményez.")
119
Fúziós fehérjék oldhatósága
Oldhatatlan "iclusion bodies" trpEII, vagy cII, nincs proteolízis, differenciális centrifugálással az "inclusion body" könnyen tisztítható, probléma: nem aktív fehérje pl sometostatin, inzulin A, B, ellenanyag termelés Oldható forma biológiailag aktív fehérje, affinitás oszlopon való tisztítás problémák a stabilitással, kevésbé jósolható

120
Egyéb hasznosítási lehetőségek
1. a szignál peptidekkel való fúzió előnyei (ld. később) a periplazmában, illetve az extracelluláris térben oxidatívabb környezet, diszulfid hidak kialakulása, pl. EGF, IGF A periplazmában az összfehérje 4%-a található, értelemszerűen kevesebb proteáz,könnyebb tisztítás 2. A fúzionált fehérje linkerként szolgál, és így a kívánt fehérje immobilizálható 3. funkcionális, struktúrális vizsgálatok, pl. immunológiai felhasználás alkalmasan választott fúziós partner esetén nincs szükség hasításra.
 a periplazmában, illetve az extracelluláris térben oxidatívabb környezet, diszulfid hidak kialakulása, pl. EGF, IGF. A periplazmában az összfehérje 4%-a található, értelemszerűen kevesebb proteáz,könnyebb tisztítás. 2. A fúzionált fehérje linkerként szolgál, és így a kívánt fehérje immobilizálható. 3. funkcionális, struktúrális vizsgálatok, pl. immunológiai felhasználás. alkalmasan választott fúziós partner esetén nincs szükség hasításra.")
121
A fúziós fehérjék tisztításának elve
I. 1. affinitás kromatográfia II. hasítás III. 2. affinitás kromatográfia tiszta fehérje fúziós partner célfehérje

122
A fúziós fehérjék típusai I.
I. Szekréció Megjegyzés pl. humán növekedési hormon alkalikus foszfatáz szignál szekvenciával S X T P II. Polimerizáció fehérje-élettartam növekedés, pl. proinzulin 1-2 percről 60 perc, IGF 200x-os növekedés P T X X III. C-terminális fúzió a fúziós partner intakt szabályozó régiója használható, pontosabb proteolitikus hasítás A X T P IV. N-terminális fúzió amino terminális szekvenciák meghatározása, a proteolititkus hasítás csonkot hagyhat a “B” N-terminálisán X B T P

123
A fúziós fehérjék típusai II.
V. A szekréció és a C-terminális fúzió kombinációja A X S T P a termék folyamatosan kinyerhető affinitáskromatográfiával VI. Kettős fúzió A X B T P nagy tisztaságú, nagy stabilitású termék, hátrány: két tisztítás, hasítás szükséges P T A B X P T X A B VII. Szekréció-inszerció X I S T P membrán fehérjék termeltetése a membránban

124
Fúziós partnerfehérjék
Fehérje Származási hely molekulasúly (kDa) szekréció ligand b-galaktozidáz Escherichia coli 116 - TPEG, APTG Protein A Staphylococcus aureus 31 + IgG Z szintetikus 7 CAT 24 klóramfenikol sztreptavidin Streptomyces 13 biotin PhoS 36 hidroxilapatit Protein G Streptococci 28 albumin MBP 40 keményítô GST 26 glutation poliarginin 1-3 ioncsere poliglutamát 1-2 polihisztidin 1-7 Ni2+, Zn2+, Cu2+ CAT: klóramfenikol-acetiltranszferáz; Z: A Protein A fehérje IgG kötô doménje; PhoS: foszfát kötô fehérje; MBP: maltóz-kötô fehérje; GST: glutation S-transzferáz; TPEG: p-aminofenil--d-tiogalaktozidáz; APTG: p-aminofenil--d-tiogalaktozid.
 szekréció. ligand. b-galaktozidáz. Escherichia coli TPEG, APTG. Protein A. Staphylococcus aureus IgG. Z. szintetikus. 7. CAT. 24. klóramfenikol. sztreptavidin. Streptomyces. 13. biotin. PhoS. 36. hidroxilapatit. Protein G. Streptococci. 28. albumin. MBP. 40. keményítô. GST. 26. glutation. poliarginin ioncsere. poliglutamát polihisztidin Ni2+, Zn2+, Cu2+ CAT: klóramfenikol-acetiltranszferáz; Z: A Protein A fehérje IgG kötô doménje; PhoS: foszfát kötô fehérje; MBP: maltóz-kötô fehérje; GST: glutation S-transzferáz; TPEG: p-aminofenil--d-tiogalaktozidáz; APTG: p-aminofenil--d-tiogalaktozid.")
125
FÚZIÓS FEHÉRJÉK HASÍTÁSI MÓDOZATAI I.
KÉMIAI hasító ágens hasítási hely brómcián - Met hangyasav - Asp Pro - hidroxilamin - Asn Gly - ENZIMATIKUS aminopeptidázok 1 - 3 aminosavat hasítanak az N-terminálisról karboxipeptidázok 1 - 2 aminosavat hasítanak a C-terminálisról

