Előadást letölteni
1
EXPRESSZIÓS RENDSZEREK
Buday László

2
cDNS EXPRESSZIÓS KÖNVTÁR
Leggyakrabban a λgt11 vektort használják expressziós könyvtár készítésére EcoRI lac promoter β-galaktozidáz λgt11 vektort A vektor és a cDNS inzert emésztése EcoRI-gyel β-galaktozidáz cDNS inzert EcoRI-gyel emésztve ligálás cDNS β-galaktozidáz

3
Molecular Cell Biology
Lodish, 2000, W.H. Freeman and Company

4
MILYEN MÓDON LEHET A FEHÉRJÉKET AZONOSÍTANI?
Általános eljárás: immunológiai kimutatás specifikus ellenanyagok segítségével (elsődleges és jelzett másodlagos ellenanyagok felhasználásával) Eseti eljárás: a vizsgálandó fehérjék természete alapján 1, Oligonukleotid próba segítségével – ha a fehérje DNS-kötő (pl. transzkripciós faktorok esetén) 2, Fehérje-fehérje interakció alapján (pl. kalmodulin-kötő fehérjék kimutatása 125I-dal jelzett kalmodulinnal) 3, jelzett kis organikus molekulák segítségével (pl. jelzett cAMP) 4, Foszfotirozinra specifikus ellenanyag segítségével tirozinon foszforilált fehérjéket lehet kimutatni
 Eseti eljárás: a vizsgálandó fehérjék természete alapján. 1, Oligonukleotid próba segítségével – ha a fehérje DNS-kötő. (pl. transzkripciós faktorok esetén) 2, Fehérje-fehérje interakció alapján (pl. kalmodulin-kötő fehérjék. kimutatása 125I-dal jelzett kalmodulinnal) 3, jelzett kis organikus molekulák segítségével (pl. jelzett cAMP) 4, Foszfotirozinra specifikus ellenanyag segítségével tirozinon. foszforilált fehérjéket lehet kimutatni.")
6
A FEHÉRJÉK EXPRESSZIÓJÁNAK LEHETSÉGES MÓDJAI
In vivo: 1, prokarióta expressziós rendszer 2, eukariota expressziós rendszer - élesztő SF9 sejtek –bakulovírus Drosophila emlős sejtek a, tranziens b, stabil c, indukálható 3, transzgenikus állat 4, humán génterápia In vitro transzláció

7
PROKARIÓTA EXPRESSZIÓS RENDSZEREK
Igen gyakran az E.coli-ban expresszálni kívánt fehérjét fúziós fehérjeként termeltetik, ennek okai: -- nem a fehérje baktériumban betöltött szerepét vizsgálják -- emlős fehérje önmagában gyakran rosszul expresszálódik -- a bakteriális fehérje vagy más alsóbbrendű élőlény fehérjéje (β-galaktozidás, glutation-S-traszferáz v. GST) viszont jól expresszálódik, így nagy a valószínűsége, hogy a vele fúzionált fehérje is jól fog expresszálódni -- lehetőleg egy lépésben lehessen a baktérium kivonatából a vizsgált fehérjét megtisztítani
 viszont jól. expresszálódik, így nagy a valószínűsége, hogy a vele. fúzionált fehérje is jól fog expresszálódni. -- lehetőleg egy lépésben lehessen a baktérium kivonatából a vizsgált. fehérjét megtisztítani.")
8
EGY ADOTT GÉN BAKTERIÁLIS EXPRESSZIÓJÁNAK SÉMÁJA
SD szekvencia= Shine-Delgarno szekvencia, ami szükséges a mRNS bakteriális riboszómákhoz való kötődéséhez The Cell G.M. Cooper, 1997 ASM Press

9
BAKTERIÁLIS FÚZIÓS FEHÉRJE RENDSZEREK
1, Glutation-S-transzferáz: 26 kDa fehérje Schistosoma japonica génterméke pGEX vektor-sorozat gyors izolálás glutation-agaróz gyantán 2, Maltóz-kötő fehérje: E. coli malE génterméke pMEL vektor-sorozat izolálás maltóz affinitás oszlopon 3, Tioredoxin 17 kDa fehérje, nagyon szolubilis, hőstabil Ribonukleotid-reduktáz redukáló enzim E. coli trxA gén terméke pTrxFus vektor

