Előadást letölteni
1
A baktérium genom integritását védelmező rendszerek
restrikció modifikáció DNS sérülés DNS javítórendszerek

2
Az örökítés lényege, az örökítő anyag állandósága
Általában az idegen eredetű örökítő anyag káros Eukarióta szervezetekben a meiózis, mitózis során a kromoszóma párosodás során csak a megfelelő kromoszómák kerülhetnek egymás mellé Prokarióta szervezetknél ez a mechanizmus nem létezik, mégis van olyan rendszerük, mellyel saját DNS-üket meg tudják jelölni

3
Ilyen módon megkülönböztethető a saját és idegen DNS
A saját DNS jelölésére metiláz rendszer 6-metil adenin (metilezett nukleotidok) 5 metil citozin Metiláz gének hsd, gazdaspecifikus metiláció dam, adenin metilálás Újonnan keletkezett szál metilezése Replikáció kontroll Repair rendszerhez jelölt DNS dcm, citozin metiláz A baktériumban a DNS módosítása biztosít Amelyik DNS nem metilezett azt a gazda restrikciós rendszere elhasítja
 5 metil citozin. Metiláz gének. hsd, gazdaspecifikus metiláció. dam, adenin metilálás. Újonnan keletkezett szál metilezése. Replikáció kontroll. Repair rendszerhez jelölt DNS. dcm, citozin metiláz. A baktériumban a DNS módosítása biztosít. Amelyik DNS nem metilezett azt a gazda restrikciós rendszere elhasítja.")
4
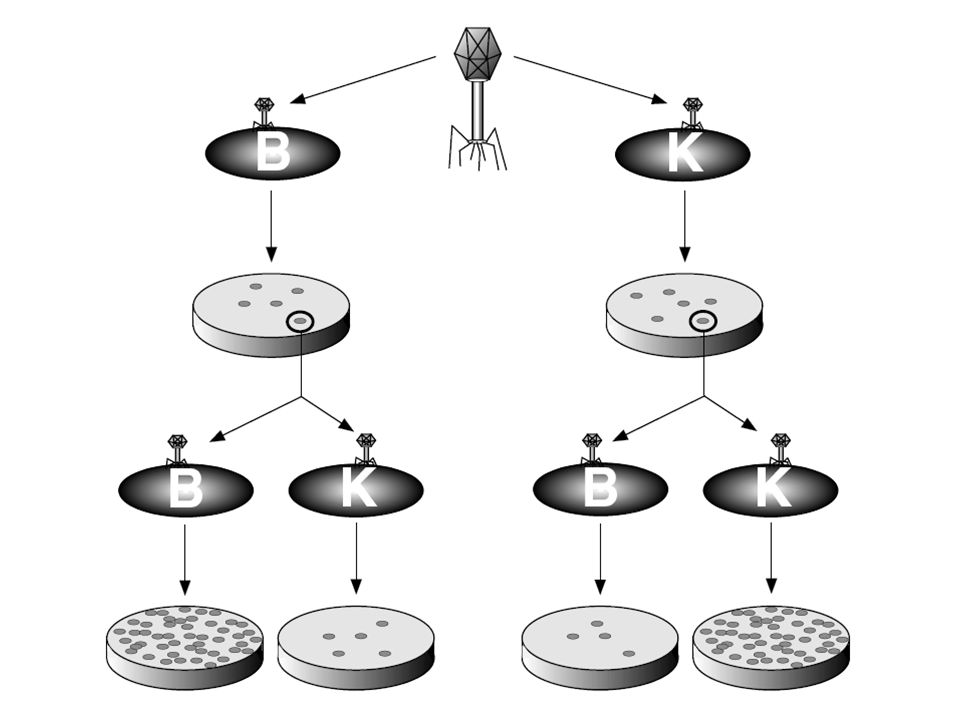
Fág/baktérium rendszerben fedezték fel
A restrikció/modifikáció teremtette meg a „génsebészet, génmérnökség, genetic engineering” alapját Fág/baktérium rendszerben fedezték fel Egy törzsből kikerülő fág képes ugyanazon baktériumtörzs fertőzésére Az a fág, mely más törzsről származik restrikciónak van kitéve A fág DNS a gazda szervezet mintázata szerint módosul (metilálódik) Célja, hogy ellensúlyozza a bakteriális szekvenciák mozgását (plazmidok, fágok)
 Célja, hogy ellensúlyozza a bakteriális szekvenciák mozgását (plazmidok, fágok)")
6
Más védekezés fágok ellen

7
RM rendszer mint addikiós modul: Toxin/Antitoxin

8
Stimulálja a rekombinációt

9
A restrikció/modifikáció lényege tehát
A baktérium törzs DNS-e metilálódik (a saját DNS jelölődik, vagy a baktériumban lévő fág DNS-e, ha fágfertőzés van) A nem metilált DNS-t a restrikciós enzimek hasítják (baktériumsejtben lévő idegen /fág/ DNS elbomlik) A metilezett DNS-t nem hasítja a restrikciós enzim (a sajátként jelölt DNS védett, illetve az ugyanazon a törzsből származó fág DNS is védett)
 A nem metilált DNS-t a restrikciós enzimek hasítják (baktériumsejtben lévő idegen /fág/ DNS elbomlik) A metilezett DNS-t nem hasítja a restrikciós enzim (a sajátként jelölt DNS védett, illetve az ugyanazon a törzsből származó fág DNS is védett)")
10
Restrikciós modifikációs rendszerek
I típus II típus, leggyakoribb III típus IV típus

11
Felismerési szekvencia
I típus II típus III típus (IV típus) Fehérje szerkezet Bifunkcionális enzim 3 alegységgel Endonukleáz és metiláz 2 külön enzim Bifunkcionális enzim 2 alegységgel Felismerési szekvencia Bipartit és asszimmetrikus (TGAN8TGCT) 4-6 bp (gyakran) palindrom szekvencia 5-7 bp asszimmetrikus szekvencia Metilált szekvencia Hasító hely Nem specifikus, a felismerő helytől 1000 bp-ra Ugyanaz, mint a felismerő hely 24-26 bp, a felismerőhelytől visszafele Restrikció/ metiláció Egymást kizáró aktivitás Külön reakció Szimultán reakció Hasítás, ha metilált szekvencia ATP igény van nincs
 Fehérje szerkezet. Bifunkcionális enzim 3 alegységgel. Endonukleáz és metiláz 2 külön enzim. Bifunkcionális enzim 2 alegységgel. Felismerési szekvencia. Bipartit és asszimmetrikus (TGAN8TGCT) 4-6 bp (gyakran) palindrom szekvencia. 5-7 bp asszimmetrikus szekvencia. Metilált szekvencia. Hasító hely. Nem specifikus, a felismerő helytől 1000 bp-ra. Ugyanaz, mint a felismerő hely bp, a felismerőhelytől visszafele. Restrikció/ metiláció. Egymást kizáró aktivitás. Külön reakció. Szimultán reakció. Hasítás, ha metilált szekvencia. ATP igény. van. nincs.")
12
Restrikciós/modifikáló rendszer működése
DNS replikáció DNS állapota metiláz restrikció eredmény Eredeti DNS metilált - Éppen replikálódó DNS hemi metilált + metiláció Replikálódott DNS Idegen DNS nem metilált/ másképpen + ? degradáció

