Előadást letölteni
2
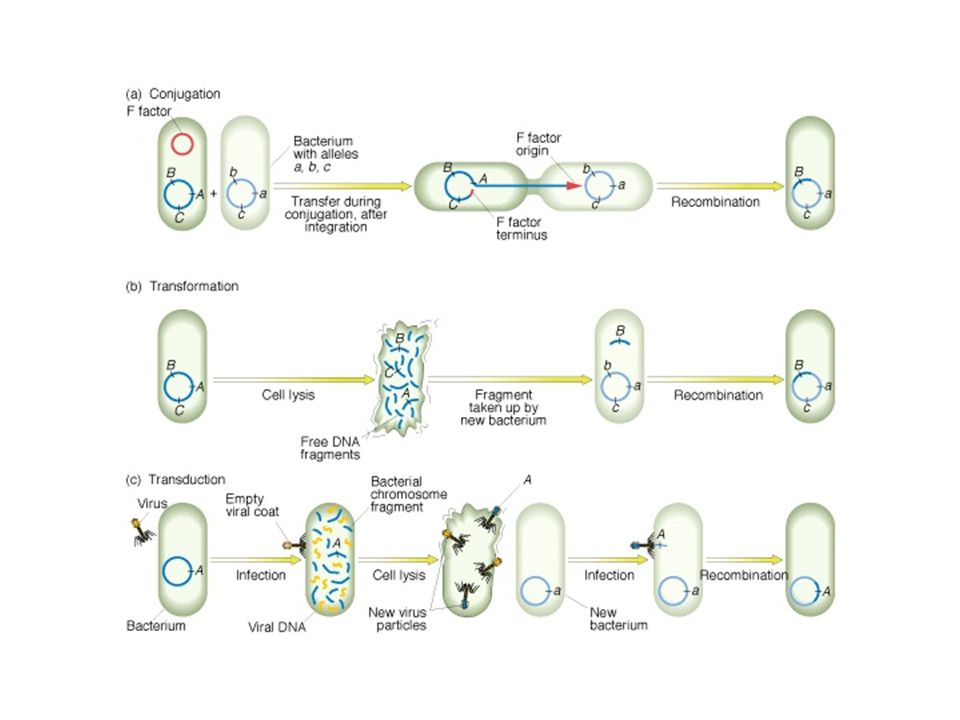
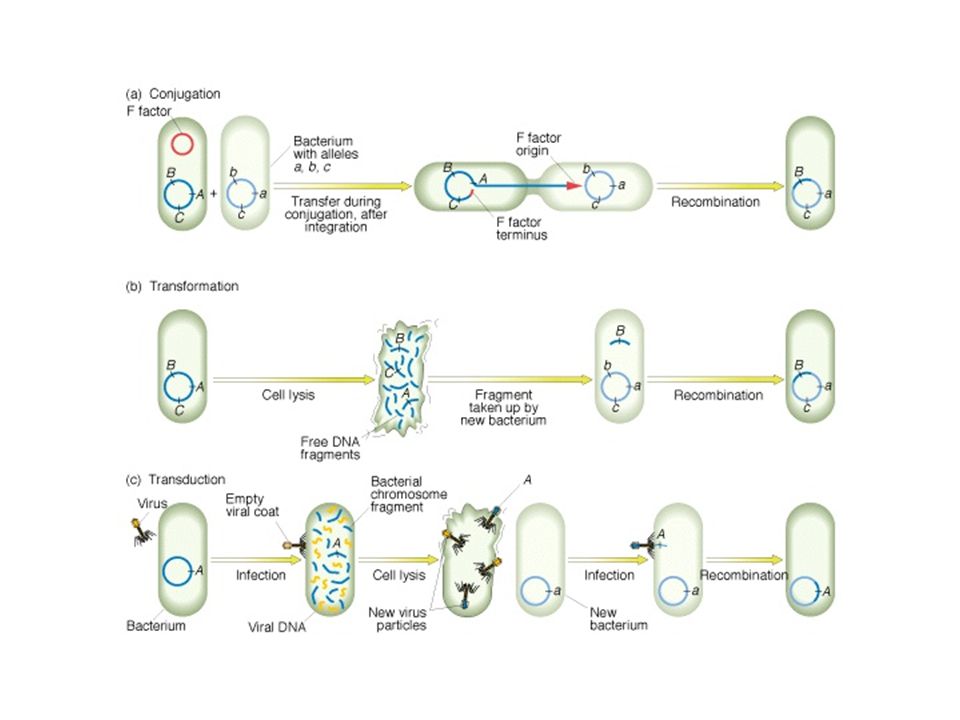
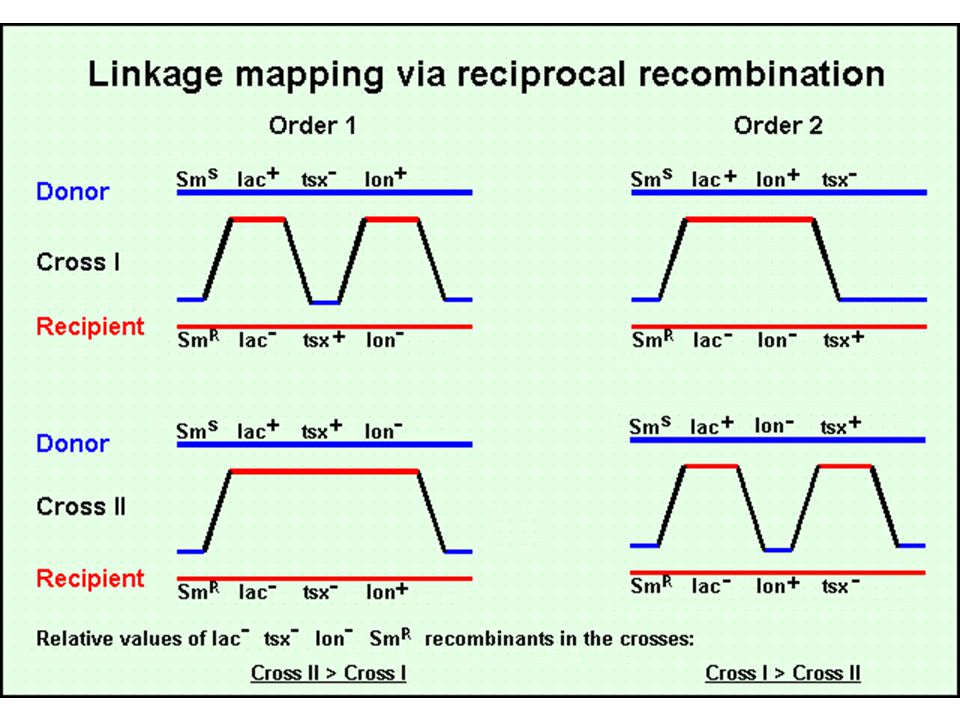
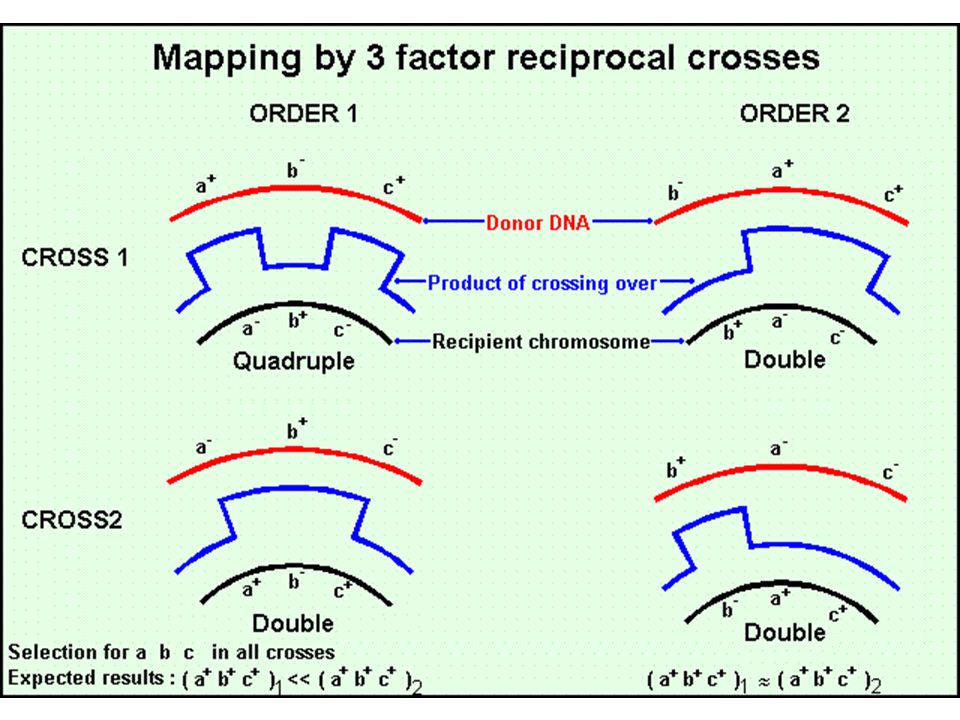
Transzdukció Bakteriofágok közvetítte genetikai információ csere
Két fő bakteriofág típus Virulens fágok/lítikus fágok – fertőzik a sejtet, gyorsan szaporodnak, lizálják a sejtet Mérsékelt fágok/temperált fágok – fertőzik a sejtet, be tudnak épülni a genomba és profágként alvó állapotban – néha kivágódnak, szaporodnak és lizálják a sejtet