10
pGEX GLUTATION-S-TRANSZFERÁZ (GST) EXPRESSZIÓS RENDSZER
--- Tetszőleges fehérje, vagy fehérje domén nagy mennyiségben E. coli-ban történő expresszálására Polilinker v. Multicloning site Lac promóter IPTG GST Represszor fehérje pGEX Lac inhibitor gén Ampicillin rezisztencia gén Ori

11
pGEX, azaz GST vektorok az Amersham-Pharmacia cégtől

12
Restrikciós enzimek, illetve a trombin hasítási helye
a pGEX plazmidban

13
EUKARIÓTA EXPRESSZIÓS RENDSZEREK I. BAKULOVÍRUS/SF9 SEJTEK
-- bakteriális expressziós rendszerekben nem történik meg az eukarióta fehérjék ún. poszt-transzlációs módosulása v. processzálása pl. fehérje foszforiláció, acetiláció, izopreniláció, palmitoiláció stb. -- viszont ezek a módosulások számos esetben nélkülözhetetlenek az expresszálni kívánt fehérje biológiai aktivitásához ban közölték a rendszert először Miller et al. -- A bakulovírus genomja nagy ~ 130 kb, tehát nagy DNS darabokat lehet benne kifejezni, valamint rekombináció segítségével könnyű a kívánt gént a vírus genomba jutattni -- a bakulovírus gerinceseket nem fertőz meg!!!

14
A BAKULOVIRUSNAK A FERTŐZÉST KÖVETŐEN KÉT
FORMÁJA LÉTEZIK Polihedrin fehérje (29 kDa) a sejtben maradó vírus által termelt fehérje, mely az okklúziós testek fehérje tartalmának több mint 50%-áért felelős. sejt mag Sejtmagban található vírus, ami az okklúziós testekben v. polihedrában található a ferőzést követő 72 óráig Extracelluláris vírus
 a sejtben maradó vírus által termelt fehérje, mely az okklúziós testek fehérje tartalmának több mint 50%-áért felelős. sejt. mag. Sejtmagban található. vírus, ami az okklúziós. testekben v. polihedrában. található a ferőzést követő. 72 óráig. Extracelluláris vírus.")
15
REKOMBINÁNS VÍRUS LÉTREHOZÁSA
bakulovírus genomja Polihedrin intakt polihedrin gén promóter Az expresszálni kívánt gént a Polihedrin gén helyére lehet REKOMBINÁCIÓ segítségével bejuttatni