126
FÚZIÓS FEHÉRJÉK HASÍTÁSI MÓDOZATAI II.
ENZIMATIKUS Endopeptidázok ENZIM HASÍTÁSI HELY kollagenáz - Pro - Val Gly - Pro - enterokináz - Asp - Asp - Asp - Lys Xa faktor - Ile - Glu - Gly - Arg trombin - Gly - Pro - Arg tripszin - Arg, Lys klosztripain - Arg Ala64-szubtilizin - Gly - Ala - His - Arg

127
A fehérje szekréció Nómenklatúra: szekréció: az extracelluláris térbe
export: az sejtmembránokba vagy a periplazmatikus térbe Előnyei A tisztítás sokkal egyszerűbb, esetleg folyamatos üzemű lehet Az periplazma illetve az extracelluláris tér sokkal oxidatívabb, korrektebb protein folding A fehérje degradáció sokkal csekélyebb mértékű, proteolitikusan stabilabb termékek

128
Baktériumok membránszerkezete

129
SZEKRÉCIÓ E. coli-ban nem hatékony vagy nem teljes transzport a membránon keresztül alacsony transzport-kapacitás, stresszválasz, idő előtti fehérjedegradáció ha a transzporthoz a fehérje poszttranszlációs módosítása szükséges, akkor ennek hiánya kis molekulákra, mint növekedési faktorok vannak jó eredmények nagy molekulák esetén problémák lehetnek, pl az ER hiánya (módosítások színhelye) vannak fehérjék, amelyek nem szekretálhatóak, esetleg letális, pl -galaktozidáz fúziós fehérjék exportja
 vannak fehérjék, amelyek nem szekretálhatóak, esetleg letális, pl -galaktozidáz fúziós fehérjék exportja.")
130
Fő protein transzport mechanizmusok baktériumokban
Kotranszlációs transzport SRP, signal recognition particle, és receptora E. coli homológ: Ffh forty five homolog, FtsY Poszttranszlációs transzport Sec útvonal: egyszerű kofaktor mentes polipeptidekre Tat útvonal: több alegységes enzimekre, amelyek kofaktorokat is tartalmaz(hat)nak transzport: a fehérje összeszerelődése után
nak. transzport: a fehérje összeszerelődése után.")
131
Sec útvonal E. coli-ban - szignál szekvencia,
- szignál szekvencia, - szignál peptidázok, szignál peptidázI (lep), Ala-X-Ala felismerőhely - szignál peptidázII (lsp), szignál szekvenciák eltávolítása lipoproteinekrôl - sec A, B, D, E, Y gének mutációja a prekurzorok felhalmozódásához vezet, Némi átfedés van közöttük. - Régebben prl nómenklatúra is volt SecA, 102 kDa, citoplazmatikus, és belső membránkötött, ATP-áz aktivitás, kölcsönhatás a szállítandó fehérjével SecB szekréció kompetens konformációért felelős SecYEFG a csatorna kialakításáért felelős SecD a periplazmatikus térbe való “leválásért”
, Ala-X-Ala felismerőhely. - szignál peptidázII (lsp), szignál szekvenciák eltávolítása lipoproteinekrôl. - sec A, B, D, E, Y gének mutációja a prekurzorok felhalmozódásához vezet, Némi átfedés van közöttük. - Régebben prl nómenklatúra is volt. SecA, 102 kDa, citoplazmatikus, és belső membránkötött, ATP-áz aktivitás, kölcsönhatás a szállítandó fehérjével. SecB szekréció kompetens konformációért felelős. SecYEFG a csatorna kialakításáért felelős. SecD a periplazmatikus térbe való leválásért")
132
SZIGNÁL SZEKVENCIÁK GRAM- BAKTÉRIUMOKBAN I.
Standard szignál peptid COOH “N” “H” “C” 1 K vagy R hidrofób hélix, sok A és L, nem lehet P, K, R, D, E, H, R fordulat P G hasítóhely A S -1 +1 X semleges vagy negatív fM V -3 Lipoprotein szignál peptid “C” COOH “N” “H” 1 K vagy R hidrofób hélix, sok A és L, nem lehet P, K, R, D, E, H, R hasítóhely A G -1 +1 D lipoprotein kiválasztó szignál semleges vagy negatív fM L