3
Bakteriofágok morfológiája változatos
RNS-t vagy DNS-t tartalmaznak fehérje burokban Szinte minden baktériumnak van fágja.

4
Nukleinsavukat a gazda sejtbe injektálják
A gazdabaktériumuk sejtjének felületén lévő specifikus receptorhoz kapcsolódnak Nukleinsavukat a gazda sejtbe injektálják Smithsonian (Oct 2000) T4 bakteriofágok egy E. coli sejt felületén
 T4 bakteriofágok egy E. coli sejt felületén.")
5
Mérsékelt fág A gazda genomba épül be – félalvó állapot (a baktérium így lizogén bakt.) (pl. λ bakteriofág)
 .")
6
Fágkonverzió Az alvó profág – az integrálódott bakteriofág – a mikróba fenotípusát megváltoztató géneket hordoz - pl. a patogének és a toxin termelés toxin profág inzerciós hely (att site) Corynebacterium diptheriaea A fág génekről termelődik a toxin. Ez betegíti meg az embert. C. diptheriaea A fág nélkül a törzs nem termel toxint. Nem okoz diftériát.
 Corynebacterium diptheriaea. A fág génekről termelődik. a toxin. Ez betegíti meg az embert. C. diptheriaea. A fág nélkül a törzs nem termel toxint. Nem okoz diftériát.")
7
A fágfertőzés lizogén útvonala