13
II típusú restrikciós/modifikáló rendszer
A teljesen metilált hely nem metilálódik és nem hasítja a restrikciós enzim A hemimetilált helyet nem ismeri fel a restrikciós enzim, de a metiláz teljesen metilálhatja A nem metilált hely szubsztrátja lehet a restrikciós enzimnek (in vitro a modifikáló enzimnek, in vivo hasítás) A legelterjedtebb rendszer (baktériumok 1/3-ban) Gén manipulációban nagyon fontos (sok különböző felismerési helyet ismerünk) 3’ és 5’ túlnyúló végek Kompatibilis és nem kompatibilis Tompa vég
 A legelterjedtebb rendszer (baktériumok 1/3-ban) Gén manipulációban nagyon fontos (sok különböző felismerési helyet ismerünk) 3’ és 5’ túlnyúló végek. Kompatibilis és nem kompatibilis. Tompa vég.")
14
Minden egyes enzim egy funkcióval rendelkezik
A szerkezetük egyszerű (az EcoRI rendszer a legismertebb) A restrikciós enzim két azonos alegység dimerje A metiláz monomer A restrikciós enzimmel közös célszekvencia felismerése (duplikáció vagy konvergens evolúció eredményezhette) Restrikciós hasítás Több baktériumban is találtak ugyanolyan felismerőhelyű enzimet, ez az izoschizomer A nem metilált célszekvenciánál (néha mellette) hasít az enzim Metiláció Egy metilcsoportot ad hozzá majd disszociál A következő metil csoportnál ugyanez ismétlődik
 A restrikciós enzim két azonos alegység dimerje. A metiláz monomer. A restrikciós enzimmel közös célszekvencia felismerése (duplikáció vagy konvergens evolúció eredményezhette) Restrikciós hasítás. Több baktériumban is találtak ugyanolyan felismerőhelyű enzimet, ez az izoschizomer. A nem metilált célszekvenciánál (néha mellette) hasít az enzim. Metiláció. Egy metilcsoportot ad hozzá majd disszociál. A következő metil csoportnál ugyanez ismétlődik.")
15
Kevésbé gyakori, mint a II típusú rendszer
I és III típusú enzimek I típus Multimerekből áll Mindkét reakciót (restrikció/metiláció) elvégzik Kevésbé gyakori, mint a II típusú rendszer 3 különböző méretű alegységből áll (EcoK, EcoB) R restrikció M metiláció S szekvencia felismerés R és M egyszerre nem működhet
 elvégzik. Kevésbé gyakori, mint a II típusú rendszer. 3 különböző méretű alegységből áll (EcoK, EcoB) R restrikció. M metiláció. S szekvencia felismerés. R és M egyszerre nem működhet.")
16
A működéshez, ATP, Mg++, S-adenozil-metionin (SAM) szükséges
Az enzim hozzáköt a felismerőhelyhez Ha az metilált nem történik semmi Ha hemimetilált (frissen replikálódott) metiláz aktivitás stimulálódik Nem metilált szekvenciánál az enzim kötve marad Az enzim egyik vége ATP felhasználásával hurkot alkot Amikor a hurok 1 kb nagyságú az egyik szálat elhasítja, egy másik enzim molekula a másik szálat hasítja
 metiláz aktivitás stimulálódik. Nem metilált szekvenciánál az enzim kötve marad. Az enzim egyik vége ATP felhasználásával hurkot alkot. Amikor a hurok 1 kb nagyságú az egyik szálat elhasítja, egy másik enzim molekula a másik szálat hasítja.")
17
IV típus (nem igazi besorolás)
III típusú rendszer Kevés rendszer ismert (P1 fágé, EcoP1) A metiláz fehérje egyedül vagy komplexben működik A restrikciós fehérje csak komplexben működhet A felismerőhelytől (visszafele) bp-ra hasít IV típus (nem igazi besorolás) A legtöbb rendszerben a restrikciós enzim nem ott hasít, ahol metilált a DNS (azaz a metiláció védettséget ad) A IV típusnál éppen a metilált DNS hasítódik HpaII és MspI izoschizomerek, ugyanazt a szekvenciát ismerik fel HpaII akkor hasít, ha a DNS nem metilált MspI akkor hasít, ha a szekvencia metilált
 A metiláz fehérje egyedül vagy komplexben működik. A restrikciós fehérje csak komplexben működhet. A felismerőhelytől (visszafele) bp-ra hasít. IV típus (nem igazi besorolás) A legtöbb rendszerben a restrikciós enzim nem ott hasít, ahol metilált a DNS (azaz a metiláció védettséget ad) A IV típusnál éppen a metilált DNS hasítódik. HpaII és MspI izoschizomerek, ugyanazt a szekvenciát ismerik fel. HpaII akkor hasít, ha a DNS nem metilált. MspI akkor hasít, ha a szekvencia metilált.")
18
R/M a bakteriofágokban
Gyakran védekeznek a gazda restrikció ellen Saját metilációs enzimmel rendelkeznek Olyan fehérje is létezik, amely gátolj a gazda restrikciós enzimjét Elnevezésük EcoRI Genus, faj rövidítéssel, restrikció, ha több is van akkor római számokkal

19
DNS sérülés Mikroorganizmusok káros környezeti hatásra elvesztik életképességüket Ezt a veszteséget grafikonon ábrázolva dózisgörbét kapunk y=túlélő frakció, x=dózis A túlélési görbe vizsgálatával a sugárzási hatások magyarázhatók voltak, itt mutatkozott először, hogy a környezeti hatásra sérült DNS kijavítódik

20
A legegyszerűbb eset, ha 1 érzékeny hely van
Ezt a fajta viselkedést egy matematikai módszer a célelmélet (target) írja le A túlélési görbe egy populációból,mely N darab azonos szervezetből (sejtből) áll Ez a populáció D dózisú besugárzásnak (vagy más behatásnak) van kitéve Matematikai egyenlettel jellemezhető A legegyszerűbb eset, ha 1 érzékeny hely van Ha ezt sugárzás éri, akkor elpusztul a szervezet dN elpusztultak száma, amit dD dózis ért a fenti arányos a populáció besugárzás előtti egyedszámával N-el -dN/dD=kN, ahol k konstans a dózis hatékonyságának mértéke
 írja le. A túlélési görbe egy populációból,mely N darab azonos szervezetből (sejtből) áll. Ez a populáció D dózisú besugárzásnak (vagy más behatásnak) van kitéve. Matematikai egyenlettel jellemezhető. A legegyszerűbb eset, ha 1 érzékeny hely van. Ha ezt sugárzás éri, akkor elpusztul a szervezet. dN elpusztultak száma, amit dD dózis ért. a fenti arányos a populáció besugárzás előtti egyedszámával N-el. -dN/dD=kN, ahol k konstans a dózis hatékonyságának mértéke.")
21
Integrálás után N=N0 D=0 N=N0e-kD A túlélők aránya S=N/N0 S=N/N0=(e-kD)n itt n=1 lnS a D dózis függvényében egyenest ad, melynek meredeksége –k Ez az egytalálatos (single hit) vagy exponenciális görbe
 vagy exponenciális görbe.")
22
Mi történik, ha olyan populációnk van, melyben az egyes szervezet elpusztításához n helyet kell eltalálni a pusztuláshoz Ebben az esetben az inaktivációhoz legalább n találatra van szükség (legalább, mert statisztikailag néhány hely 2 találatot is kap, de azt feltételezzük, hogy 2 találat ugyanazon a helyen nem hatékonyabb, mint egy találat) Annak valószínűsége, hogy minden n cél találatot kap Pn Pn=(1-e-kD)n (független események) A populációban a túlélők aránya S=1-Pn S=1-Pn=1-(1-e-kD)n S=1-(1-ne-kD+…e-nkD) (kibontva)
 Annak valószínűsége, hogy minden n cél találatot kap Pn. Pn=(1-e-kD)n (független események) A populációban a túlélők aránya S=1-Pn. S=1-Pn=1-(1-e-kD)n. S=1-(1-ne-kD+…e-nkD) (kibontva)")
23
Ha a dózis (D) nagy, akkor a magasabbrendű tagok elhanyagolhatóak lesznek az ne-kD taghoz képest
Azaz nagy dózis esetén S=ne-kD ln S=ln n – kD Kis dózis estén S lassan változik Nagy dózis esetén S=ne-kD érvényesül Az egyenes extrapolációja és y tengelymetszete megadja célpontok (n, target) számát Ha kísérletesen elég nagy dózist tudunk elérni, akkor becsülhetjük a célhelyek számát
 számát. Ha kísérletesen elég nagy dózist tudunk elérni, akkor becsülhetjük a célhelyek számát.")
24
Fágokra egyenes vonalat kapunk Baktériumoknál nem egyértelmű
A populáció nem egységes, egyes sejtek a replikáció más és más fázisában vannak Lehet olyan sejt, ahol több kópia is van az egyik sérül, de a másik ép marad Sok besugárzási károsodás javításra kerül, így a potenciálisan halálos találatok egy részét nem észleljük Ionizáló sugárzás 3 típusú károsodást okozhat Egyszálú törés, legtöbbször a ligáz kijavítja 2 szálú törés, gyakran letális, mert a szabad dsDNS végek a nukleázok támadási helyei Bázisokban történt károsodás (elemi oxigént igényel), gyakran halálos, a sérült bázis akadályozza a replikációt
, gyakran halálos, a sérült bázis akadályozza a replikációt.")
25
Ionizáló sugárzás esetén
A „k” konstans egyfajta mértékegysége a találat valószínűségének Az ionizáció száma egységnyi térfogatra arányos a dózissal V target volume (cél térfogat) A V térfogaton belüli átlagos találatok száma cVD, c arányossági állandó, megmondja hány találatból hány ionizáció keletkezik Véletlenszerű találatnál, n találat valószínűsége V térfogaton belül Poisson eloszlással adható meg P(n)=e-cVD(cVD)nn! Single hit mechanizmusban a túlélők 1 találatot kell kapjanak S=1-P(n) S=e-cVD k=cV, azaz k arányos a cél térfogatával Minél nagyobb a céltábla annál nagyobb a találat valószínűsége
 A V térfogaton belüli átlagos találatok száma cVD, c arányossági állandó, megmondja hány találatból hány ionizáció keletkezik. Véletlenszerű találatnál, n találat valószínűsége V térfogaton belül Poisson eloszlással adható meg. P(n)=e-cVD(cVD)nn! Single hit mechanizmusban a túlélők 1 találatot kell kapjanak S=1-P(n) S=e-cVD k=cV, azaz k arányos a cél térfogatával. Minél nagyobb a céltábla annál nagyobb a találat valószínűsége.")
26
k=cV; k arányos a cél térfogatával
A legnagyobb genomú fág a legérzékenyebb (T5) A k értékek aránya megegyezik a DNS molekulasúlyok arányával
 A k értékek aránya megegyezik a DNS molekulasúlyok arányával.")
27
DNS javító mechanizmusok
UV sugárzást tanulmányozták talán a legjobban DNS/fehérje abszorpciós maximuma 260/280nm Különböző hullámhosszú UV besugárzáskor a túlélési görbe hasonló kinetikájú, csak a meredeksége más Leghatékonyabb hullámhossz a 260 nm (ez a DNS elnyelési maximuma) UV sugárzás fő eredménye primidin dimer de főleg timin dimer (ciklo-butil-timin dimer)
 UV sugárzás fő eredménye primidin dimer de főleg timin dimer (ciklo-butil-timin dimer)")
28
Timin dimer hatásai DNS hélix torzul, a timinek közötti (szálon belüli) távolság csökken A torzulás eredménye, hogy a komplementer szálon lévő adeninekkel gyengül a hidrogénhíd kötés (maga a dimerizáció nem szünteti meg a H-híd kötés képességét) Ez a szerkezeti torzulás megakadályozza a replikációs villa továbbhaladását
 Ez a szerkezeti torzulás megakadályozza a replikációs villa továbbhaladását.")
29
Hogyan blokkolódik a replikáció?
A PolIII eléri a T dimert, a replikációs villa megakad A T dimer továbbra is képes H-híd kötésre, mert a dimerizáció nem érinti a H-híd kötésben résztvevő csoportokat A dimer torzulást visz a hélixbe Amikor a PolIII adenint szintetizál a szálra a T dimer miatt torzulás jön létre A PolIII úgy érzékeli, hogy rossz párok alakultak ki 3’-5’ exonukleáz aktivitással visszalép, kivágja a „rossz” nukleotidot A PolIII újra beépíti az adenint „helyben jár” A replikáció elakad, a baktériumok nem nőnek

