Az Immunválasz negatív szabályozása
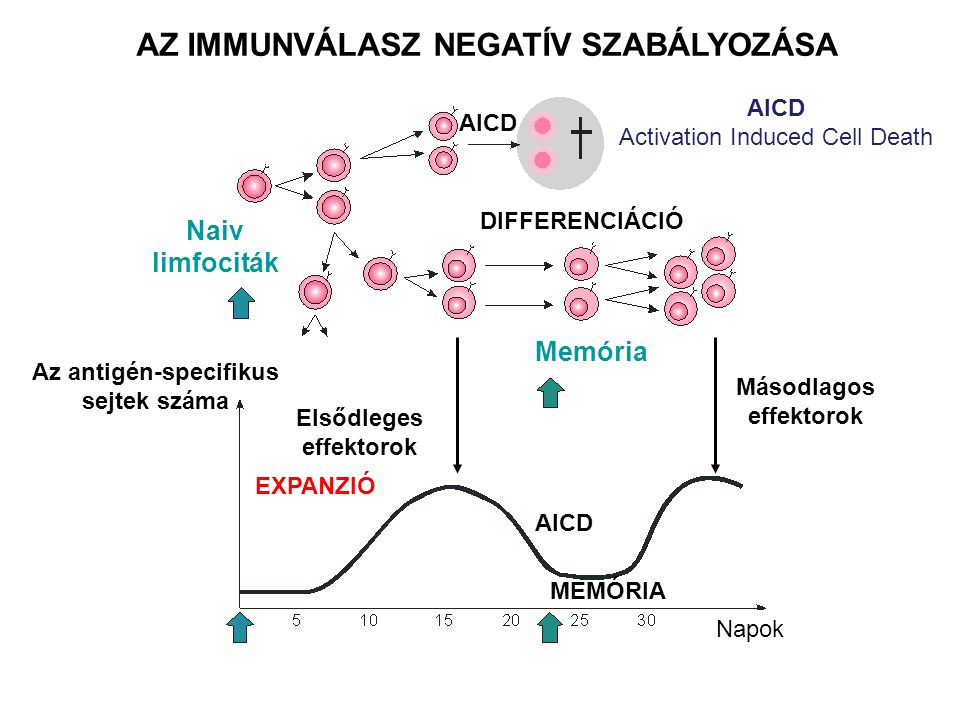
AZ IMMUNVÁLASZ NEGATÍV SZABÁLYOZÁSA AICD Activation Induced Cell Death AICD DIFFERENCIÁCIÓ Naiv limfociták Memória Az antigén-specifikus sejtek száma Másodlagos effektorok Elsődleges effektorok EXPANZIÓ AICD MEMÓRIA Napok
Az effektor T-sejtek eliminációja az immunválasz végén Antigén-indukálta sejthalál (AICD – antigen-induced cell death) A folyamatos T-sejt aktiváció pro-apoptotikus szignálokat indukál (Fas, FasL, Bad, Bax expresszió nő, Bcl-2 expresszió csökken)
Ligandum kötés hatására a TNFR1 és TNFR2 által közvetített pro- és anti-apoptótikus jelpályák
IMMUNOGLOBULIN KÖTŐ Fc RECEPTOROK Ig supergene family, MIRR FcγRI (CD64) FcγRII (CD32) FcεRI FcεRII Iγ-γ α β γ-γ α γ-γ FcγRIII (CD16) FcRI IIIaζ-ζ IIIaζ-γ IIIaγ-γ IIIaβ γ-γ α IgG Fc receptors ITIM – inhibitory motif Ig supergene family, MIRR
A B SEJTEK NEGATÍV SZABÁLYOZÁSA IMMUN KOMPLEXEK ÁLTAL ITAM ITIM FcRIIb
ACTIVATING AND INHIBITORY RECEPTORS IN THE IMMUNE SYSTEM Activating receptor ITAM Expression Inhibitory receptor ITIM Fc receptorok Ig supergene family BCR* B cell TCR,* CD3* T cell FcRIa (CD64) FcRI* Macrophage, DC (internalization) FcRIIa (CD32) FcRIIb (CD32) FcRIIIa (CD16) FcRI* vagy * NK cell (ADCC) FcRI, FcRI* Mast cell (ADCC) C-type lectin FcRIIb (CD23b) FcRIIa (CD23a) MHC I receptorok Human KAR Human KIR NK and T cell C-típusú lektin Egér KIR Human NKG2C/D NK and regulatory T cell Human NKG2A/B Other receptors BCR CD22 CD28 CTLA-4 MIRR Multisubunit Immune Recognition Receptors The ligand binding and signal transducing subunits are separated, they co-localize in membrane microdomains.
