Előadást letölteni
1
A DNS szerkezete és replikációja

2
Mit kell „tudnia” a genetikai anyagnak?
1. Rendelkeznie kell az információ tárolásának képességével. Tehát kémiailag elegendően stabilnak kell lennie és valamilyen módon képesnek kell lennie adatok tárolására, amelyek kiolvashatók belőle. 2. Képesnek kell lennie ezen információt pontosan megkétszerezni és továbbadni az osztódások során. 3. Képesnek kell lenni az állandóság mellett változásra is. A változás mértéke azonban nem lehet túl nagy (nem lenne öröklődés), de nem lehet nulla, mert akkor nem lenne evolúció. A DNS 1900-as évek elején ismert kémiai szerkezete (4-féle szerves bázis, foszforsav és dezoxiribóz) túlságosan egyszerű felépítésűnek tűnt ahhoz, hogy a fenti feladatoknak megfelelhessen. Sokkal alkalmasabbnak tűntek a fehérjék (20-féle aminosav építi fel).
, de nem lehet nulla, mert akkor nem lenne evolúció. A DNS 1900-as évek elején ismert kémiai szerkezete (4-féle szerves bázis, foszforsav és dezoxiribóz) túlságosan egyszerű felépítésűnek tűnt ahhoz, hogy a fenti feladatoknak megfelelhessen. Sokkal alkalmasabbnak tűntek a fehérjék (20-féle aminosav építi fel).")
3
Az örökítő anyag, - a DNS - szerkezetét és működésmódját
1953-ban írta le James Watson és Francis Crick. Munkájukat a következő előzményekre alapozhatták: Az egyes tulajdonságokat öröklődő részecskék (gének) alakítják ki. (Mendel) - A gének fehérjék szerkezetét befolyásolják,egy gén ~ egy enzim. (Beadle és Tatum). - A gének kromoszómákon vannak (Bridges). A kromoszómák DNS-ből és fehérjékből állnak. A DNS röntgendiffrakciós szerkezetvizsgálata (Wilkins és Franklin). - Az örökítőanyag a DNS.
 alakítják ki. (Mendel) - A gének fehérjék szerkezetét befolyásolják,egy gén ~ egy enzim. (Beadle és Tatum). - A gének kromoszómákon vannak (Bridges). A kromoszómák DNS-ből és fehérjékből állnak. A DNS röntgendiffrakciós szerkezetvizsgálata (Wilkins és Franklin). - Az örökítőanyag a DNS.")
4
A baktérium transzformáció felfedezése
Frederick Griffith kísérlete 1928. R törzs S törzs A Streptococcus pneumoniae (tüdőgyulladást okozó bakt.) virulens, S (tokképző) törzsével beoltott egerek tüdőgyulladásban elpusztulnak, az avirulens, R (tokot nem képző) törzzsel beoltottak túlélnek. Az S törzs baktériumai tokot választanak ki, ami megvédi őket az immunrendszertől.
 virulens, S (tokképző) törzsével beoltott egerek tüdőgyulladásban elpusztulnak, az avirulens, R (tokot nem képző) törzzsel beoltottak túlélnek. Az S törzs baktériumai tokot választanak ki, ami megvédi őket az immunrendszertől.")
5
A baktérium transzformáció felfedezése
Griffith kísérlete 1928. A hővel elölt S baktériumok és az élő nem-virulens R baktériumok keverékével beoltott egerek elpusztulnak. A hővel kezelt S törzs nem pusztítja el az egereket. Az utolsó kísérlet döglött egereiből élő S baktériumok tenyészthetők ki. Az elölt baktériumok anyagából valami az R baktériumokat S-é alakította át (transzformálta).
.")
6
Az S törzs elölt baktériumaiból kiszabadult anyagot felvették az R törzs sejtjei és ezáltal megváltozott a természetük: maguk is tok termelésre váltak képessé, átalakultak, transzformálódtak. Kérdés, hogy mi lehet az az anyag, amely a transzformációt okozza?