30
Hogyan indulhat újra a replikáció?
A dimer direkt kijavítása fotoreakcióval Dimer kivágódhat, PolI beépíti a helyes bázispárt DNS szintézis a T dimer mögött újra indulhat így a T dimer rekombinációs javítással eltűnik SOS repair lehetővé teszi a hibás replikációt Melyek a bizonyítékok a javító mechanizmusok létére Baktériumoknál az UV besugárzás során nem volt egytalálatos túlélési görbe A DNS mindkét szálát kell érje találat? Kéttalálatos sem volt a túlélési görbe Nagyon nagy dózisok esetén sem volt lineáris a görbe

31
Mi lehet az oka, hogy a target elmélet nem működik
Hatékony javító mechanizmusok léteznek Kis dózis esetén nem elég hatékony a pusztítás A dózis emelkedésével a javító rendszer is telítődik, néhány találat javíthatatlan, vagy a repair rendszer is sérül Bármelyiknek eredménye az, hogy a túlélési görbe a kezdeti lapos rész után lefele görbül További megfigyelések A túlélő baktérium frakció növekedése, bizonyos besugárzás utáni kezelés eredményeképpen Olyan mutáns törzsek izolálása, melyek érzékenyebbek voltak a besugárzásra, mint a vad típus

32
Fotoreaktiváció UV kezelt tenyészet, ha fényen állt több túlélő sejtet tartalmazott, ez a fotoreaktiváció Biokémiai vizsgálatok kimutatják, hogy enzimes folyamat Az enzim fotoliáz T dimereket hasítja, mellyel a monomer állapot visszaáll Az enzim inaktív mindaddig, amíg nm hullámhosszú fény nem éri Az enzimhez asszociált folsav kofaktor elnyeli a fény energiáját Az enzim ezt az elnyelt energiát használja a kötés elbontására Phr- mutáns izolálása, mely nem fotoreaktiválódik és nincs fotoliáz enzimje Más fenotípusos változás nincsen