133
SZIGNÁL SZEKVENCIÁK II.
Gram-pozítív baktériumok a szignál peptid szignifikánsan hosszabb kiterjedtebb “H” régió Eukarióták a “H” régióban a Leu dominál nem olyan szigorú a (-1, -3) szabály a hasítóhely után nincs kitüntetett aminosav az eukarióta szekretált fehérjéket az E. coli export mechanizmusa felismer(het)i, illetve fordítva
 szabály. a hasítóhely után nincs kitüntetett aminosav. az eukarióta szekretált fehérjéket az E. coli export mechanizmusa felismer(het)i, illetve fordítva.")
134
A Sec függő fehérje transzlokáció modellje E. coli-ban
ATP ADP+Pi SPáz Y E G A B D H+ periplazma citoplazma INICIÁCIÓ TRANSZLOKÁCIÓ KISZABADULÁS N+ C+ C N Átjutás a C-terminális véggel, ez az áltatános Átjutás az N-terminális véggel, inszerció

135
A Sec útvonal - másképpen

136
A Sec útvonal - másképpen

137
A prokarióta és eukarióta transzportmechanizmusok összehasonlítása

138
Az SRP (Ffh-FtsY) útvonal
membrán fehérjékre jellemző A Sec és az SRP közötti választás: “trigger” faktor

139
+ + Signal sequences for targeting Sec szignál peptid
erősen hidrofób + n-region h-region c-region SRP: erősen hidrofób szignál, nem hasad le Twin-arginine szignál peptid moderáltan hidrofób + N S/T-R-R-x-F-L-K n-régió h-régió c-régió

140
A periplazmatikus hidrogenázok bioszintézise citoplazmatikus összeszerelődési modell
Holoenzim Periplazma csatorna Citoplazma szignál peptid prekurzor kofaktor(ok)
")
141
The twin-arginine translocase (Tat) proteins
A TAT modell I. The twin-arginine translocase (Tat) proteins citoplazma membrán periplazma N C TatA TatE TatB TatC
 proteins. citoplazma. membrán. periplazma. N. C. TatA. TatE. TatB. TatC.")
142
aktív formát felvett enzim
A TAT modell II. citoplazma TatC TatB TatA membrán aktív formát felvett enzim

143
Rekombináns fehérjék termeltetési stratégiái
Két fő stratégia: Aktív formában termeltetni a fehérjét, ahogy in vivo előfordul, szekretáltatni, hogy a diszulfid hidak kialakuljanak ha a fehérje mérgező a gazdasejtre, akkor először inaktív fehérjét állítunk elő, majd ezt aktiváljuk, ehhez kell az in vitro “refolding” Az aktív forma esetén “csak” tisztítani kell a fehérjét