7
A genetikai anyag a DNS Sokáig úgy gondolták, hogy a transzformáló anyag a fehérje, csak 1944-ben igazolták először, hogy a genetikai anyag a DNS. O. Avery (és C.M. Mac Leod és M. McCarty) kísérlete, 1944. A DNS a transzformáló anyag. Az S sejtekből kivont anyagokból egyedül a DNS az, amivel az R sejtek S formává alakíthatók. A transzformáló anyagot nem lehetett elbontani proteázokkal, szénhidrát és zsírontó enzimekkel, de hatástalanná vált nukleázok alkalmazása esetén. Csak a tisztított DNS képes a baktériumokat transzformálni. A DNS transzformáló képessége igazolta először, hogy a gének DNS-ből állnak.
 kísérlete, A DNS a transzformáló anyag. Az S sejtekből kivont anyagokból egyedül a DNS az, amivel az R sejtek S formává alakíthatók. A transzformáló anyagot nem lehetett elbontani proteázokkal, szénhidrát és zsírontó enzimekkel, de hatástalanná vált nukleázok alkalmazása esetén. Csak a tisztított DNS képes a baktériumokat transzformálni. A DNS transzformáló képessége igazolta először, hogy a gének DNS-ből állnak.")
8
A Hershey-Chase kísérlet (1952)
A radioaktivitás a leváló üres fág fejekben észlelhető. jelölt fehérje elválasztás A radioaktivitás a baktériumokban észlelhető, majd a következő fág generációban is megjelenik. jelölt DNS elválasztás A Hershey-Chase kísérlet igazolta, hogy a fágok örökítő anyaga a DNS nem pedig a fehérje. A kísérlet kétféleképpen előkészített T2 fágot használt. Az egyik esetén a fehérje burkot radioaktív kénnel (35S) jelölték, mert a kén nem fordul elő a DNS-ben. A másik esetben radioaktív foszforral (32P) a DNS-t jelölték, mert a foszfor nem fordul elő a fehérjében. Csak a 35P injektálódott az baktériumba, jelezve, hogy a DNS az a szükséges anyag, ami az új fágok létrejöttéhez szükséges.
 jelölték, mert a kén nem fordul elő a DNS-ben. A másik esetben radioaktív foszforral (32P) a DNS-t jelölték, mert a foszfor nem fordul elő a fehérjében. Csak a 35P injektálódott az baktériumba, jelezve, hogy a DNS az a szükséges anyag, ami az új fágok létrejöttéhez szükséges.")
9
A DNS kémiai összetevői
A DNS kémiai felépítésének alapegysége a nukleotid (nukleozid-monofoszfát). A DNS ezek polimerje – polinukleotid (hasonlóan az RNS-hez). A nukleotid foszforsavat (foszfátot), de(z)oxiribóz cukrot és négyféle N tartalmú heterociklusos szerves bázisból egyet tartalmaz. A négy bázis az adenin (A), a guanin (G), a citozin (C) és a timin (T). Ezek közül az A és G purinvázas, nagy méretű, a C és T pirimidinvázas, kis méretű bázisok. A nukleotidok teljes kémiai neve: rövidítése dezoxiadenozin-5’-monofoszfát, dAMP A dezoxiguanozin-5’-monofoszfát, dGMP G dezoxicitidin-5’-monofoszfát, dCMP C dezoxitimidin-5’-monofoszfát, dTMP T
. A DNS ezek polimerje – polinukleotid (hasonlóan az RNS-hez). A nukleotid foszforsavat (foszfátot), de(z)oxiribóz cukrot és négyféle N tartalmú heterociklusos szerves bázisból egyet tartalmaz. A négy bázis az adenin (A), a guanin (G), a citozin (C) és a timin (T). Ezek közül az A és G purinvázas, nagy méretű, a C és T pirimidinvázas, kis méretű bázisok. A nukleotidok teljes kémiai neve: rövidítése. dezoxiadenozin-5’-monofoszfát, dAMP - A. dezoxiguanozin-5’-monofoszfát, dGMP - G. dezoxicitidin-5’-monofoszfát, dCMP - C. dezoxitimidin-5’-monofoszfát, dTMP - T.")
10
A nukleinsav bázisok és cukrok
nagy, purin bázisok kicsi, pirimidin bázisok A nukleinsav bázisok és cukrok A nukleinsavak viszonylag egyszerű felépítésű makromolekulák, amelyekben öt féle szerves bázis fordul elő. A DNS-ben lévő cukor egy aldopentóz, a dezoxiribóz C5H10O4 (az RNS-ben a ribóz C5H10O5). A DNS-ben lévő cukornak nincs szabad hidroxil csoportja, ami növeli a DNS kémiai stabilitását. A bázisok közül négy csak a DNS-ben, négy csak az RNS-ben fordul elő. DNS: ATCG RNS: AUCG A szeves bázisok oxo-enol átalakuláson mehetnek keresztül és hidrogén-hid kötések kialakítására képes csoportjaik vannak.
. A DNS-ben lévő cukornak nincs szabad hidroxil csoportja, ami növeli a DNS kémiai stabilitását. A bázisok közül négy csak a DNS-ben, négy csak az RNS-ben fordul elő. DNS: ATCG. RNS: AUCG. A szeves bázisok oxo-enol átalakuláson mehetnek keresztül és hidrogén-hid kötések kialakítására képes csoportjaik vannak.")
11
A DNS kémiai összetevői