A T-sejt válasz szabályozása gátló hatású ko-receptorok által
A CD4+ T SEJTEK SZEREPE AZ APOPTÓZIS KIVÁLTÁSÁBAN T SEJT HOMEOSZTÁZIS AZ IMMUNVÁLASZ LEÁLLÍTÁSA
A kostimuláló molekulák gátló receptorokhoz is kapcsolódhatnak CD28 T sejt CD28 Aktivált T sejt A CTLA-4 - B7 kapcsolat gátolja a T sejt aktivációt - 2 Signal 1 + /CTLA-4 B7 B7 CD28 - B7 kapcsolat A kostimuláció kiváltja a CTLA-4 kifejeződését A CTLA-4 nagyobb affinitással kötődik a B7 molekulához, mint a CD28 A 2. signál hiánya és az ITIM-közvetített jel gátolja a T-sejt választ
A T SEJT AKTIVÁCIÓ NEGATÍV SZABÁLYOZÁSA A B7 MOLEKULÁHOZ KÖTŐDŐ CTLA-4 ÁLTAL APC CD28 aktiváció CTLA-4 ITIM B71/2 KÉSŐI KIFEJEZŐDÉS A MEMBRÁNBAN NAGYOBB AFFINITÁS A B7 MOLEKULÁHOZ MINT A CD28
A CTLA-4 fontos gátló receptor amelyen keresztül a Treg sejtek szabályozzak a hivatásos antigén prezentáló sejteket A CTLA-4 a Treg sejteken direkt és indirekt módon is gátolja a CD80 és CD86 közvetítette szignálokat és gátló jeleket vált ki, amelyek gátolják a gyulladási citokinek termelését valamint aktiválják a gátló hatású indoleamine-2,3-dioxygenase (IDO) termelést A CTLA-4 blokkolása egerekben szerv-specifikus autoimmun betegségeket és bélgyulladást (IBD) indukál
Reguláló T-sejtek
A T LIMFOCITÁK ELSŐDLEGES AKTIVÁLÁSA TÖBBSZINTŰ ELLENŐRZÉS ALATT ÁLL 1. Jel pMHC - TCR 3. Jel Citokinek 2. Jel Kostimuláció Hivatásos APC A DENDRITIKUS SEJTEK A T SEJT VÁLASZ ELSŐDLEGES SZABÁLYOZÓI
A REGULÁLÓ T-LIMFOCITÁK ÉS DENDRITIKUS SEJTEK KAPCSOLATA THYMUS Természetes – nTreg PERIFÉRIA Indukált – iTreg REGULÁLÓ T SEJTEK Treg Kiváltott szabályozás Egyensúlyi szabályozás DC AKTIVÁCIÓ INDUKCIÓ Autoimmun betegségek Transzplantációs tolerancia Malignus kórképek
DENDRITIKUS SEJT REGULÁLÓ T SEJT A REGULÁLÓ T-LIMFOCITÁK DENDRITIKUS SEJTEKTŐL FÜGGŐ KÉPZŐDÉSE DC hiányában nincs immunválasz DC hiánya komplex myelo-/lymphoproliferative szindrómához vezet DENDRITIKUS SEJT Egyensúlyi szabályozás –aktivációs állapot függvénye Szupresszív aktivitás Treg instrukció T effector instrukció DC moduláció Treg – DC szám korrelál DC szám Treg MHC II függő Treg osztódás REGULÁLÓ T SEJT Egyensúlyi szabályozás - állandó osztódás IL-2 függő MHC II függő, saját antigének túlélés ko-stimulációt igényel Treg moduláció Treg DC Treg DC Flt3L függő DC expanzió IFNγ IL-17 A REGULÁLÓ T SEJTEK KÉPZŐDÉSÉT ÉS AKTIVÁCIÓJÁT A DENDRITIKUS SEJTEK IRÁNYÍTJÁK Darrasse-Jéze G et al. JEM 2009
HATÁSMECHANIZMUS A REGULÁLÓ T SEJTEK FUNKCIÓI Perifériás tolerancia fenntartása Autoimmunitás megelőzése Gyulladási folyamatok korlátozása (asthma, gyulladásos bélbetegségek) A fertőző betegségek elleni védelem gátlása A tumor ellenes immunitás korlátozása HATÁSMECHANIZMUS Belső és külső szabályozás Különböző gátló mechanizmusok Sejt kontaktus – Citokinek Kölcsönhatás a célpont effector T sejtekkel
A REGULÁLÓ T SEJTEK EREDETE, TÍPUSAI, FUNKCIÓI PERIFÉRIA FoxP3+ IL-2/TGFβ Fenntartás nTreg mTEC nTreg Effektor T IL-10/IL-35/TGFβ Szupresszió PERIFÉRIA CD4+T FoxP3- iTreg FoxP3+ FoxP3- Szupresszió FoxP3+ Th3 TGFβ IL-10/ TGFβ Effektor T DC FoxP3- Tr1 IL-10/ TGFβ IL-10 Szupresszió TÍMUSZ