144
REKOMBINÁNS FEHÉRJÉK AKTÍV FORMÁBA VITELE
Három fő lépés: 1. Az oldhatatlan “inclusion body”-t szolubilizáljuk, alacsony vagy magas pH, emelt hőmérséklet, detergensek, magas koncentrációjú szervetlen sók, vagy szerves oldószerek. A legelterjedtebbek: urea, vagy guanidin-hidroklorid, esetleg redukálószer (pl. DTT) jelenlétében. 2. Refolding híg fehérje oldattal - ha nincs diszulfid híd, akkor a fehérje denaturálószer lassú eltávolítása - ha diszulfid hidak vannak, akkor a renaturáció mellett a diszulfid hidakat is ki kell alakítani, szerkezet stabilizáló hatásuk van. levegőztetés, tiol-diszulfid párok hozzáadása, pH optimalizálása 3. Az aktív és inaktív forma szétválasztása pl. affinittás kromatográfia, preparatív natív PAGE, RP-HPLC, FPLC stb.
 jelenlétében. 2. Refolding. híg fehérje oldattal. - ha nincs diszulfid híd, akkor a fehérje denaturálószer lassú eltávolítása. - ha diszulfid hidak vannak, akkor a renaturáció mellett. a diszulfid hidakat is ki kell alakítani, szerkezet stabilizáló hatásuk van. levegőztetés, tiol-diszulfid párok hozzáadása, pH optimalizálása. 3. Az aktív és inaktív forma szétválasztása. pl. affinittás kromatográfia, preparatív natív PAGE, RP-HPLC, FPLC stb.")
145
T7 RNS polimeráz alapú vektorok
- T7 promoter nagyon szelektív, a gazdában nincs ilyen promoter általában - T7 RNS polimeráz 5x gyorsabb, mint az E. coli-é, azaz már a transzkript is domináns Gazdasejtek: Standard klónozó vektorok klónozásra BL21 (F-, ompT-, lon-) expresszióra, lizogén sejtváltozatok: DE3 bakteriofág, immunitási régió, lacI gén, lacUV5 promoter lacZ eleje + T7 RNS polimeráz génje, integrálva kromoszómába Nagyon toxikus fehérjék esetén pLysS, pLysE T7 lizozim: természetes inhibitora a T7 RNS polimeráznak, E. coli tudja tolerálni, mert nem képes áthatolni a belső membránon pACYC184 vektorba van beépítve, catr, kompatibilis vektor pLysE nagy mennyiséget, pLysS kevesebb mennyiséget termel (ellentétes orientáció, tet promoter CE6 bakteriofág tartalmazza a T7 RNS polimeráz génjét, így az indukció kívülről bejuttatott gén
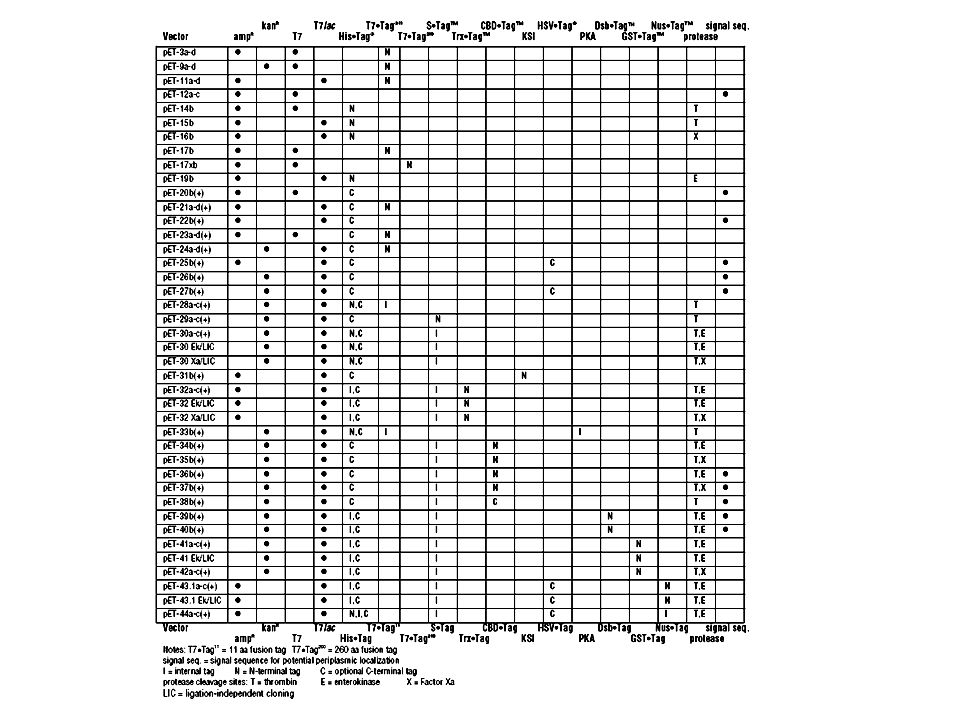
 expresszióra, lizogén sejtváltozatok: DE3 bakteriofág, immunitási régió, lacI gén, lacUV5 promoter lacZ eleje + T7 RNS polimeráz génje, integrálva kromoszómába. Nagyon toxikus fehérjék esetén. pLysS, pLysE. T7 lizozim: természetes inhibitora a T7 RNS polimeráznak, E. coli tudja tolerálni, mert nem képes áthatolni a belső membránon. pACYC184 vektorba van beépítve, catr, kompatibilis vektor. pLysE nagy mennyiséget, pLysS kevesebb mennyiséget termel (ellentétes orientáció, tet promoter. CE6 bakteriofág tartalmazza a T7 RNS polimeráz génjét, így az indukció kívülről bejuttatott gén.")
146
A pET vektorokon alapuló, indukálható fehérje termelés elvi sémája
kromoszóma lacI lacUV5 lacZ T7 RNS polimeráz int vektor Prom. RBS 10 vagy T7/lac termeltetendõ fehérje génje ORI bla