12
Nukleotid szerkezete 3’ C atom – ezzel kapcsolódik a polinukleotid lánc következő nukleotidjának foszforsav részéhez N tartalmú, heterociklusos szerves bázis (adenin) Foszforsav rész (foszfát csoport) Cukor – aldopentóz (ribóz) 1’ C atom (glikozidos OH csop.) – bázis kapcsolódási helye 5’ C atom – a foszforsav rész kapcsolódási helye 2’ C atom – ezen nincs O a DNS-ben
 Foszforsav rész (foszfát csoport) Cukor – aldopentóz (ribóz) 1’ C atom (glikozidos OH csop.) – bázis kapcsolódási helye. 5’ C atom – a foszforsav rész kapcsolódási helye. 2’ C atom – ezen nincs O a DNS-ben.")
13
A Chargaff szabályok Kölönböző élőlényekből kivonható DNS összetételének vizsgálata érdekes törvényszerűségeket tárt fel. A törvényszerűségeket Erwin Chargaff ismerte fel: Az élőlényekből származó DNS-ekben a pirimidin nukleotidok (T + C) mennyisége egyenlő a purin (A + G) nukleotidok mennyiségével. A T mennyisége egyenlő az A-val, és C mennyisége egyenlő G-vel. Az A + T és C + G mennyiségek nem egyenlők, azok aránya jellemző az élőlényre amiből a DNS származik.
 mennyisége egyenlő a purin (A + G) nukleotidok mennyiségével. A T mennyisége egyenlő az A-val, és C mennyisége egyenlő G-vel. Az A + T és C + G mennyiségek nem egyenlők, azok aránya jellemző az élőlényre amiből a DNS származik.")
14
A DNS röntgen diffrakciós képe (R.Franklin és M.Wilkins 1953.)
")
15
A DNS röntgen diffrakciós képe
(R.Franklin és M.Wilkins 1953) A röntgen diffrakcióval kapott adatok azt jelezték, hogy - a molekula fonálszerű, - a fonál két párhuzamos szerkezetből áll. - egyenletes átmérőjű, - spirál alakú.
 A röntgen diffrakcióval kapott adatok azt jelezték, hogy. - a molekula fonálszerű, - a fonál két párhuzamos szerkezetből áll. - egyenletes átmérőjű, - spirál alakú.")
16
A DNS térszerkezetét Watson és Crick oldotta meg
l953-ban. A modell kidolgozása során összeillesztették a röntgen diffrakciós adatokat, a Chragaff szabályokat és a DNS és alkotórészeiről felhalmozódott kémiai ismereteket olymódon, hogy a modell eleget tehessen az örökítő anyag által támasztott követelményeknek.