A TÍMUSZ EREDETŰ TERMÉSZETES Treg SEJTEK MARKEREI REGULÁLÓ T-SEJTEK A TÍMUSZ EREDETŰ TERMÉSZETES Treg SEJTEK MARKEREI CD4+CD25+FOXP3+ GITR CTLA4 B7 ligand Treg CD25 IL-2Rα FoxP3 CD127 IL-7Rα ↓ Treg differenciálódás, fenntartás, funkció Transzkripciós faktor – sok célgén Önmagában nem elegendő a szupressziv funkcióhoz A TGFβ nem indukál reguláló képességet
A DENDRITIKUS SEJTEK TOLEROGENITÁSA A STIMULÁCIÓS ÁLLAPOT FÜGGVÉNYE TOLEROGÉN DC STIMULÁLT DC IMMUNOGENITÁS DIFFERENCIÁCIÓ TOLEROGENITÁS
A REGULÁLÓ T LIMFOCITÁK KÉPZŐDÉSE KÖRNYEZET FÜGGŐ THYMUS nTreg PERIFÉRIA iTreg Treg NYUGVÓ Foxp3+ Treg Lokális hatások PAMP, TLR, NLR, RLR Gyulladás, IFN, TGFβ T sejt activació AKTIVÁLT Foxp3+ Treg Treg aktiváció CTLA4, GITR, IL-10, IDO PD-1/PD-L Treg át-programozás IL-6, IL-1 Treg Treg lokális tolerancia lokális immunválasz TOLEROGÉN VÁLASZ HELPER VÁLASZ
A REGULÁLÓ T LYMPHOCITÁK SZABÁLYOZÓ FUNKCIÓI Gátló citokinek Citolízis TGFβ IL-10 IL-35 Anyagcsere zavar Dendritikus sejt érés gátlása Indolamin2,3-dioxigenáz LAG-3 – CD4 homológ Treg : effektor T cell = 1 : 8 Treg : DC = 1 : 0,8 Csökkent citokin termelés (IL-2) Sejtkörüli adenozin cAMP transzfer
A REGULÁLÓ T SEJTEK SEJTFELSZÍNI ENZIMJEI EXTRACELLULÁRIS NUKLEOTIDOKAT TERMELNEK Ectonucleoside triphosphate diphosphohydrolase (E-NTPDase Naiv T sejtek toborzása, aktiválása, polarizálása CD4+CD25- effektor sejtek A2A receptort fejeznek ki Sejt körüli/szupresszív Ecto-5’-nucleotidase EBI3 Ebstein-Barr virus induced gene 3 IL-27 és IL-35
AZ AKTIVÁCIÓ INDUKÁLT IL-12 CITOKIN CSALÁD TAGJAI IL-12p70 IFNβ IL-12p35 IFNβ IL-35 Treg EBI3 IL-23p19 IL-12p40 IL-27p28 IL-27 IL-23
AZ IL-35 SZEREPE A REGULÁLÓ T SEJTEK MŰKÖDÉSÉBEN Kezdeti T sejt aktiválás T reg sejtek érzékelik Szupresszív mechanizmusok fokozódnak Az aktivált Treg sejtek molekuláris mintázata effektor sejtek jelenlétében és anélkül más Az EBI3 és IL-12α/p35 (IL-35) alegységek kifejeződése effektor T sejtek jelenlétében nő Az effektor sejtekkel kontaktusban lévő Treg sejtek az IL-35 révén a nem kontaktusban lévő effektor T sejtekre is hatnak Indukált képesség, amiben az effektor sejt is részt vesz NEM CSUPÁN FUNKCIÓ
Dendritikus sejt funkciók gátlásaTreg sejtek által Sakaguchi, Nat Immunol, 2010 Treg sejtek hiányában az effektor T-sejtek adjuvánsként működnek fokozzak a DC-k aktivacióját, növelik az MHC és a ko-stimuláló receptorok expressziójat, valamint fokozzak a gyulladási citokinek termelését
HOGYAN HATNAK A REGULÁLÓ T SEJTEK Treg : effektor T cell = 1 : 8 Treg : DC = 1 : 0,8 Teff Citokin/metabolit közvetített Koncentració/stabilitás DC Treg DC Treg Teff Sejt kontaktustól függő A dendritikus sejtek általi gátlás hatékonyabb, mint az effektor T sejtek szintjén kiváltott hatás