33
UV besugárzott fágok nem fotoreaktiválódnak, mert nincs fotoliázuk
Lehet azonban fotoreaktivációt elérni A fágot úgy kell adszorbeálni a baktérium sejtekre, hogy a baktériumokban gátolt legyen a szaporodás A fág DNS bejut a baktériumsejtben, de a szaporodás gátolt Fény hatására a baktérium fotoliáza kijavítja a fág DNS-ét is, gazda sejtes reaktiváció (host cell reactivation)
")
34
Sötét javítás liquid-holding javítás
Az UV fénnyel besugárzott sejteket nem tápanyagforrású pufferben több óráig tartva (sötétben) a túlélők száma növekszik Phr- mutánsban is van liquid-holding recovery Fény nélkül megy végbe Két rendszer tehát az E. coliban Fotoreaktiváció Fénytől független javító rendszer Alátámasztás Mutáns E. coli Bs törzs, amely UV érzékeny Normál fotoreaktiváció Sötétben (liquid holding reaktiváció) nincs coli Bs törzsben nincs gazdasejtes reaktiváció, és az UV besugárzott T1 fág túlélési görbéje meredekebb a Bs törzsben
 a túlélők száma növekszik. Phr- mutánsban is van liquid-holding recovery. Fény nélkül megy végbe. Két rendszer tehát az E. coliban. Fotoreaktiváció. Fénytől független javító rendszer. Alátámasztás. Mutáns E. coli Bs törzs, amely UV érzékeny. Normál fotoreaktiváció. Sötétben (liquid holding reaktiváció) nincs. coli Bs törzsben nincs gazdasejtes reaktiváció, és az UV besugárzott T1 fág túlélési görbéje meredekebb a Bs törzsben.")
35
Sötét javítási rendszerben mutáns coli előállítása (Bs elérhető volt, de genetikai analízis akkor még nem volt baktériumokra) K12 törzsből a host cell reaktivációt használva UV besugárzott T1 fágot és mutagenizált colit szélesztettek Ahol működik a gazdasejtes reaktiváció, ott a T1 fág növekszik és a baktérium elpusztul Ahol nem volt gazdasejtes reaktiváció ott a baktérium növekedhetett (a T1 nem javítódik) A kinőtt baktériumokat tesztelték UV érzékenységre
 A kinőtt baktériumokat tesztelték UV érzékenységre.")
36
A mutánsok komplementációjával 3 osztályt kaptak
uvrA, uvrB, uvrC Biokémiai analízis azt mutatta, hogy defektívek a T dimert kivágó endonukleázra Más mutánsok (recA, recB, recC mutánsok) esetén azt tapasztalták, hogy a recA- UV érzékeny, de a T dimereket javították DNS replikációban (PolI) hiányos mutáns szintén UV érzékeny
 esetén azt tapasztalták, hogy a recA- UV érzékeny, de a T dimereket javították. DNS replikációban (PolI) hiányos mutáns szintén UV érzékeny.")
37
A T dimerek eltávolításáért E. coliban 4 fő út felelős
Fotoreaktiváció Sötét javító mechanizmus Excíziós (kivágásos) javítás Rekombinációs javítás SOS javítás, SOS válasz (károsodás) Excíziós javítás A legtöbb javítás alapja a T dimer eltávolítása a DNS-ből Nem fotoreaktiváció Bizonyíték UV besugárzott baktérium Liquid holding reaktiváció T dimerek száma csökken (biokémiai vizsgálattal) T dimerek jelennek meg a sejten belül és kívül
 javítás. Rekombinációs javítás. SOS javítás, SOS válasz (károsodás) Excíziós javítás. A legtöbb javítás alapja a T dimer eltávolítása a DNS-ből. Nem fotoreaktiváció. Bizonyíték. UV besugárzott baktérium. Liquid holding reaktiváció. T dimerek száma csökken (biokémiai vizsgálattal) T dimerek jelennek meg a sejten belül és kívül.")
38
A kivágásos javítás több enzimes lépésből álló folyamat
Kétféle módszer az első bevágásnál coli T dimer két végénél vág (8 nukleotiddal az 5’ vég felé és 3-5 nukleotiddal a 3’ vég felé M. luteus T (pirimidin) dimer glikoziláz Kivágódik a T dimer rész PolI szálkicserélődés Egyszálú rész nukleáz hasítás Ligáz összekapcsolja a szálakat
 dimer glikoziláz. Kivágódik a T dimer rész. PolI szálkicserélődés. Egyszálú rész nukleáz hasítás. Ligáz összekapcsolja a szálakat.")
40
coliban a bemetszést az Uvr rendszer végzi
UvrA köti a torzult hélixet UvrB köt a fenti komplexhez UvrC a fenti komplexhez kötődve hasítja a DNS-t Az Uvr rendszer más javítást is végez T dimer Bázis eltolódás Nagyobb molekula beépülése (interkaláció) Az enzim komplex valószínűleg a hélix torzulását ismeri fel
 Az enzim komplex valószínűleg a hélix torzulását ismeri fel.")
41
Methyl directed mismatch repair (olyan javítás, ahol a DNS kicsit torzul)
DNS replikáció PolIII editáló funkciója, rossz beépülés esetén visszalép egyet és kijavítja rossz bázist A mismatch repair még egy biztosíték a hibátlan replikációra Mi történik, ha mégis beépül egy rossz bázis Honnan tudja az enzim, hogy melyik szálon van a „rossz” bázis A szülői szál metilált, az utód szál (egy ideig) nem metilált A metilált szál szolgál templátul mutS, mutL, mutH gének mutánsai növelik a spontán mutáció arányát dam gén adenin metiláz GATC >GA*TC, ha hibás nem működik a rendszer
 nem metilált. A metilált szál szolgál templátul. mutS, mutL, mutH gének mutánsai növelik a spontán mutáció arányát. dam gén adenin metiláz GATC >GA*TC, ha hibás nem működik a rendszer.")
42
A MutS köti az apró torzulást tartalmazó DNS darabot
A MutH két molekulája a MutS-hez köt, a DNS átcsúszik és egy hurok jön létre Addig csúszik a DNS, amíg a MutH hemimetilált GATC-hez nem érkezik (mindkét oldalon) A nem metilált szál degradálódik Az új szálat a PolIII szintetizálja A komplexben a MutL is ott van, de funkciója nem ismert
 A nem metilált szál degradálódik. Az új szálat a PolIII szintetizálja. A komplexben a MutL is ott van, de funkciója nem ismert.")
43
Rekombinációs javítás
Sejtések Ha a sötét javításért csak az excíziós javítás felel, akkor kevés T dimer is letális lenne, ezzel szemben T dimer kell a letális dózishoz, azaz van más repair mód is recA mutánsok UV érzékenyek Ráadásul a mindkét rendszerben mutánsok (uvr+recA) érzékenyebbek, mint az egyszeres mutáns T dimer kialakul Replikációs villa megakad Tovább indul a dimer után Ez a poszt-dimer iniciáció, a DNS replikáció újra indul Az utód szálon folytonossági hiányok a T dimer után (mintha Okazaki fragmensek lennének) Ez a folyamat azonban még nem elég a normális replikációhoz, mert az utód szálon hiányok vannak
 érzékenyebbek, mint az egyszeres mutáns. T dimer kialakul. Replikációs villa megakad. Tovább indul a dimer után. Ez a poszt-dimer iniciáció, a DNS replikáció újra indul. Az utód szálon folytonossági hiányok a T dimer után (mintha Okazaki fragmensek lennének) Ez a folyamat azonban még nem elég a normális replikációhoz, mert az utód szálon hiányok vannak.")
44
Sister strand exchange
A defektustól mentes egyszálú DNS kerül a kivágott T dimer helyére Ehhez a RecA fehérjére van szükség PolI szintetizálja az új szálat A ligáz összekapcsolja a fragmenseket A donor molekulából kivágódott egyszálú szakaszt a PolI javítja, a ligáz összekapcsolja Ha minden dimer kijavításra kerül 2 komplett szál keletkezik Probléma, ha a két komplementer (ellentétes) szálon, egymáshoz közel vannak T dimerek, akkor nincs jó donor molekula
 szálon, egymáshoz közel vannak T dimerek, akkor nincs jó donor molekula.")
45
két szál közötti keresztkötést az UvrABC rendszer és rekombinációs repair együtt oldja meg
A rekombinációs javítás előnye, hogy A replikáció tovább folytatódhat, nem kell megvárni, amíg minden T dimer javításra kerül Olyan hibák is javíthatók, melyek nem okoznak nagy hélix torzulást Excíziós javítással szemben ez poszt-replikatív vagy daughter strand javításnak hívják Fágoknál is létezik: recA- törzsön, az UV besugárzott fágok rosszabbul szaporodnak

46
Kimutatták, hogy az UV besugárzás erős mutagén
Az eddigi rendszerek azonban tökéletesen kijavították a hibákat, nincs mutagén hatás Az UV mutagenezis nem arányos a dózissal, csak nagyon erős UV dózis okoz mutagén hatást Ha az UV dózis során a T dimerek száma túllépi a javító mechanizmus kapacitását, egy másik rendszer lehetővé teszi a túlélést, de annak árán, hogy mutáció(k) jön(nek) létre Ez az SOS válasz Legutolsó mentsvár Lehetővé teszi a replikációt A ki nem javított DNS indukálja ezt a választ Az SOS elkerülő válaszreakció A DNS szál replikálódik, de hibákat halmoz fel A legtöbb mutagén hatásért ez a rendszer a felelős
 jön(nek) létre. Ez az SOS válasz. Legutolsó mentsvár. Lehetővé teszi a replikációt. A ki nem javított DNS indukálja ezt a választ. Az SOS elkerülő válaszreakció. A DNS szál replikálódik, de hibákat halmoz fel. A legtöbb mutagén hatásért ez a rendszer a felelős.")
47
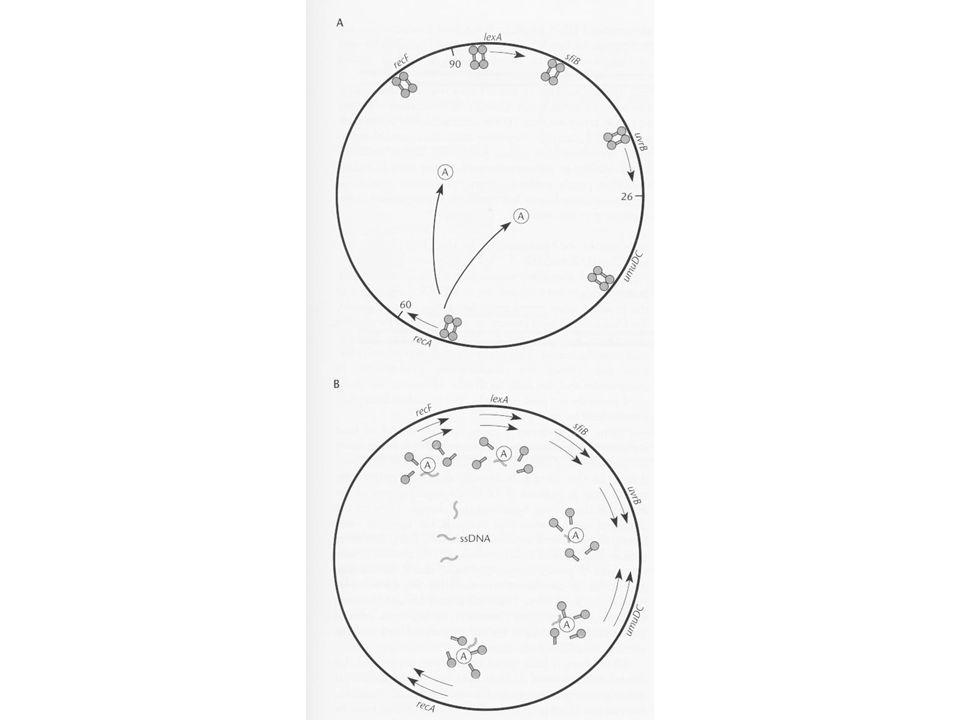
A SOS válasz szabályozása
SOS válasz kb. 20 gén koordinált szabályozott működését jelenti kiterjedt DNS károsodás esetén A lambda indukció és RecA fehérje közötti kapcsolat felfedezése volt a kulcs az SOS felfedezéséhez A mérsékelt fág lizogén és lítikus ciklusa Profág integrálódik a gazda kromoszómájába A baktérium amely hordozza a profágot a lizogén A profág a cI represzort termeli A represszor molekula megakadályozza, hogy a fág lítikus ciklusba kerüljön UV fény hatására azonban indukálódik a fág Ez a lizogén indukció, a fág replikálódik, megöli a gazda sejtet, lizálja és kiszabadul A süllyedő hajó esete A fág érzékeli, hogy a gazda elpusztul Még mielőtt a gazda elpusztul kivágja magát, replikálódik, kiszabadul és újabb gazdasejtet fertőzhet, azaz túlél Lizogén indukció csak nagy UV dózisnál fordul elő, amely indukálja az SOS választ, tehát a lambda fág indukciója része az SOS válaszreakciónak