17
H-hidakkal kapcsolódó komplementer bázispárok
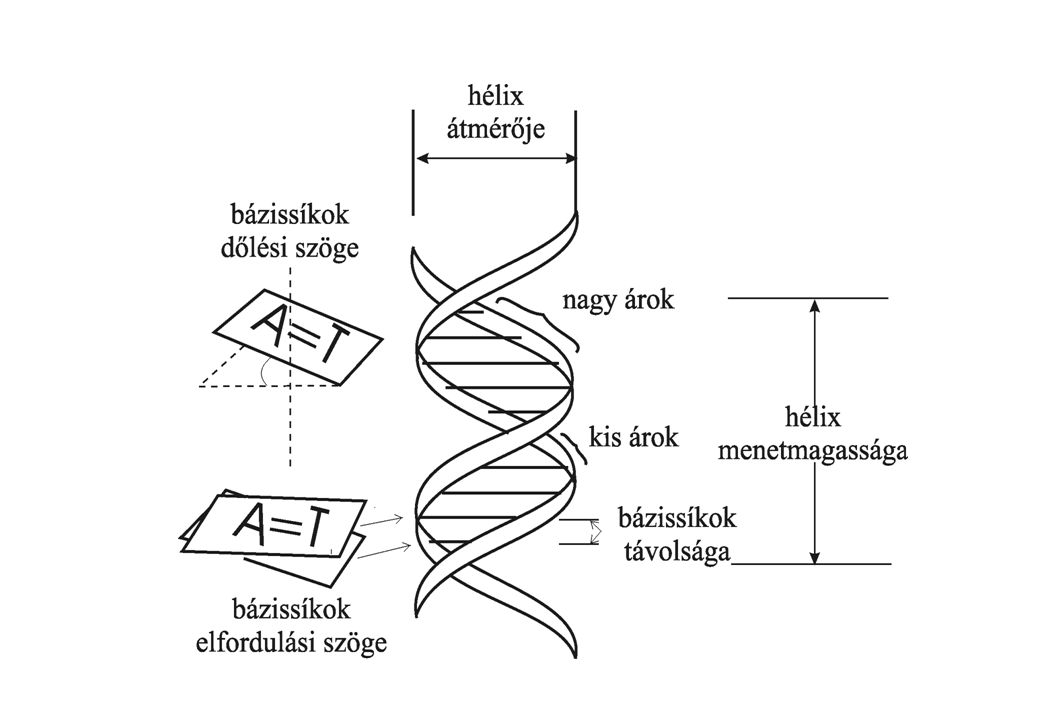
A kettős spirál szalag modellje A kettős spirál egyszerűsített képe. A pálcák a bázispárokat képviselik. A szalagok a két antiparallel lánc cukorfoszfát gerincét képviselik. A méretek angström-ben (1Å = 0,1 nm) mutatják a távolságokat. A spirál 10 bázisonként fordul csaknem pontosan 360o-ot. Cukor-foszfát lánc H-hidakkal kapcsolódó komplementer bázispárok
 mutatják a távolságokat. A spirál 10 bázisonként fordul csaknem pontosan 360o-ot. Cukor-foszfát lánc. H-hidakkal kapcsolódó komplementer bázispárok.")
18
A kettős spirál jellemzői
A nukleotidok szabályosan ismétlődő távolságokban egymás felett helyezkednek el. A nukleotidok lapos molekuláinak síkja merőleges a szál hossztengelyére. A DNS tér-modell két ellentétes lefutású, u.n. antiparallel szálból épül. A modell egyenletes átmérője a Chargaff szabályok követésével biztosítható úgy, hogy purin bázissal pirimidin bázis áll szemben. Ezeket egymáshoz hidrogén hidak rögzítik.