8
Transzdukció (specializált)
specializált transzdukció LFT, kis frekvenciájú transzdukáló lizátum HFT, nagy frekvenciájú transzdukáló lizátum A specializált transzdukció fág

9
A specializált transzdukció egyszerű mechanizmusa

10
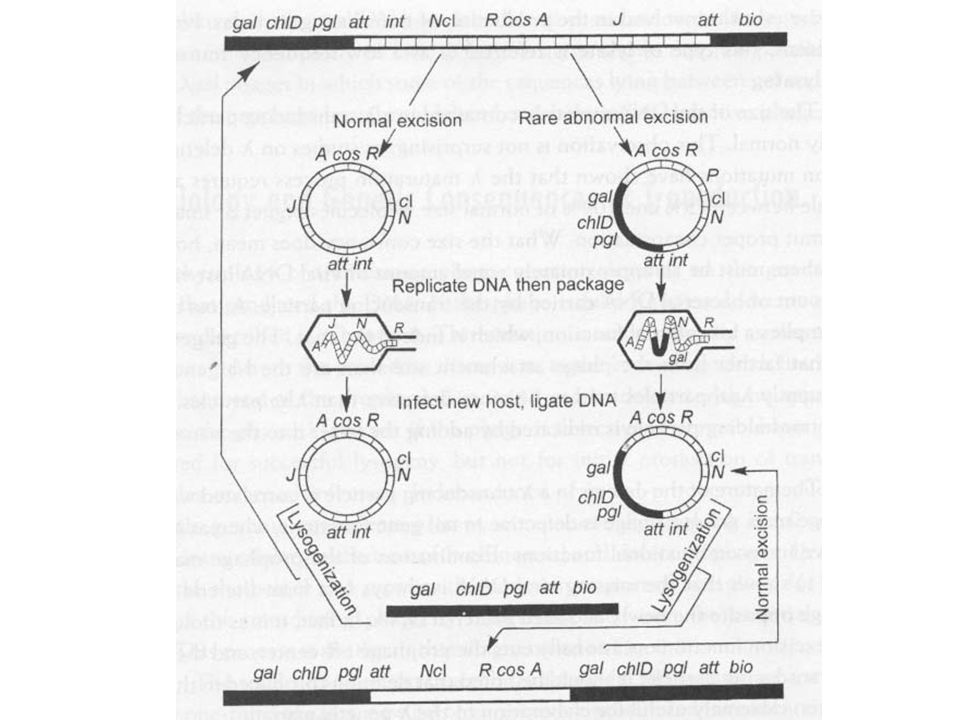
Specializált transzdukció (Hibás kivágódás!!!)
")
11
Specializált transzdukció
fágnál (gal, bio markerek) A profág pontatlan kivágódásával A profág beépülése melletti gének is bejutnak a fágfejbe Kis gyakoriságú jelenség A hiba folytán kis valószínűséggel keletkeznek olyan fágok is, melyek hordoznak baktérium gént LFT, kis gyakoriságú transzdukáló lizátum Mivel a lambda fág fejmérete nagyobb, mint ami a fág DNS-hez szükséges méret (75%-109%) transzdukció esetén fág funkció sérülhet (defektív fág, minél nagyobb az átvitt bakterium DNS darab annál defektívebb a fág)
 A profág pontatlan kivágódásával. A profág beépülése melletti gének is bejutnak a fágfejbe. Kis gyakoriságú jelenség. A hiba folytán kis valószínűséggel keletkeznek olyan fágok is, melyek hordoznak baktérium gént. LFT, kis gyakoriságú transzdukáló lizátum. Mivel a lambda fág fejmérete nagyobb, mint ami a fág DNS-hez szükséges méret (75%-109%) transzdukció esetén fág funkció sérülhet (defektív fág, minél nagyobb az átvitt bakterium DNS darab annál defektívebb a fág)")
13
Ha a fág defektív, helper fág kellhet a sikeres lizogénia kialakításához
A termelésnél nincs probléma, mert a jelenlévő intakt fág funkciók biztosítják a normális működést (többszörös genom ekvivalens) Transzdukáló fággal lizogenizált baktérium is előfordul Ugyanúgy indukálható, mint más lizogén Ha defektív, akkor nem keletkezik fágrészecske Ha a defektív funkciót teljes helper fággal kiegészítjük, indukció során az összes fágrészecske 50%-a transzdukáló HFT High frequency
 Transzdukáló fággal lizogenizált baktérium is előfordul. Ugyanúgy indukálható, mint más lizogén. Ha defektív, akkor nem keletkezik fágrészecske. Ha a defektív funkciót teljes helper fággal kiegészítjük, indukció során az összes fágrészecske 50%-a transzdukáló HFT High frequency.")
14
A virulens fágok egylépéses szaporodási görbéje
- A latencia periódus alatt a fággenom replikálódik és a gazda genom teljesen leépül

15
A fágfertőzés látható hatása a gazda DNS-re
T4 fág DNS Fertőzés előtt Fertőzés után

16
A fágfertőzés lítikus ciklusa

17
Általános transzdukció (hibás pakolás!!!)
")
18
Transzdukció DNS átvitel baktérium sejtek között bakteriofág segítségével Általános transzdukció A transzdukáló fágrészecske csak bakteriális DNS-t hordoz Pszeudovirion, mert a saját szaporodásához szükséges tulajdonságokat nem hordozza

19
A fágok egy részénél fejméretű a pakolás, ami lehet hibás is – a pakoló mechanizmusnak nincs specifikus felismerő helye Így pakol a Salmonella typhimurium P22 fágja és B. subtilis PBS1fágja

20
Mi történik a recipiens sejteben?

21
A baktériumok közötti géncsere hatékony módja a generalizált, vagy általános transzdukció
Hibás pakolás!!! Közeli genetikai bélyegek térképezése!