48
A recA mutáns sejtek különösen érzékenyek az UV fényre
A recA-ra szükség van a rekombinációs hatásnál A rekombinációban mutáns recBC sejtek UV tűrők A recA-nak szerepe van más UV javításban is A recA mutánsokban az UV nem volt mutagén (nagyobb dózisra több sejt pusztult el, de a mutációk száma nem nőtt) A recA-ra szükség van az SOS válaszban recA mutánsokban nem lehet SOS választ kiváltani recA mutánsokban nem lehet UV fénnyel lambda fágot indukálni Vizsgálták a lambda cI represszort UV besugárzás után a cI represszor kettéhasadt recA mutánsban a cI intakt recA proteáz aktivitással rendelkezik Kimutatták, hogy a RecA fehérje egyszálú DNS és ATP kötése után átalakul proteáz formájúvá RecA*, a RecA* bontja a cI represszort, a profág indukálódik Később kiderült, hogy a RecA* nem proteáz, hanem csak elősegíti a cI represszor autokatalitikus aktivitását
 A recA-ra szükség van az SOS válaszban. recA mutánsokban nem lehet SOS választ kiváltani. recA mutánsokban nem lehet UV fénnyel lambda fágot indukálni. Vizsgálták a lambda cI represszort. UV besugárzás után a cI represszor kettéhasadt. recA mutánsban a cI intakt. recA proteáz aktivitással rendelkezik. Kimutatták, hogy a RecA fehérje egyszálú DNS és ATP kötése után átalakul proteáz formájúvá RecA*, a RecA* bontja a cI represszort, a profág indukálódik. Később kiderült, hogy a RecA* nem proteáz, hanem csak elősegíti a cI represszor autokatalitikus aktivitását.")
49
A RecA szerepe az SOS válaszban
A RecA* fehérje elősegíti az SOS válaszban szereplő más fehérjék autokatalitikus aktivitását A RecA szerepe az SOS válaszban Direkten gátolja a polIII editáló funkcióját, mert erősen köt a pirimidin dimer által torzított DNS szálhoz A polIII eléri a dimert, amelyhez RecA kapcsolódott A RecA kölcsönhatásba lép a polIII exonukleáz alegységével Gátlódik az editáló funkció A replikációs villa tovább halad (UV hatására a T dimerrel szemben adeninek épülnek be, de a torzulás miatt más is beépülhet A hiba nem editálódik, és a mutáció kialakul

50
Az umuDC gének termékei is szükségesek a hibával terhelt (error prone) replikációhoz (az abban mutánsok nem mutagenizálódnak) Az aktivált RecA fehérje elősegíti az UmuD fehérje autokatalitikus bomlását (UmuD’ keletkezik, aktív C-terminális rész) UmuD’+UmuC a RecA fehérjével együtt hozzájárulnak a hibás polimerizációhoz UmuDC gyakran plazmidon található UmuDC nélkül az UV besugárzás nem mutagenizál
 UmuD’+UmuC a RecA fehérjével együtt hozzájárulnak a hibás polimerizációhoz. UmuDC gyakran plazmidon található. UmuDC nélkül az UV besugárzás nem mutagenizál.")
51
SOS indukció DNS károsodásra indukálódik
UV reaktiváció jelensége (W, Weigle reaktiváció) Nagy dózisú UV-vel besugárzott fágot titrálunk UV besugárzott baktériumon > több plakk UV nem besugárzott baktériumon > kevesebb plakk A túlélő fágok DNS-e mutációkat tartalmazott UV besugárzás aktiválja az SOS választ (csak recA+) Mivel hibával terhelt az SOS válasz, ezért be/ki kapcsolhatónak kell lennie, hogy a hibák ne halmozódjanak fel A válasz csak bizonyos DNS sérüléseknél kell Néhány jellemzőnek kell indukálnia a javítást, de az összes SOS gént kell bekapcsolnia (ez a regulon, azaz olyan operonok készlete, amelyek közösen szabályzódnak)
 Nagy dózisú UV-vel besugárzott fágot titrálunk. UV besugárzott baktériumon > több plakk. UV nem besugárzott baktériumon > kevesebb plakk. A túlélő fágok DNS-e mutációkat tartalmazott. UV besugárzás aktiválja az SOS választ (csak recA+) Mivel hibával terhelt az SOS válasz, ezért be/ki kapcsolhatónak kell lennie, hogy a hibák ne halmozódjanak fel. A válasz csak bizonyos DNS sérüléseknél kell. Néhány jellemzőnek kell indukálnia a javítást, de az összes SOS gént kell bekapcsolnia (ez a regulon, azaz olyan operonok készlete, amelyek közösen szabályzódnak)")
52
Amikor sérülés van, az SOS bekapcsolt állapotban van hogyan:?
Az SOS szabályzó rendszernek két összetevője van a lexA és recA géntermékek A lexA gén az összes SOS operon represszora, a LexA fehérje az összes gén előtt található közös operátorra köt, és gátolja az expressziót A LexA a lexA operátorához is kötődik, azaz saját magát is szabályozza A RecA fehérje bekapcsolja az SOS választ a LexA fehérje autokatalízisének elősegítésével (mint a cI represszor) A DNS sérülés konformációs változást okoz a RecA fehérjében (RecA*) ez segíti a LexA autokatalízisét Amikor sérülés van, az SOS bekapcsolt állapotban van hogyan:? Sérülés RecA->RecA* A LexA érzékeny a proteolízisre lexA saját magát szabályozza
 A DNS sérülés konformációs változást okoz a RecA fehérjében (RecA*) ez segíti a LexA autokatalízisét. Amikor sérülés van, az SOS bekapcsolt állapotban van. hogyan: Sérülés RecA->RecA* A LexA érzékeny a proteolízisre. lexA saját magát szabályozza.")
53
Normál sejtben UV hatására a sejtben 50x aktivitás változás
RecA normális nem segíti a LexA bomlását LexA nem bomlik LexA represszálja az SOS génjeit LexA represszálj a lexA gént UV hatására a sejtben A RecA egyszálú DNSt köt (a T dimereknél felnyílt rész) RecA>RecA* RecA* segíti a LexA autokatalízisét LexA elhasad SOS géneket nem represszálja a LexA lexA gén is derepresszál, indukálódik 50x aktivitás változás
 RecA>RecA* RecA* segíti a LexA autokatalízisét. LexA elhasad. SOS géneket nem represszálja a LexA. lexA gén is derepresszál, indukálódik. 50x aktivitás változás.")
55
Léteznek ún. speciális javító rendszerek
Mindezek általános javító rendszerek voltak (kivéve a fotoreaktivációt) Léteznek ún. speciális javító rendszerek Bázis dezamináció Reaktív oxigén károsodás Alkiláció Adaptív válasz Meg kell különböztetni a DNS sérülést és mutációt A DNS sérülés önmagában nem mutagén, csak akkor válik azzá, ha sérülés következtében olyan változás keletkezik a DNS-ben mely tovább öröklődik
 Léteznek ún. speciális javító rendszerek. Bázis dezamináció. Reaktív oxigén károsodás. Alkiláció. Adaptív válasz. Meg kell különböztetni a DNS sérülést és mutációt. A DNS sérülés önmagában nem mutagén, csak akkor válik azzá, ha sérülés következtében olyan változás keletkezik a DNS-ben mely tovább öröklődik.")
56
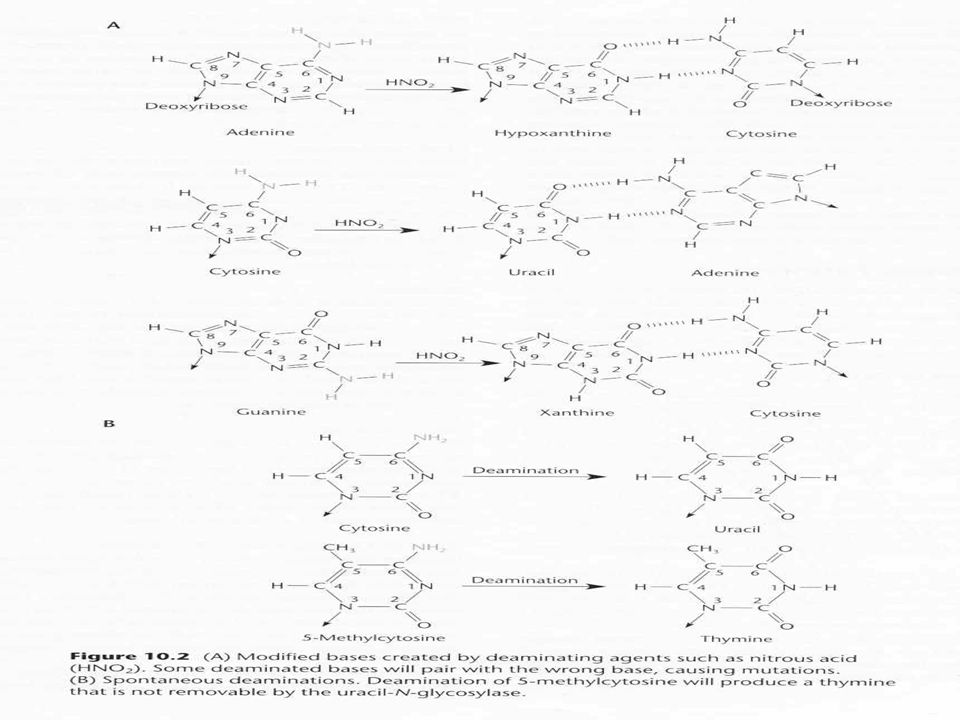
Bázisok dezaminálása A bázisokban található (adenin, guanin, citozin) amino csoportok sérülékenyek, spontán vagy kémiai behatásra eltávolíthatók Adenin dezaminálva hipoxantin Guanin dezaminálva xantin Citozin dezaminálva uracil Mutagén, mert rossz bázispárosodást okoz Pl adenin dezaminálódik, hipoxantin lesz belőle ez citozinnal párosodik AT ből CG tranzíció keletkezik Dezamináló ágensek Hidroxilamin (citozin) Biszulfit (citozin) Salétromos sav
 Biszulfit (citozin) Salétromos sav.")
58
Dezaminált bázisok javítása
DNS glikozilázok Hasítják a módosított bázist a nukleotid cukorjában Minden bázisra specifikus glikoziláz Amikor a bázis nincs a szálon, az AP (apurin, apirimidin) endonukleázok hasítják a cukor-foszfát láncot A hasítás után a szabad 3’OH indítja a PolI javítást coliban
 endonukleázok hasítják a cukor-foszfát láncot. A hasítás után a szabad 3’OH indítja a PolI javítást coliban.")
59
Very short patch repair (VSR) dezaminált 5 metil citozin javítása
Metil csoport védi a DNS-t restrikció ellen Magasabbrendűekben génszabályozó szerep Dezamináció során uracil helyett timin keletkezik, azaz mutagén hely A legtöbb metil citozin a 5’CCWGG3’/3’GGWCC5’ szekvenciában a második citozin (W=AT v. TA) A másik citozint a Dcm (DNS citozin metiláz metilálja) CmCAGG/GGTCmC szekvencia Ez sérülékeny, ezért a coli repair rendszere kijavítja ezt a szekvenciát, ha a Cm helyett T jelenik meg Ebben a rövid szekvenciában a vsr gén VSR endonukleáza a TG mismatchhez köt és a T mellett hasít, ezután PolI javítja a szálat Nagyon speciális csak ebben a szekvenciában javítja a TG mismatch-et
 A másik citozint a Dcm (DNS citozin metiláz metilálja) CmCAGG/GGTCmC szekvencia. Ez sérülékeny, ezért a coli repair rendszere kijavítja ezt a szekvenciát, ha a Cm helyett T jelenik meg. Ebben a rövid szekvenciában a vsr gén VSR endonukleáza a TG mismatchhez köt és a T mellett hasít, ezután PolI javítja a szálat. Nagyon speciális csak ebben a szekvenciában javítja a TG mismatch-et.")
60
Reaktív OXIGÉN-ek Ezek közül a legmutagénebb a 7,8-dihidro-8-oxoguanin (8-oxo-G, GO) Gyakran adeninnel párosodik mutM, mutY, mutT gének termékei játszanak szerpet MutM N-glikoziláz, mely specifikusan eltávolítja a GO bázist, a dezoxiribóz cukorról Itt a MutM el is hasítja a szálat más esetben exonukleáz hasítja MutY N-glikoziláz, de nem GO-t, hanem a vele szembe beépült adenint távolítja el, a többit más repair rendszer végzi el. MutT megakadályozza, hogy 8-oxoG kerüljön a DNSbe, a d8-oxoGTP-t hidrolizálja