17
Spodoptera frugiperda

18
BAKULOVÍRUS-SF9 RENDSZERBEN TERMELT FEHÉRJE
nagy mennyiségben expresszálódó fehérje bakulovírus receptor mag SF9 sejtek, azaz hernyó bélhámsejtek (27º C-on nőnek) A sejtek feltárását követően a kivonatból lehet a vizsgált fehérjét tovább tisztítani
 A sejtek feltárását követően. a kivonatból lehet a vizsgált. fehérjét tovább tisztítani.")
19
EMLŐS EXPRESSZIÓS RENDSZEREK
Rekombináns gének expressziója emlős sejtekben több, a gén- expresszió szabályozásában alapvető elem meglétét igényli: 1, promóter (minél több sejtben működjön) 2, enhanszer 3, poliadenilációs szignál 4, intron jelenléte fokozhatja az expresszió hatásfokát 5, szelekciós marker (ampicillin, neomicin, stb.) Promóter leggyakrabban: simian vírus 40 (SV40) (erős promóterek) papovavírus Rous sarcoma vírus human citomegalovírus (CMV) Expresszió formája lehet: konsitutív: tranziens vagy stabil indukálható
 2, enhanszer. 3, poliadenilációs szignál. 4, intron jelenléte fokozhatja az expresszió hatásfokát. 5, szelekciós marker (ampicillin, neomicin, stb.) Promóter leggyakrabban: simian vírus 40 (SV40) (erős promóterek) papovavírus. Rous sarcoma vírus. human citomegalovírus (CMV) Expresszió formája lehet: konsitutív: tranziens vagy stabil. indukálható.")
20
pSI EMLŐS EXPRESSZIÓS VEKTOR (PROMEGA)
Konstitutív expresszióra alkalmas, SV40 promóter és enhanszer, polilinker, SV40 late poly(A), ampicillin rezisztencia, T7 RNA polimeráz promóter in vitro transzkripcióra alkalmas
, ampicillin rezisztencia, T7 RNA polimeráz promóter in vitro. transzkripcióra alkalmas.")
21
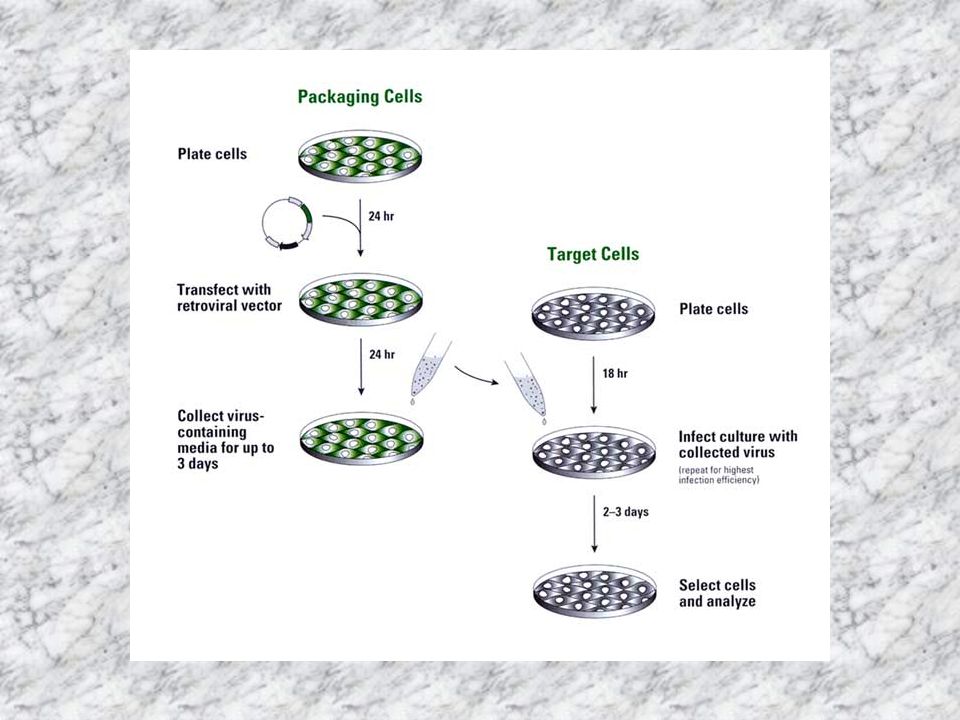
VIRÁLIS EXPRESSZIÓS RENDSZEREK EMLŐS SEJTEKBEN
ADENOVÍRUS expressziós rendszer: emberi sejtekben a legmagasabb fehérje expressziót biztosítja nem integrálódik a vírus DNS a célsejt genomjába osztódó és nem-osztódó sejteket egyaránt fertőzhetőek jelenleg a humán génterápia egyik legtöbbet használt vektora RETROVÍRUS expressziós rendszer: RNS vírusok, 100%-os hatásfokkal fertőznek a vírus genom integrálódik a sejt genomjába csak osztódó sejteket fertőz