22
Példa – transzdukió a Salmonella typhimurium P22 fágjával
P22 HT – hatékony általános transzdukáló - hanyag fág – a fágrészecskék 50%-ában gazda DNS, azaz transzdukáló részecske ( transducing particles = TP) Minden fejben (TP) 44 kb-nyi DNS – a Salmonella genom kb kb nagyságú Ezért, ha a pakolás teljesen véletlenszerű, akkor 100 különböző fágrészecske a teljes genomot reprezentálhatja. 1011 P22 HT fág/ml szaporítható – tehát ebből 5 x 1010 –ben gazda DNS van 1 ml-ben kb. 5 x 108 Salmonella genomnyi (0.5)(1011 fág/ml)/(100 TP [1 genom]) = 108 genom kópia/ml a lizátumban
 Minden fejben (TP) 44 kb-nyi DNS – a Salmonella genom kb kb nagyságú. Ezért, ha a pakolás teljesen véletlenszerű, akkor 100 különböző fágrészecske a teljes genomot reprezentálhatja P22 HT fág/ml szaporítható – tehát ebből 5 x 1010 –ben gazda DNS van 1 ml-ben kb. 5 x 108 Salmonella genomnyi. (0.5)(1011 fág/ml)/(100 TP [1 genom]) = 108 genom kópia/ml a lizátumban.")
23
P1 fág 100 kbp Pac helytől fejméretű pakolás
DNS - Terminális redundancia, cirkuláris permutáció Általános transzdukció

24
MÁS FÁGOK T4 (virulens) mutánsa Lambda
Mu (transzpozon), DNS-e két végén gazda DNS
, DNS-e két végén gazda DNS.")
25
Transzformáció Pneumococcus (Stapylococcus) Bacillus transzformáció
Kompetencia DNS felvétel/bejutás Donor DNS megtartása a recipiensben Más transzformációs rendszerek Haemophilus influenzae Escherichia coli Elektroporáció Protoplaszt transzformáció Transzfekció – fág DNS-sel E. coli B. subtilis Géntérképezés transzformációval