61
Alkiláló ágensek Javító rendszerek
A bázisok és a foszfát is alkilálódhat Alkiláló ágensek alkil csoportot CH3, CH3CH2 ragasztanak a foszfát csoporthoz, vagy bázisokhoz Pl. etil-metán-szulfonát (EMS, mustágáz, metil-metánszulfonát, N-metil-N’-nitro-N-nitrozoguanidin, NTG) Guanin N7 és adenin N3 a legérzékenyebb Az alkilált bázis hajlamos rossz bázispárosodára Javító rendszerek Specifikus N-glikozilázok, az alkilált bázis levágódik, AP endonukleáz hasít, exonukleáz degradáció, PolI javítás alkA gén Metil transzferázok (O6 metil-guanin, O4 timin) Az alkil csoportot eltávolítják, miközben maguk metilálódnak Nem igazi enzimek, mert nem katalizálják a folyamatot, hanem felhasználják magukat
 Guanin N7 és adenin N3 a legérzékenyebb. Az alkilált bázis hajlamos rossz bázispárosodára. Javító rendszerek. Specifikus N-glikozilázok, az alkilált bázis levágódik, AP endonukleáz hasít, exonukleáz degradáció, PolI javítás alkA gén. Metil transzferázok (O6 metil-guanin, O4 timin) Az alkil csoportot eltávolítják, miközben maguk metilálódnak. Nem igazi enzimek, mert nem katalizálják a folyamatot, hanem felhasználják magukat.")
62
E. coli adaptálódik alkiláló ágensek hatására
NTG kezelés (kis mennyiség) után a sejt képes tolerálni nagyobb mennyiségű alkiláló ágenst Számos gén aktiválódik amelyek szerepet játszanak az alkiláló ágensek okozta sérülések javításában Szabályozás (néhány kópiától több ezerig) Az Ada protein metilációja a fő szabályozó Két helyen metilálódhat Az egyik hely O6-metilguanin, O4-metiltimin-től vesz fel metil csoportot, a másik foszfát csoportról Bármelyik helyen vesz fel metil csoportot több metil csoport nem kötődhet AdaC terminális részen metil csoport metG, metT-től, nem enged több metilációt önmagán>öngyilkos inaktiváció Az ada N terminális részen, ha felvesz metil csoportot a foszfát vázról, akkor az Ada fehérje aktiválja saját génjét, csakúgy, mint más adaptív válasz génekét.
 után a sejt képes tolerálni nagyobb mennyiségű alkiláló ágenst. Számos gén aktiválódik amelyek szerepet játszanak az alkiláló ágensek okozta sérülések javításában. Szabályozás (néhány kópiától több ezerig) Az Ada protein metilációja a fő szabályozó. Két helyen metilálódhat. Az egyik hely O6-metilguanin, O4-metiltimin-től vesz fel metil csoportot, a másik foszfát csoportról. Bármelyik helyen vesz fel metil csoportot több metil csoport nem kötődhet. AdaC terminális részen metil csoport metG, metT-től, nem enged több metilációt önmagán>öngyilkos inaktiváció. Az ada N terminális részen, ha felvesz metil csoportot a foszfát vázról, akkor az Ada fehérje aktiválja saját génjét, csakúgy, mint más adaptív válasz génekét.")
63
Összefoglalás Genom integritást védő mechanizmusok
Restrikció-modifikáció Restrikciós-modifikációs rendszerek Javító mechanizmusok DNS sérülések DNS repair folyamatok Általános javítófolyamatok Excíziós Methyl directed mismatch repair Rekombinációs repair SOS rendszer Speciális javítórendszerek Fotoreaktiváció Dezaminált bázisok javítása Very short patch repair Reaktív oxigén okozta károk javítása (oxo-guanin) Alkiláció (adaptív válasz)
 Alkiláció (adaptív válasz)")
64
MUTÁCIÓ

65
A korai mutáció következménye

66
Valószínűség Valószínűség: az, hogy valami történni fog, bekövetkezik. Például ma éjszaka csillagos lesz az ég. Definíció szerint 0,0 azt jelenti, hogy soha 1,0 azt jelenti, minden alkalommal bekövetkezik Mit jelent a 0,5 –es érték? Mi a valószínűsége, hogy fej lesz egy normális pénzdarab feldobásának eredménye? Mi a valószínűsége annak, hogy 3-ast dobunk egy kockával?