23
TRANSZFEKCIÓ LEHETSÉGES MÓDJAI
1, DEAE-dextrán 2, Kálcium-foszfát 3, Lipofekciós – kationos liposzómák segítségével 4, Elektroporáció 5, Mikroinjekció 6, RNS és DNS vírusok által ( in vivo génterápia alapja) Az kölünféle sejttípusok más és más transzfekciós eljárást „szeretnek”.
 Az kölünféle sejttípusok más és más transzfekciós eljárást „szeretnek .")
24
INDUKÁLHATÓ EXPRESSZIÓS RENDSZEREK
1, Szteroid hormon indukció: adenovírus promóter glucocorticoid response element (GRE) indukció: dexametazon 2, Tetraciklin operon: CMV promóter Tet operátor szekvencia, tet represszor fehérje indukció: tetraciklin 3, Ektizon-indukálta rendszer: SV40 promóter, 2 vektor kell hozzá humán retinoid X receptor (RXR) és a Drosophila Ektizon Receptor (VgEcR) alkot egy funkcionális komplexet (transzkip- ciós faktor heterodimer), mely aktiválható ponasterone A adásával dózis-függő expresszió érhető el vele!
 indukció: dexametazon. 2, Tetraciklin operon: CMV promóter. Tet operátor szekvencia, tet represszor fehérje. indukció: tetraciklin. 3, Ektizon-indukálta rendszer: SV40 promóter, 2 vektor kell hozzá. humán retinoid X receptor (RXR) és a Drosophila. Ektizon Receptor (VgEcR) alkot egy funkcionális komplexet (transzkip- ciós faktor heterodimer), mely aktiválható ponasterone A adásával. dózis-függő expresszió érhető el vele!")
25
TETRACIKLIN-INDUKÁLTA EXPRESSZIÓS RENDSZER I.
TetO2 = tetracilkin operator szekvencia TetR = tetraciklin represszor fehérje

26
TETRACIKLIN-INDUKÁLTA EXPRESSZIÓS RENDSZER II.

27
RIPORTER GÉNEK A gén-expresszió szabályozását (promóter,enhanszer, stb.) riporter gének segítségével lehet többek között vizsgálni. Itt az expresszálódó fehérje egy enzim, melynek enzimaktivitásából lehet következtetni a gén-expresszió szabályozására. Leggyakrabban használt reporter enzimek: CAT (chlofarfenikol acil-transzferáz, bakteriális) β-galaktozidáz (bakteriális) Luciferáz (szentjánosbogárból) Alkalikus foszfatáz (humán placenta) szekretált protein! Zöld fluoreszcens fehérje (GFP, medúza)
 β-galaktozidáz (bakteriális) Luciferáz (szentjánosbogárból) Alkalikus foszfatáz (humán placenta) szekretált protein! Zöld fluoreszcens fehérje (GFP, medúza)")
28
phRL-null VEKTOR SZERKEZETE (PROMEGA)
Luciferáz enzim génjét tartalmazza, T7 promótert tartalmaz in vitro transzkripcióra, de nem tartalmaz egyéb promótert vagy szabályozó génszakaszt, biztosan nem tartalmaz kb. 300 ismert konszenzus transzkipciós faktor kötőhelyet ezeket lehet a polilinker helynél a vektorba klónozni