147
A pET rendszer

148
pET vektorok 1. Transzkripciós vektorok T7lac promoteres változatok
pBR322 származékok, ampicillin, vagy kanamicin rezisztensek 10 promoter, a T7 fág 10-es génjének (kapszid) erős promótere 1. Transzkripciós vektorok T7 promoter van, de nincs transzlációs iniciációs szignál különféle verziók, stop szignálok bevitele vagy sem, fúziós partner esetleg a C terminálison Visszaszorulóban vannak T7lac promoteres változatok Nagyon toxikus fehérjék esetén dupla biztosítás: a T7 promotertől startpont környékére a lac operátor szekvencia klónozása, elegendő lacI biztosítása, a vektorba beépítették ellentétes irányban
 erős promótere. 1. Transzkripciós vektorok. T7 promoter van, de nincs transzlációs iniciációs szignál. különféle verziók, stop szignálok bevitele vagy sem, fúziós partner esetleg a C terminálison. Visszaszorulóban vannak. T7lac promoteres változatok. Nagyon toxikus fehérjék esetén dupla biztosítás: a T7 promotertől startpont környékére a lac operátor szekvencia klónozása, elegendő lacI biztosítása, a vektorba beépítették ellentétes irányban.")
149
Két transzkripciós vektor példa

150
Transzlációs vektorok
10 transzkripciós és transzlációs szignálok, mindegyik típusú egy sorozat különböző leolvasási keretekkel Egy rövid fúzió az első 10 aminosavval (g10), de NdeI-gyel elkerülhető. Esetenként 206aa fúzió stabilitás
, de NdeI-gyel elkerülhető. Esetenként 206aa fúzió stabilitás.")
153
Arabinóz alapú expressziós vektorok
Vektor felépítés Szabályozás Szekréciós vektor Kontrollálhatóság

154
Fehérje termeltetés in vitro In vitro transzláció
Előnyei: Oxidatív környezet Citotoxicitás elkerülése, farmakológiai alkalmazások Nincs membrán, a termék azonnal a reaktortérbe kerül gyors Hátrányai: Drága Kitermelés korlátozott Két fő stratégia RNS polimerázzal +sejt extraktummal Tiszta komponensekből

155
Az in vitro fehérjetermeltetés folyamatábrája

156
T7 alapú rendszerek

157
Az igazán in vitro rendszer

Hasonló előadás

 DNS-ből,>")



 prokariótában>")






>")