26
Transzformáció Genetikai információátvitel (az első amit felfedeztek)
Donor sejt nagyméretű DNS-t bocsájt ki A DNS a tápközegben diffundál (a recipiens felé) A DNS transzportálódik sejtfalon, sejtmembránon keresztül a citoplazmába, ahol rekombináció történik Ez a transzformáció genetikai információátvitel, nem a normál sejtek tumorrá történő transzformációja (eukarióták esetén)
 A DNS transzportálódik sejtfalon, sejtmembránon keresztül a citoplazmába, ahol rekombináció történik. Ez a transzformáció genetikai információátvitel, nem a normál sejtek tumorrá történő transzformációja (eukarióták esetén)")
27
Jelentősége? Mivel a DNS könnyen degradálódik, azt gondolták nincs valós szerepe (csak a laboratóriumban) Ezzel szemben számos természetes genetikai transzformációs rendszer létezik: Achrobacter, Azotobacter Bacillus, Butyvibrio, Campylobacter, Clostridium, Hemophilus, Micrococcus, Mycobacterium, Neiserria, Pseudomonas, Streptococcus, Streptomyces, Synechoccus Valamilyen jelentőségnek kell lennie Kimutatták még: Talajba jutattott plazmid és kromoszóma DNS fennmarad, mi több a Pseudomonas stutzeri és a Bacillus subtilis transzformálódik is Egér > DNS etetés után kimutatható a DNS a vérben is, így lehet olyan sejt is , amelyik felveszi ( immunizálás )
 Ezzel szemben számos természetes genetikai transzformációs rendszer létezik: Achrobacter, Azotobacter Bacillus, Butyvibrio, Campylobacter, Clostridium, Hemophilus, Micrococcus, Mycobacterium, Neiserria, Pseudomonas, Streptococcus, Streptomyces, Synechoccus. Valamilyen jelentőségnek kell lennie. Kimutatták még: Talajba jutattott plazmid és kromoszóma DNS fennmarad, mi több a Pseudomonas stutzeri és a Bacillus subtilis transzformálódik is. Egér > DNS etetés után kimutatható a DNS a vérben is, így lehet olyan sejt is , amelyik felveszi ( immunizálás )")
28
Transzformáció felfedezése és utóhatásai
Pneumococcus Sima, poliszacharid burokkal rendelkező telep patogén Durva telep, kapszid poliszacharid hiánya Nem patogén Klasszikus transzformációs kísérlet Avery kimutatja a transzformáló elv a DNS Senki nem akarja elhinni Hershey és Chase DNS és fehérje radioaktív jelölés (külön) Watson és Crick, a DNS szerkezete Pneuomococcus = Streptococcus pneumoniae Bacillus subtilis 168
 Watson és Crick, a DNS szerkezete. Pneuomococcus = Streptococcus pneumoniae. Bacillus subtilis 168.")
29
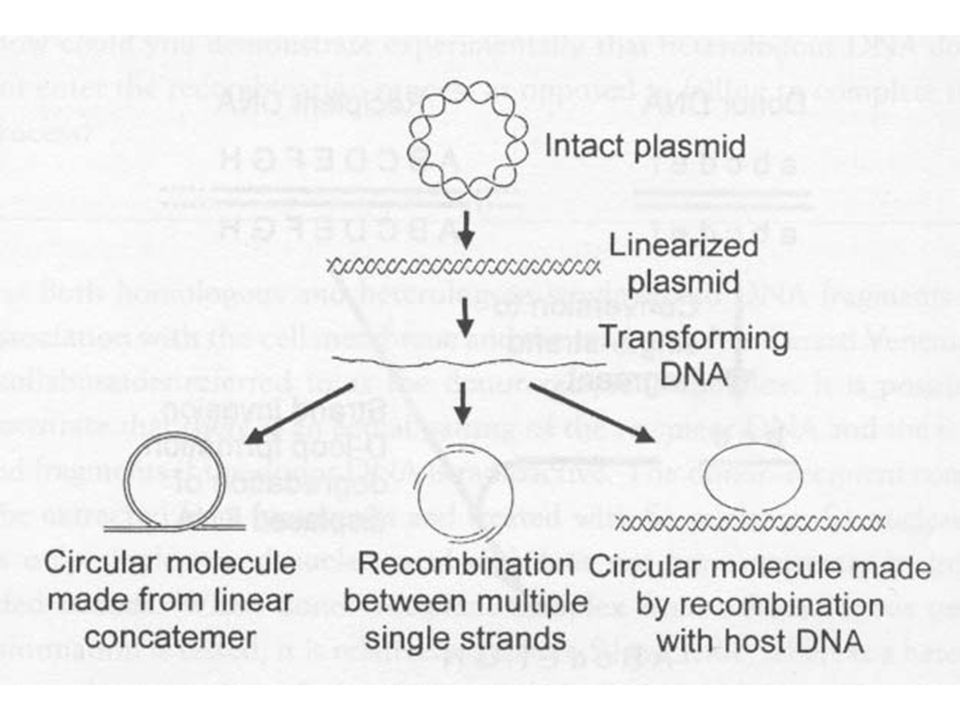
Transzformáció A transzformáció az a folyamat, amikor a sejtek „tiszta” DNS-t vesznek fel és építenek be. Két fő típus Természetes transzformáció (a mikróbáknak csak kis csoportja) - általában lineáris DNS 2. Mesterségesen indukált (a legtöbb, de nem az összes mikroorganizmus képes így DNS felvételre) - általában plazmid DNS Az a sejt, amelyik képes a DNS felvételére az kompetens.
 - általában lineáris DNS. 2. Mesterségesen indukált (a legtöbb, de nem az összes mikroorganizmus képes így DNS felvételre) - általában plazmid DNS. Az a sejt, amelyik képes a DNS felvételére az kompetens.")
30
Mesterséges kompetencia
A molekuláris biológia egyik legfontosabb, alapvető technikája, lehetőség, hogy idegen DNS-t vigyünk be a baktérium gazdába. Gyakran csak úgy érhető el, hogy mesterséges-kompetens sejteket készítünk. Az E. coli felvesz és replikál gyűrűs DNS-t. A mesterséges transzformáció két típusa: Kémiai kompetencia Elektroporáció/elektrotranszformáció, biolisztikus (génpuska) PROTOPLASZT PoliEtilénGlikol = PEG
 PROTOPLASZT PoliEtilénGlikol = PEG.")
31
Kémiai kompetencia Néhány baktériumba, köztük az E. coli-ba, a kétértékű kationok, kis hőmérsékleten segítik a plazmid DNS felvételét ( a lineáris DNS-t is felveszik, de a citoplazma DNáz-ai gyorsan feldarabolják, még mielőtt bármit is tudna tenni) Nem teljesen világos, hogy hogyan is van. Felvételi csatornák: poliP, PHB, és Ca
 Nem teljesen világos, hogy hogyan is van. Felvételi csatornák: poliP, PHB, és Ca.")
32
Elektroporáció Nagy térerősség átmeneti lyukakat eredményez a sejtfalban Alkalmas körülmények között a DNS be/ki szivárog. A kinti nagy plazmid koncentráció gyors beáramlást eredményez. Elektroporációs küvetta Ide kerülnek a sejtek Nagy feszültség sokk

33
A transzformáció hatékonysága?
Transzformációs gyakoriság Sejtszámra (transzformánsok száma/μg DNS) /μg pBR322 kb plazmid/μg pBR322 úgy is mérhető, hogy a sejtek hány %-a kapott plazmidot DNS-re a DNS molekulák hány %-a transzformált sejtet Módszer sejtszámra DNS-re kémiai 1% 12% elektro 10% 90%
 /μg pBR322. kb plazmid/μg pBR322. úgy is mérhető, hogy a sejtek hány %-a kapott plazmidot. DNS-re. a DNS molekulák hány %-a transzformált sejtet. Módszer. sejtszámra. DNS-re. kémiai. 1% 12% elektro. 10% 90%")
34
Természetes kompetencia/transzformáció
gram-pozitívokban Pl.: Streptococcus pneumoniae Bacillus subtilis -nem specifikus -korlátozott számú felvevőhely (30-75) -nick képződés -a komplementer szál degradálódik a felvétel közben -rekombináció a recipiensben
 -nick képződés. -a komplementer szál degradálódik a felvétel közben. -rekombináció a recipiensben.")
35
Természetes transzformáció
gram-negatív Pl.: Haemophilus influenzae Neisseriae gonorrhoeae -szekvencia specifikus -4-8 hely/sejt -nincs sejthez kötött intermedier -dsDNS import a periplazmába -komplementer degradálódik a citoplazmába történő transzport során -rekombináció a recipiensben