29
AFFINITÁS CÍMKÉVEL (TAG-gal) JELÖLT FEHÉRJE EXPRESSZIÓ
Affinitás címke haszna: 1, fehérje tisztítás megkönnyítése (pl. His-címke) 2, fehérje detektálása (FLAG-címke elleni ellenanyag) 3, fehérje-fehérje interakció vizsgálata (GST fehérjék) 4, jelzett fehérje láthatóvá tétele (zöld fluoreszcens fehérje, GFP) Az címkék lehetnek bakteriális, élesztő vagy emlős expresszióra alkalmasak, de van olyan, mely több élőlényben is jól expresszálódik: pl. GST lehet bakterilis és emlős, vagy a TAP-címke eredetileg élesztőben lett kifejlesztve, de van már TAP emlős expressziós rendszer is.
 2, fehérje detektálása (FLAG-címke elleni ellenanyag) 3, fehérje-fehérje interakció vizsgálata (GST fehérjék) 4, jelzett fehérje láthatóvá tétele (zöld fluoreszcens. fehérje, GFP) Az címkék lehetnek bakteriális, élesztő vagy emlős expresszióra alkalmasak, de van olyan, mely több élőlényben is jól expresszálódik: pl. GST lehet bakterilis és emlős, vagy a TAP-címke eredetileg élesztőben lett. kifejlesztve, de van már TAP emlős expressziós rendszer is.")
30
AFFINITÁS CIMKÉK TÍPUSAI
His-címke: N- vagy C-terminális 6xHisztidin, Nikkel-affinitás gyantához kötődik T7-címke: T7 gén 1 kezdő szekvenciája (11 aminosav) transzláció enhanszer S-címke: ribonukleáz A S-peptid (15 aminosav) detektálás, izolálás: biotinilált S-protein, S-protein affinitás Strap-címke: C-terminális AWRHPQFGG szekvencia (affinitás sztreptavidinhez Epitóp-címkék: jó, általában monoklonális ellenanyag van ellenük 1, FLAG-címke (NYKNNNNK) 2, c-myc-címke (QGKLISGGNL) TAP-címke: „tandem-affinity purification”, kalmodulin-kötő fehérje és protein A egyaránt a vizsgált fehérjéhez van fúzionálva ma az in vivo fehérje-fehérje interakciók egyik leghatásosabb vizsgálati rendszere
 transzláció enhanszer. S-címke: ribonukleáz A S-peptid (15 aminosav) detektálás, izolálás: biotinilált S-protein, S-protein affinitás. Strap-címke: C-terminális AWRHPQFGG szekvencia (affinitás. sztreptavidinhez. Epitóp-címkék: jó, általában monoklonális ellenanyag van ellenük. 1, FLAG-címke (NYKNNNNK) 2, c-myc-címke (QGKLISGGNL) TAP-címke: „tandem-affinity purification , kalmodulin-kötő fehérje és. protein A egyaránt a vizsgált fehérjéhez van fúzionálva. ma az in vivo fehérje-fehérje interakciók egyik leghatásosabb. vizsgálati rendszere.")
31
IN VITRO TRANSZLÁCIÓ Alapvetően az in vitro transzlációs rendszerek két csoportba oszthatók: RNS szolgál mintául vagy DNS Lehet DNS plazmidban vagy RT/PCR termék Lehet teljes sejt-RNS, mRNS vagy in vitro transzkript Tovább lehet osztani az in vitro trasnzkripciós rendszereket aszerint, hogy prokarióta vagy eukarióta gént kerül transzlációra. A fentiek miatt számtalan „kit” került forgalomba, melyek pontos leírását a különböző cégek katalógusaiban (Promega, Novagen, Invitrogen) lehet megtalálni.
 lehet megtalálni.")
32
NÖVEKEDÉSI FAKTOROK JELPÁLYÁJA
tirozin kináz receptor Sos P Grb2 Ras sejtmembrán GTP GDP Grb2 Raf SH SH SH3 MEK P P Erk1,2 P (MAP kinázok) P TCF P SRF SRF c-fos SRE
 P. TCF. P. SRF. SRF. c-fos. SRE.")
33
EGF receptor és Sos exchange fehérje kimutatása

34
FELTÉTELEZTÜK, HOGY A GRB2 KÖTI ÖSSZE AZ EGF
RECEPTOR AZ SOS-SEL, DE NEM VOLT ELLENANYAGUNK A GRB2 ELLEN (MÁSNAK SEM) Arra nincs időnk, hogy anti-grb2 ellenanyagot készítsünk (kb. 3-4 hónap)! EGF receprtor MIT CSINÁLJUNK??? Grb2 P SH3 Ras SH2 SOS GTP GDP SH3
 Arra nincs időnk, hogy anti-grb2 ellenanyagot. készítsünk (kb. 3-4 hónap)! EGF. receprtor. MIT CSINÁLJUNK Grb2. P. SH3. Ras. SH2. SOS. GTP. GDP. SH3.")
35
C-myc CÍMKÉVEL JELZETT GRB2 KÉSZÍTÉSE ÉS STABIL
EXPRESSZIÓJA FIBROBLASZTOKBAN c-myc-Grb2 MYC SH SH SH3 Kalcium-foszfát transzfekcióval jutattuk be a sejtbe a Grb2-DNS-t MYC SH SH SH3 Myc-címke ellen már létezett kiváló mono- klonális ellenanyag fibroblaszt

36
Grb2-SOS IN VIVO KOMPLEXET FORMÁL EGF-KEZELT SEJTEKBEN


 DNS-ből,>")





 prokariótában>")





 Csernetics Árpád Bioinformatika SZIT 2005. ápr. 18.>")