67
Nem független események
Mi a valószínűsége annak, hogy 4-est dobunk, majd 6-ost? Ha a mutációs ráta 10-6/sejtosztódásonként, mi a valószínűsége annak, hogy két mutáció lesz ugyanabban a sejtben? Független események Mi a valószínűsége annak, hogy vagy 4-est, vagy 6-ost dobunk? Mi a valószínűsége annak, hogy a trpA, vagy pedig a trpB génben lesz mutáció?

68
Permutáció Fontos az események sorrendje P=? 4, majd 6 P=? 6, majd 4
Két különböző permutáció

69
Kombináció Hány permutáció lehetséges , ha egy dobókockát kétszer dobunk el? 6 kimenetel az első dobásra 6 lehetőség a másodikra is 36 permutáció P=?, hogy két dobásra 4 és 6 Bármelyik két permutáció lehet: 4, 6; 6, 4 Ugyanaz a kombináció

70
Születésnap probléma Mi a valószínűsége annak, hogy itt a teremben két ember ugyanazon hónapban és ugyanazon a napon van a születésnapja? Nehéz kiszámítani Könnyebb az ellenkezőjét. Mi a valószínűsége annak, hogy nincs két azonos születésnap? 365/365 * 364/365 * 363/365…*353/365 = 78% Igy annak a valószínűsége, hogy két embernek ugyanaz a születésnapja (1–0,78 )*100 = 22%
*100 = 22%")
71
Binomiális eloszlás Gyakran két eseménnyel dolgozunk
Sikeres (S) / sikertelen (N) A sikeres kimenetelt önkényesen definiáljuk P (siker) + P (sikertelen) = 1,0 S + N = 1 S2 + 2SN + N2 = 1 (két független esemény) Általános öf: (S+N)n=1 , ahol n a próbálkozások száma Gyorsan bekövetkezik, hogy nehéz pontosan kiszámítani
 / sikertelen (N) A sikeres kimenetelt önkényesen definiáljuk. P (siker) + P (sikertelen) = 1,0. S + N = 1. S2 + 2SN + N2 = 1 (két független esemény) Általános öf: (S+N)n=1 , ahol n a próbálkozások száma. Gyorsan bekövetkezik, hogy nehéz pontosan kiszámítani.")
72
Poisson eloszlás A binomiális kiterjesztés közelítő meghatározása
Könnyebb vele számolni Csak akkor érvényes, ha a mintaszám n nagy és a siker valószínűsége, P(S) kicsi m = várható érték = np A siker r exakt értéke kiszámítható Pr=e–m mr/r! 0! =1
 kicsi. m = várható érték = np. A siker r exakt értéke kiszámítható. Pr=e–m mr/r! 0! =1.")
73
Mutációs ráta A fluktuációs teszt redményei felhasználhatók
A csövek hány százalékában nem volt mutáns? Ez lesz a Pr=0 Oldjuk meg az egyenletet m-re. Ha tudjuk szélesztett sejtek számát, akkor p kiszámítható, azaz a mutáció valószínűsége A mutációs ráta általában 10–6 - 10–8

74
Poisson eloszlás Target elmélet.
Px annak valószínűsége, hogy egy gént (target) pontosan x találat ér. h átlagos találatszám per target Px meghatározásának legegyszerűbb módja: nincs mutáció P0 értékének meghatározása és behelyettesítése a fenti egyenletbe.
 pontosan x találat ér. h átlagos találatszám per target. Px meghatározásának legegyszerűbb módja: nincs mutáció P0 értékének meghatározása és behelyettesítése a fenti egyenletbe.")
75
Mutációs ráta, mutáns gyakoriság
Mutációs ráta (a), összes sejt a tenyészetben (N), mutánsok száma (h) Mutációs rátát nehéz meghatározni, ezért sokkal gyakrabban a mutáns gyakoriságot adják meg = mutánsok aránya a tenyészetben A spontán mutációs ráta E. coli-ban 1/300 kromoszóma replikáció (hipermutabilitás!!)
, összes sejt a tenyészetben (N), mutánsok száma (h) Mutációs rátát nehéz meghatározni, ezért sokkal gyakrabban a mutáns gyakoriságot adják meg = mutánsok aránya a tenyészetben. A spontán mutációs ráta E. coli-ban 1/300 kromoszóma replikáció (hipermutabilitás!!)")
76
Példák a mutációs rátára
Emlősökben viszonylag konstans mutációs ráta sok génnél.

77
Szelekció és a lappangások
Fenotípusos lag (recesszív mutáció) Szegregációs lag (domináns mutáció) Mutáció a replikálódó DNSben Nincs mutáns fenotípus, mert a citoplazmában még van wt géntermék Először jelenik meg a mutáns fenotípus Kész a replikáció 1. Sejtosztódás A mutáns fenotípus akkor jelenik meg, ha a wt géntermék kellően kihígul, vagy eltűnik. A mutáns sejtek száma a következő osztódásnál kezd nőni. Vad típ sejtek 2. Sejtosztódás Mutáns sejtek száma nő
 Szegregációs lag. (domináns mutáció) Mutáció a replikálódó DNSben. Nincs mutáns fenotípus, mert a citoplazmában. még van wt géntermék. Először jelenik meg. a mutáns fenotípus. Kész a replikáció. 1. Sejtosztódás. A mutáns fenotípus akkor jelenik meg, ha a wt géntermék kellően kihígul, vagy eltűnik. A mutáns sejtek száma a következő osztódásnál kezd nőni. Vad típ sejtek. 2. Sejtosztódás. Mutáns sejtek. száma nő.")
78
Grafikus példa Sejtek az exponenciális fázisban szaporodnak Lag
Konjugációs géntranszfer

79
Mutáns dúsítás penicillinnel
Hogyan találjuk meg a hiánymutánst (auxotróf)? Közvetlen szelekció általában nem lehetséges Gorini penicillint (vagy származékait) használta A penicillin a sejtfalszintézist gátolja Az eredmény Protoplaszt G(+) sejt sejtfal nélkül Spheroplast G(–) sejt minimális sejtfallal Ha éheztetjük a szükséges tápanyagra, akkor a hiánymutáns biztonságban van, mert nem tud növekedni
 Közvetlen szelekció általában nem lehetséges. Gorini penicillint (vagy származékait) használta. A penicillin a sejtfalszintézist gátolja. Az eredmény. Protoplaszt G(+) sejt sejtfal nélkül. Spheroplast G(–) sejt minimális sejtfallal. Ha éheztetjük a szükséges tápanyagra, akkor a hiánymutáns biztonságban van, mert nem tud növekedni.")
80
A genetikai kód Univerzális? nagyjából
A mitokondriumokban kicsit különbözik UAA trp-t kódolhat a transzlációs STOP helyett A kód nagyon redundás Crick wobble hipotézis A harmadik bázis kis különbséget jelent Ha az első két bázisnál 6 H híd, akkor a harmadik irreleváns

81
Kód Az ORF start kodon ÁLTALÁBAN AUG, de lehet GUG is
Az első aminosav a metionin Csak egy kodon Ezért a szövegkörnyezet hatása Némely szignál közelében az AUG azt jelenti, hogy „itt kezdődik a transzláció” Az AUG máshol a metionint jelenti