36
Gram-pozitív felvevő rendszer
-jó gépezet a sejtekbe történő DNS szállításra a konjugációs rendszer fordítottja - néhány komponense a Tra funkciókhoz hasonlít

37
Gram-negatív felvevő gépezet
-ez se rossz - keresztezni kell a periplazmát és a külső membránt

38
A folyamat energia ellátása?
Intracelluláris ATP hidrolízis pH gradiens – PMF (proton motive force)? komplementer degradáció Funkciója Tápanyag DNS repair Genetikai identitás/diverzitás
 komplementer degradáció. Funkciója. Tápanyag. DNS repair. Genetikai identitás/diverzitás.")
39
Transzformáció Kompetencia kialakulása DNS kötődés és bejutás
A DNS integrációja a recipiens sejtben

40
Kompetencia Az alapelv nagyon hasonló (természetben előforduló)
Kompetencia = az a képesség, mellyel a DNS megkötésre kerül, ezáltal védett lesz a nukleázokkal szemben A kompetencia időzítése különbözik Szinte minden esetben (amit tanulmányoznak) fejlett szabályozás alatt áll a kompetencia
 fejlett szabályozás alatt áll a kompetencia.")
41
A kompetencia fiziológiája
Meghatározott fiziológiai állapot Valamely fejlődési szakaszhoz, vagy tápanyag ellátottsághoz kapcsolódik (legtöbbször, tápanyag hiány kialakulása kapcsolja be) B. subtilis esetében hőmérséklet csökkenés is kiválthatja (42°C>37°C) A szabályozó rendszerben olyan represszorok vannak, melyek egyrészt flagella és kemotaxis géneket másrészt kompetencia géneket aktiválnak
 B. subtilis esetében hőmérséklet csökkenés is kiválthatja (42°C>37°C) A szabályozó rendszerben olyan represszorok vannak, melyek. egyrészt flagella és kemotaxis géneket. másrészt kompetencia géneket aktiválnak.")
42
Kivételek Bakteriofág is indukálhat kompetenciát (Staphylococcus aureus) Exponenciális növekedési szakaszban alakul ki a kompetencia (S. pneumoniae) Antibiotikumok hatására, szubletális koncentráció A tenyészetben lévő kompetens sejtek száma változó (15% Bacillus-Streptococcus, 100%) A kompetencia kialakulása függ a sejtszámtól (sejtsűrüség faktorok, peptidek)
 A kompetencia kialakulása függ a sejtszámtól (sejtsűrüség faktorok, peptidek)")
43
Kompetencia kialakulása rövid ideig tart (pár perc)
Stacioner fázisban Bacillusoknál, ha a kétszereződési idő perc Mivel kemosztát kultúrában is van kompetencia, de nincs sporuláció, ezért a sporuláció nem szükségszerűen kapcsolódik a kompetenciához Hasonlóan a korai sporulációs génekben mutáns baktérium kompetens

44
A B subtilis kompetens sejtek:
Kisebb sűrűségűek (gradiens centrifugálással elkülöníthetők a normál sejtektől) Kisméretűek Egy nem replikálódó genomja van A kompetens sejtek membránja 4* több PHB-t (polihidroxi-butirát) tartalmaz, (egyébként raktározó szerepe van) Segít egy transzmembrán csatornát kialakítani
 Kisméretűek. Egy nem replikálódó genomja van. A kompetens sejtek membránja 4* több PHB-t (polihidroxi-butirát) tartalmaz, (egyébként raktározó szerepe van) Segít egy transzmembrán csatornát kialakítani.")
45
S. penumoniae Kompetens sejtek membránján kompetencia faktor
Ez a közegbe is kikerül Ettől a nem kompetens sejtek kompetenssé válnak, úgy hogy a kompetencia faktor egy másik membrán fehérjéhez köt (comD terméke) A kompetencia faktor kötésével legalább 10 fehérje kezd el szintetizálódni A kompetencia faktor megmagyarázza, miért kell nagy sejtsűrűség, nagyobb a valószínűsége, hogy megtalálja a receptorát A com lókusz mutációi megakadályozzák a kompetencia kialakulását
 A kompetencia faktor kötésével legalább 10 fehérje kezd el szintetizálódni. A kompetencia faktor megmagyarázza, miért kell nagy sejtsűrűség, nagyobb a valószínűsége, hogy megtalálja a receptorát. A com lókusz mutációi megakadályozzák a kompetencia kialakulását.")
46
comA terméke hasonlít az ATP függő transzport fehérjékhez (melyek toxin fehérjéket szállítanak a sejt felszínére, coli hemolizin gén) A kompetencia kialakulásával a sejtfal porózusabbá válik Az autolitikus enzimaktivitás növekszik Baktérium sejtek láncolata hosszabb lesz A legtöbb változás a sejt egyenlítői síkjában következik be, ahol a növekedés történik, és ahol a DNS felvétel történik

47
DNS kötés Kompetens és nem kompetens sejt is köt DNS-t
A nem kompetens sejtben lemosható a DNS, a kompetens sejtben nem mosható le egyszerűen EDTA hatására a DNS megkötődik, de minden további feldolgozás megáll Kb. 50 DNS kötő hely van a kompetens B. subtilis felszínén A kötött DNS érzékeny a nukleázokra és a nyíróerőkre Bármilyen eredetű DNS kötődhet Minimum 500 bp hosszú fragmens kell a kötéshez Egyszálú, glükozilált DNS, ds RNS, RNS-DNS hibrid nem kötődik