A tumorok környezetében a myeloid sejtek előalakjaiból myeloid suppresszor sejtek fejlődnek Immature myeloid cells (IMCs) are part of the normal process of myelopoiesis, which takes place in the bone marrow and is controlled by a complex network of soluble factors that include cytokines such as granulocyte/macrophage colony-stimulating factor (GM-CSF), stem-cell factor (SCF), interleukin-3 (IL-3), FMS-related tyrosine kinase 3 (FLT-3), macrophage colonystimulating factor (M-CSF) and cell-expressed molecules including Notch (not shown). Haematopoietic stem cells (HSCs) differentiate into common myeloid progenitor (CMP) cells and then into IMCs. Normally, IMCs migrate to different peripheral organs, where they differentiate into dendritic cells, macrophages and/or granulocytes. However, factors produced in the tumour microenvironment and/or during acute or chronic infections, trauma or sepsis, promote the accumulation of IMCs at these sites, prevent their differentiation and induce their activation. These cells exhibit immunosuppressive functions and are therefore known as myeloid-derived suppressor cells (MDSCs). MDSCs can also differentiate into tumorassociated macrophages (TAMs) within the tumour environment, which are cells that have a phenotype and function that is distinct from MDSCs. Gabrilovich and Nagaraj Page 20 Nat
Az immunválasz vagy immuntolerancia kiváltását a regulátor T-sejtek és az effektor sejtek aktivitásának aránya határozza meg A Treg/Teff arány változtatásával az autoimmun folyamatok, szövetkilökődés befolyásolható
Az AICD kiváltásáért felelős jelátviteli utak Figure 15-10 Signaling Pathways of AICD Elements of the extrinsic and intrinsic apoptotic pathways are shown in blue and grey, respectively. Extrinsic pathway: prolonged TCR engagement delivers signals via ZAP-70 and NFAT that increase TNFR1 and Fas expression. Upon the binding of several FasL molecules expressed on an APC to Fas expressed on the activated T cell (only one pair is shown for clarity), the adaptor protein FADD is recruited to the cytoplasmic tail of the Fas molecule. FADD in turn recruits procaspase-8, which cleaves itself to generate active caspase-8. Caspase-8 cleaves procaspase-3 to activate this key executioner enzyme, which in turn cleaves procaspase-6 and procaspase-7. Activated caspase-6 and -7 are responsible for many of the morphological hallmarks of apoptotic cells. Caspase-3 also degrades ICAD, releasing CAD, which promotes DNA degradation. The extrinsic pathway is also triggered by the binding of TNF to TNFR1. The adaptor TRADD recruited to the TNFR1 complex can recruit FADD, which proceeds to activate caspase-8 as described previously for Fas killing. Intrinsic pathway: external apoptotic stimuli (including granzymes) induce the release of cytochrome c from the activated T cell’s mitochondria. In the presence of dATP, cytochrome c associates with Apaf-1 to promote cleavage of procaspase-9. Activated caspase-9 joins Apaf-1 and cytochrome c to form the apoptosome. The apoptosome can then cleave caspase-3, triggering the downstream events of the extrinsic pathway. The intrinsic pathway is promoted when the anti-apoptotic molecule BiD is cleaved by caspase-8. Truncated Bid (tBiD) binds to Bax and Bak in the mitochondrial membrane and induces cytochrome c release.