20
A kettős spirál kémiai szerkezete
A modellben a vízben kevéssé oldódó (kevésbé poláros, hidrofób) bázisok belül, a vízben jól oldódó (erősen poláros és ionos, hidrofil) cukor és foszfát csoportok kívül helyezkednek el. A molekula gerincét alkotó láncban a foszforsav észter kötéssel (csoporttal) kapcsolódik a cukrok 3’ és 5’ szénatomjához. Minden bázispár egy purin (A vagy G) és egy pirimidin bázist (T vagy C) tartalmaz. Az A-T párt 2, a G-C párt 3 hidrogénhíd kapcsolja össze. A két lánc egymás komplementere. Az antiparallel irányultságot a cukor-foszfát lánc ellentétes 5’-3’ iránya adja. A cukor foszfát lánc igen rugalmas, a hajlítást, csavarást igen jól tűri. 5’ 3’ 3’ 5’
 bázisok belül, a vízben jól oldódó (erősen poláros és ionos, hidrofil) cukor és foszfát csoportok kívül helyezkednek el. A molekula gerincét alkotó láncban a foszforsav észter kötéssel (csoporttal) kapcsolódik a cukrok 3’ és 5’ szénatomjához. Minden bázispár egy purin (A vagy G) és egy pirimidin bázist (T vagy C) tartalmaz. Az A-T párt 2, a G-C párt 3 hidrogénhíd kapcsolja össze. A két lánc egymás komplementere. Az antiparallel irányultságot a cukor-foszfát lánc ellentétes 5’-3’ iránya adja. A cukor foszfát lánc igen rugalmas, a hajlítást, csavarást igen jól tűri. 5’ 3’ 3’ 5’")
21
Lehetséges bázis párosodás a kettős spirálban
Csak a purin - pirimidin párosodás felel meg a DNS szál röntgennel meghatározott átmérőjének. Ugyancsak ez a kombináció felel meg Chargaff első szabályának.

22
A bázis párosodás módja
A négy lehetséges purin-pirimidin bázispárból (A-T, A-C, G-C, G-T) csak kettő, az A-T és a G-C felel meg Chargaff második szabályának. Watson és Crick kimutatta, hogy csak az A-T és G-C bázispárok képesek hidrogén hidakkal a modellbe beillő módon összekapcsolódni. A modell azt jósolja, hogy a nagy G-C tartalmú DNS stabilabb a nagy A-T tartalmúnál. Ez a jóslat beigazolódott.
 csak kettő, az A-T és a G-C felel meg Chargaff második szabályának. Watson és Crick kimutatta, hogy csak az A-T és G-C bázispárok képesek hidrogén hidakkal a modellbe beillő módon összekapcsolódni. A modell azt jósolja, hogy a nagy G-C tartalmú DNS stabilabb a nagy A-T. tartalmúnál. Ez a jóslat beigazolódott.")
23
A DNS reverzibilis denaturációja
A bázisok közötti kölcsönhatás erőssége függ a környezet pH-jától, az ionkoncentrációjától és a hőmérséklettől. Magas hőmérsékleten (vagy nagyon alacsony ionkoncentráció mellett) a DNS-t összetartó erők nem elégségesek a két lánc összetartására, a DNS láncai széttekerednek, a DNS denaturálódik A denaturáció reverzibilis folyamat (a láncok összeállását molekuláris hibridizációnak nevezzük)
 a DNS-t összetartó erők nem elégségesek a két lánc összetartására, a DNS láncai széttekerednek, a DNS denaturálódik. A denaturáció reverzibilis folyamat (a láncok összeállását molekuláris hibridizációnak nevezzük)")
24
A DNS többféle formát vehet fel
Az élőlényekben és vizes oldatban a „B” forma a leggyakoribb, ebben a bázisok síkja majdnem merőleges a cukor-foszfát gerincre. Dehidrált körülmények között egy tömörebb „A” forma jön létre, melyben a bázisok síkja megdől. Hosszú GCGCGC.... ismétlődések a Z formát vehetik fel, amely balmenetes, zegzugos lefutású és megnyúlt. A B Z

25
A B forma létra modellje
A „B” forma részletes térszerkezetén jól látszik, hogy a bázispárok létrafokként helyezkednek el a szerkezet belsejében. A cukor gyűrű síkja majdnem merőleges a bázisok síkjára.

26
A DNS megkettőződés (replikáció) jóslata
A DNS kettős spirál szerkezetéből közvetlenül adódik a megkettőződés mikéntje. A bázis párosodás szigorú törvényéből az következik, hogy amennyiben a kettős spirál két szála a H-kötések mentén kettéválik, mindkét szál mintaként (templátként) szolgálhat egy új szál szintéziséhez, melynek során az eredeti szállal és egymással megegyező szerkezetek jönnek létre. Ezzel magyarázatot nyer a mitózis jelensége, az örökítőanyag pontos átadása. A genetikailag kódolt információt a nukleotidok sorrend adja.
 szolgálhat egy új szál szintéziséhez, melynek során az eredeti szállal és egymással megegyező szerkezetek jönnek létre. Ezzel magyarázatot nyer a mitózis jelensége, az örökítőanyag pontos átadása. A genetikailag kódolt információt a nukleotidok sorrend adja.")
27
A DNS elméletileg lehetséges replikációs módjai
szemikonzervatív konzervatív* diszpezív* A régi szál sötét színű, az újonnan szintetizált világos. *nem tananyag