48
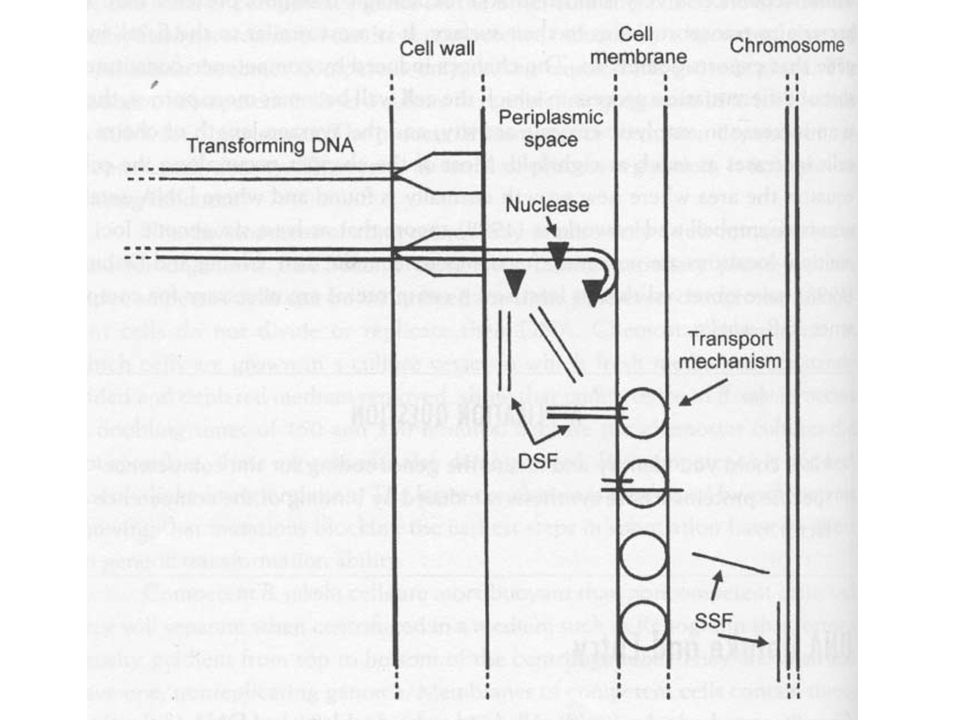
DNS bejutás A következő lépés a DNS bejutása a sejtbe
A 3’ végtől kezdve A bejutás előtt a kötőhely melletti endonukleázok megfelelő méretre hasítják a DNS-t Véletlenszerű hasítás Eredmény 15 kb-nál nem nagyobb DNS fragmensek (kisebb, mint amekkorát a transzdukáló fágok bejuttatnak) Az endonukleázoknak magnéziumra vagy kalciumra van szükség (ezért érzékeny az EDTA-ra) Ebben a fázisban, ha szétválasztjuk a DNS-t és a sejteket a kettős szálú DNS alkalmas a transzformációra
 Az endonukleázoknak magnéziumra vagy kalciumra van szükség (ezért érzékeny az EDTA-ra) Ebben a fázisban, ha szétválasztjuk a DNS-t és a sejteket a kettős szálú DNS alkalmas a transzformációra.")
49
Következő lépés az eklipsz komplex kialakulása
A DNS kötődik egy kompetencia specifikus fehérjéhez Ezáltal a DNS egyszálú lesz Védett lesz a külső nukleáz hasítással szemben Teljesen véletlenszerű, hogy melyik szál hasad le és az is, hogy milyen helynél kezdődik a hasítás A lebomlott szál nukleotidjai a médiumba kerülnek

51
Mi magyarázza a transzformáció jelenségét
Genetikai átváltozás Energia az elbontott DNS szálból Önmagában egyik sem magyarázza, de a kettő együtt igen