82
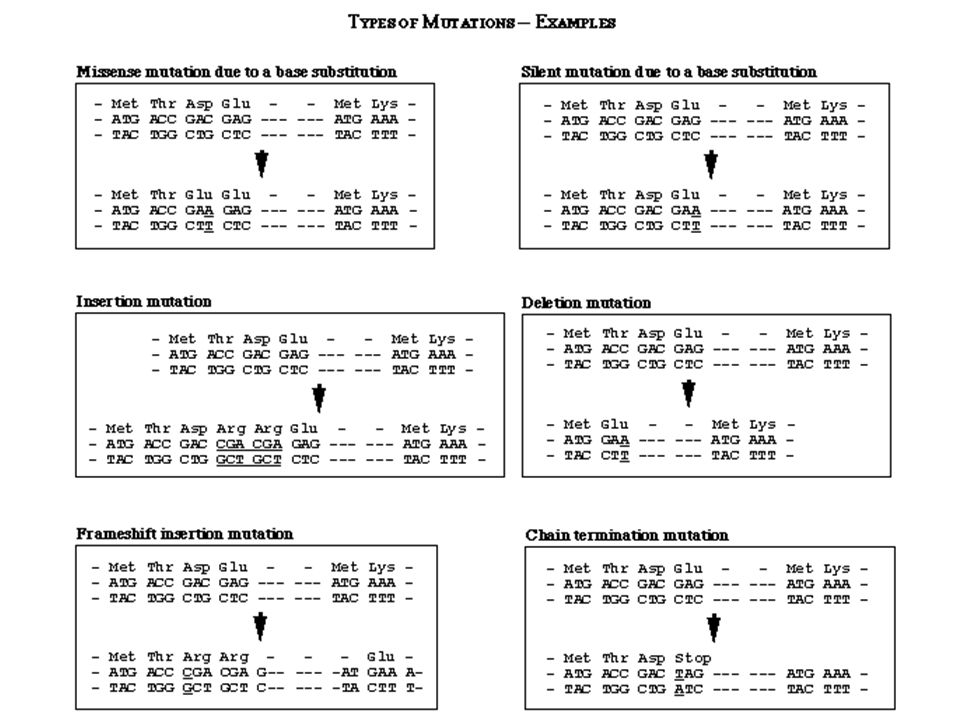
Mutáció típusok BÁRMILYEN változás a DNS szekvenciában mutáció
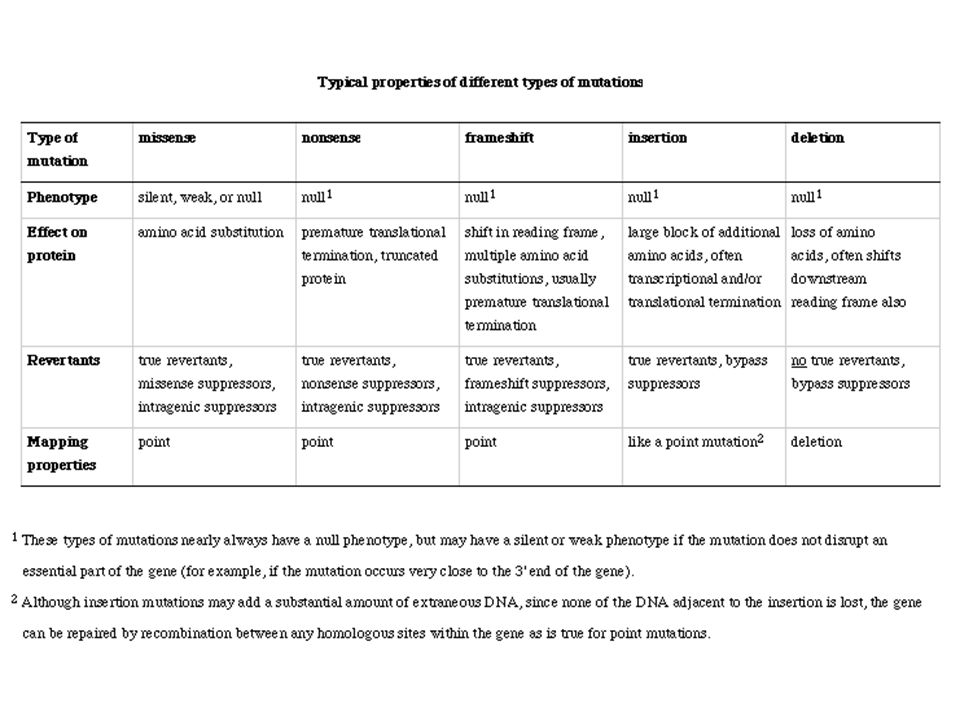
Szubsztitúciók Vad szekvencia AGC (Ser) Csendes AGT (Ser) Misszensz GGC (Pro) Nonszensz ATC (STOP) Deléció/Inzerció AGGC Inverzió CGA
 Csendes AGT (Ser) Misszensz GGC (Pro) Nonszensz ATC (STOP) Deléció/Inzerció AGGC. Inverzió CGA.")
83
Frameshift mutáció Minden 3-al nem osztható bázis addíció, vagy deléció sok sör árt sok srá rt… (1 bázisnyi deléció) A frameshift eredményeként downstream értelmetlenség 3-al osztható bázisnyi addíció, vagy deléció az adott helyen megváltoztatja az aminosavat, de downstream a kód változatlan bor nem sok sör árt

84
Szupresszor mutációk A második mutáció érvényteleníti az első hatását
Nem reverzió, hanem érvénytelenítés Lehet ugyanabban a génben (intragénes), vagy egy másik génben (intergénes) Intragénes +1 frameshift-et érvényteleníti egy –1 frameshift Rossz folding-ot kompenzálja egy másik változás Intergénes Általában tRNS mutáció A rossz kodonhoz a jó aminosavat szállítja
, vagy egy másik génben (intergénes) Intragénes. +1 frameshift-et érvényteleníti egy –1 frameshift. Rossz folding-ot kompenzálja egy másik változás. Intergénes. Általában tRNS mutáció. A rossz kodonhoz a jó aminosavat szállítja.")
85
Mutagének Kémiai, vagy fizikai → DNS sérülést okoznak
Hiba a repair folyamatokban Többé kevésbé véletlen Szokatlan típusok transzpozonok mutátor mutációk

88
Mutagén Mechanizmus Okozott mutáció típusa Spontán DNS replikáció és repair hiba, a nukleotidok spontán modifikációja Minden mutáns típus UV sugár Primidin dimer, error prone repair-t (SOS) indukál Főleg G-C → A-T tranzíció, de minden más is, deléció, frameshift és kromoszóma átrendeződés, de kisebb gyakorisággal 2-amino-purin (2AP) Bázis analóg A-T →G-C és G-C → A-T tranzíció 5-bróm-uracil G-C → A-T és A-T → G-C tranzíció Hidroxi-lamin (NH2OH) Alkiláló ágens, N4-hidroxicitozint készít G-C → A-T tranzíció in vitro használva
 indukál. Főleg G-C → A-T tranzíció, de minden más is, deléció, frameshift és kromoszóma átrendeződés, de kisebb gyakorisággal. 2-amino-purin (2AP) Bázis analóg. A-T →G-C és G-C → A-T tranzíció. 5-bróm-uracil. G-C → A-T és A-T → G-C tranzíció. Hidroxi-lamin (NH2OH) Alkiláló ágens, N4-hidroxicitozint. készít. G-C → A-T tranzíció in vitro használva.")
89
N-metil-N'-nitro-N-nitrozoguanidin (MNNG)
Alkilez, O6-metilguanint készít G-C → A-T tranzíciók, többszörös, közeli mutációk, Rep. villában Etilmetán szulfonát (EMS) G-C → A-T tranzíció Etiletán Szulfonát (DES) SOS választ indukál G-C → T-A transzverzió, más báziszubsztitúciós mutációk Salétromos sav Oxidatív dezaminálás G-C→ A-T és A-T →G-C tranzíciók, deléciók kisebb gyakorisággal ICR-191 Interkalálódó, akridin származék a kihurkolódott bázisokat közé beépülve stabilizál Frameshift, többnyire addíció, vagy deléció a G, vagy C ismétlődéseknél
 G-C → A-T tranzíció. Etiletán. Szulfonát. (DES) SOS választ indukál. G-C → T-A transzverzió, más báziszubsztitúciós mutációk. Salétromos sav. Oxidatív dezaminálás. G-C→ A-T és A-T →G-C tranzíciók, deléciók kisebb gyakorisággal. ICR-191. Interkalálódó, akridin származék a kihurkolódott bázisokat közé beépülve stabilizál. Frameshift, többnyire addíció, vagy deléció a G, vagy C ismétlődéseknél.")
90
Helyspecifikus mutagenezis Site-Directed Mutagenesis
Egyszálú komplementer primer szándékos mismatch Egyszálú templát Nukleotidok a szintézishez: dATP, dGTP, dTTP, 5-metil-dCTP T7 DNS polimeráz és DNS ligáz methyl- -methyl Nem metilezett DNS vágása MspI enzimmel. Fragmantumok eltávolítása exoIII-mal. Klónozás, vagy elektroporáció.

91
tranzíció, transzverzió, frameshift
Repair rendszer mutáns Mutációs gyak. növekedés Fő mutáció típusok Mismatch repair mutHSL tranzíció, frameshift Proofreading mutD tranzíció, transzverzió, frameshift G-A mismatch repair mutT AT → CG transzverzió mutY > 102 GC→TA transzverzió mutD = dnaQ, DNS pol III ε alegység, proofreading

92
Mutagén dózis Mutagének alkalmazásakor figyelembe venni:
Mutagén kémiai reaktivitása a DNS-sel A mutagén és a cél DNS koncentrációja A kezelés időtartama Többszörös mutánsok kerülendők (Poisson) Letális mutációk Fenotípusos lag-ra figyelni szelektív táptalajra szélesztés előtt
 Letális mutációk. Fenotípusos lag-ra figyelni szelektív táptalajra szélesztés előtt.")
93
A mutagén dózissal nő a kívánt mutánsok száma, de a sejtek pusztulása is, ezért a keresett mutás száma csökken. Egyensúly!!!



 DNS-ből,>")







 prokariótában>")





>")