28
A Meselson-Stahl kísérlet (1958)*
A több generáción keresztül 15N táptalajon tartott baktériumokból származó DNS nehéz sávot ad centrifugálással. A normál (14N) táptalajon nevelt baktériumok DNS-e pedig könnyű sávot. Ha a 15N-en tartott sejteket átteszik könnyű táptalajra, az első nemzedékben köztes, a második után könnyű és köztes sáv figyelhető meg. *nem tananyag
 táptalajon nevelt baktériumok DNS-e pedig könnyű sávot. Ha a 15N-en tartott sejteket átteszik könnyű táptalajra, az első nemzedékben köztes, a második után könnyű és köztes sáv figyelhető meg. *nem tananyag.")
29
A DNS replikáció mechanizmusa A replikációs villa

30
A DNS szintézis kezdése (priming)
A DNS szintézist egy rövid RNS primer szintézise előzi meg, melyet az RNS polimeráz (primáz) készít. A primáz egy fehérje komplex része, amelyet primoszómának neveznek. A szintézis iránya az egyik szálon a villa felé mutat, a másik szálon a villától távolodik. primáz RNS primer szintézis irány RNS primer
 készít. A primáz egy fehérje komplex része, amelyet primoszómának neveznek. A szintézis iránya az egyik. szálon a villa felé mutat, a másik szálon a villától. távolodik. primáz. RNS. primer. szintézis. irány. RNS primer.")
31
A DNS polimerázok működése
templát + primer + (minta) (indító) dATP + dGTP dCTP dTTP DNS polimeráz új DNS szál A DNS polimerázok az egyes szálú DNS templátra azt kiegészítő (komplementer) szálat szintetizálnak a rendelkezésre álló nukleotid trifoszfátokból. A szálat azonban elkezdeni nem tudják, csak hosszabbítani. A kezdéshez egy rövid kezdő (templát) szakaszra van szükségük.
 (indító) dATP. + dGTP. dCTP. dTTP. DNS polimeráz. új DNS. szál. A DNS polimerázok az egyes szálú DNS templátra azt kiegészítő (komplementer) szálat szintetizálnak a rendelkezésre álló nukleotid trifoszfátokból. A szálat azonban elkezdeni nem tudják, csak hosszabbítani. A kezdéshez egy rövid kezdő (templát) szakaszra van szükségük.")
32
A DNS polimeráz lánc hosszabbítása
5’ szén atom 3’ szén atom t e m p l a s r n d primer strand A DNS polimeráz lánc hosszabbítása A DNS polimeráz az új nukleotid a foszfátját a lánc végén lévő cukor 3’ OH csoportjához kapcsolja. Ezzel az új lánc ’ 3’ irányban hosszabbodik. a lánc hosszabodás iránya

33
Vezető szál, elmaradó szál
régi szál vezető szál elmaradó szál a villa mozgása Az elmaradó szál szintézise: RNS primer új DNS Okazaki fragment ligálás 1., 2., 3., 4., A vezető szál szintézise folytonos. Az elmaradó szálon: 1., A primáz RNS-t szintetizál. 2., A DNS polimeráz III DNS-t szintetizál a primer folytatásaként. 3., A DNS polimeráz I eltávolítja az előtte lévő RNS darabot és befejezi a láncot. 4., A DNS ligáz összekapcsolja a különálló DNS darabokat.

34
A DNS replikáció enzimei
topoizomeráz A topoizomeráz a szál kitekerése hatására a villa túloldalán felhalmozódó torziós feszültséget (csavarodást) csökkenti olymódon, hogy az egyik szálat elvágja, azt a másik szál körül „kipörgeti”, majd a vágást vissza-ligálja.
 csökkenti olymódon, hogy az egyik szálat elvágja, azt a másik szál körül „kipörgeti , majd a vágást vissza-ligálja.")
35
mozi


 DNS-ből,>")











>")