52
Elektronmikroszkópos felvételek
Adhéziós zónához hasonló részek a kompetens sejtekben (mint a T4 fág infekciókor) A bakteriális kromoszóma membránhoz kötött, a transzformáló DNS közvetlenül a célhoz jut Normál proton gradiens szükséges a DNS bejutáshoz (B. subtilis), ATP kell Streptococcus-nál A kompetencia indukció része a RecA és egy kapcsolódó fehérje (kolligrin) termelése A kolligrin membrán fehérje a RecA-t segíti a membránban Így a bejutó DNS rögtön rekombinációba lép (mert S. pneumoniae elektroporációval nem transzformálható)
 A bakteriális kromoszóma membránhoz kötött, a transzformáló DNS közvetlenül a célhoz jut. Normál proton gradiens szükséges a DNS bejutáshoz (B. subtilis), ATP kell Streptococcus-nál. A kompetencia indukció része a RecA és egy kapcsolódó fehérje (kolligrin) termelése. A kolligrin membrán fehérje a RecA-t segíti a membránban. Így a bejutó DNS rögtön rekombinációba lép (mert S. pneumoniae elektroporációval nem transzformálható)")
53
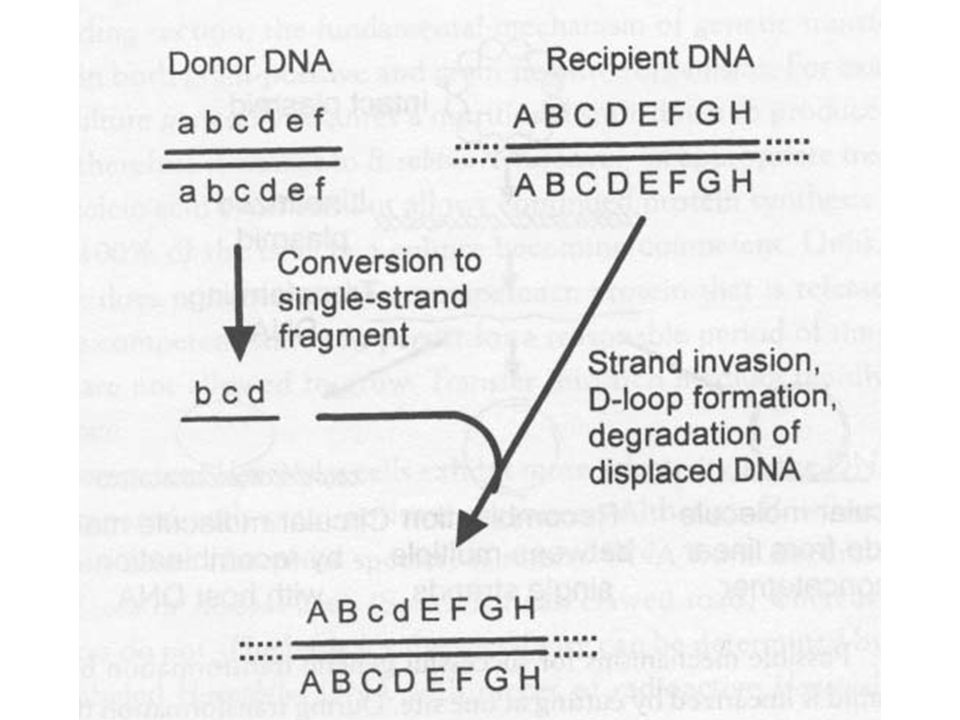
A donor DNS sorsa Kompetens sejt homológ (saját) és heterológ (idegen) DNS-t is köt Mindkettő egyszálú is lesz az internalizációkor Genetikai kicserélődés során kell lenni olyan folyamatnak, mely megakadályozza idegen DNS honosítását Bacillus és Streptococcus a rekombináció során diszkrimiál. Különbséget tesz idegen és saját (faj) DNS-e között
 DNS-e között.")
54
Korrekt hidrogén hídkötés szükséges, ahol nincs ott nincs rekombináció és kizáródik a heterológ DNS
Rekombinációban deficiensnél nincs transzformáció A rekombináció indukciója (nick-ek a recipiens DNS-ben) javítja a transzformáció hatékonyságát
 javítja a transzformáció hatékonyságát.")
56
A javítástól függően elveszhetnek transzformánsok
Az egyszálú DNS D-loop kialakításával kicseréli a recipiens homológ darabját>tranziens heteroduplex alakul ki A hetroduplexben lehet néhány rossz párosodás, ezt a repair enzimek javítják A javítástól függően elveszhetnek transzformánsok S. pneumoniae esetén Hex rendszer A Hex fehérje bizonyos helyeket felismer a genomon A helyek körül kijavítja a hibákat Azok a markerek, amelyek Hex felismerő hellyel rekombinálnak elvesznek LE (low efficiency) Hex helytől távolabbi markerek nem vesznek el HE (high efficiency) a hex limitje a fehérje molekulák száma (ha sok transzformáló DNS-t adunk, telítődik a hex rendszer
 Hex helytől távolabbi markerek nem vesznek el HE (high efficiency) a hex limitje a fehérje molekulák száma (ha sok transzformáló DNS-t adunk, telítődik a hex rendszer.")
57
Plazmid transzformáció
Önállóan képes replikálódni, ha bejutott DE!! A bejutás során linearizálódik Egyszálú DNS-ként jut a citoplazmába, pedig a replikációhoz kétszálúnak kell lennie Egy plazmid molekula nem ad transzformánst, szinte mindig extra DNS is található Hogyan oldható meg, a recipiensben kell homológ DNS (vagy kromoszóma, vagy plazmid) Ha van homológ darab, akkor rekombinálódhat, cirkularizálódhat Ha homológ DNS darab kötődik a vektorhoz, akkor nagy gyakoriságú transzformációt kapunk Marker rescue (ha rezidens plazmidot használunk a transzformánsok visszanyerésének növeléséhez)
 Ha van homológ darab, akkor rekombinálódhat, cirkularizálódhat. Ha homológ DNS darab kötődik a vektorhoz, akkor nagy gyakoriságú transzformációt kapunk. Marker rescue (ha rezidens plazmidot használunk a transzformánsok visszanyerésének növeléséhez)")
59
Más transzformációs rendszerek

60
Hemophilus influenzae
Gram negatív organizmus Kompetencia kialakulása tápanyaghiányra Nukleinsav szintézis gátlás 100% kompetencia (fehérje szintézis esszenciális) Nincs kompetencia specifikus fehérje Csak más Hemophilus fajból vesz fel DNS-t A DNS-en kell lennie egy 11 bp-os darabnak Ez kötődik a sejt membránhoz
 Nincs kompetencia specifikus fehérje. Csak más Hemophilus fajból vesz fel DNS-t. A DNS-en kell lennie egy 11 bp-os darabnak. Ez kötődik a sejt membránhoz.")
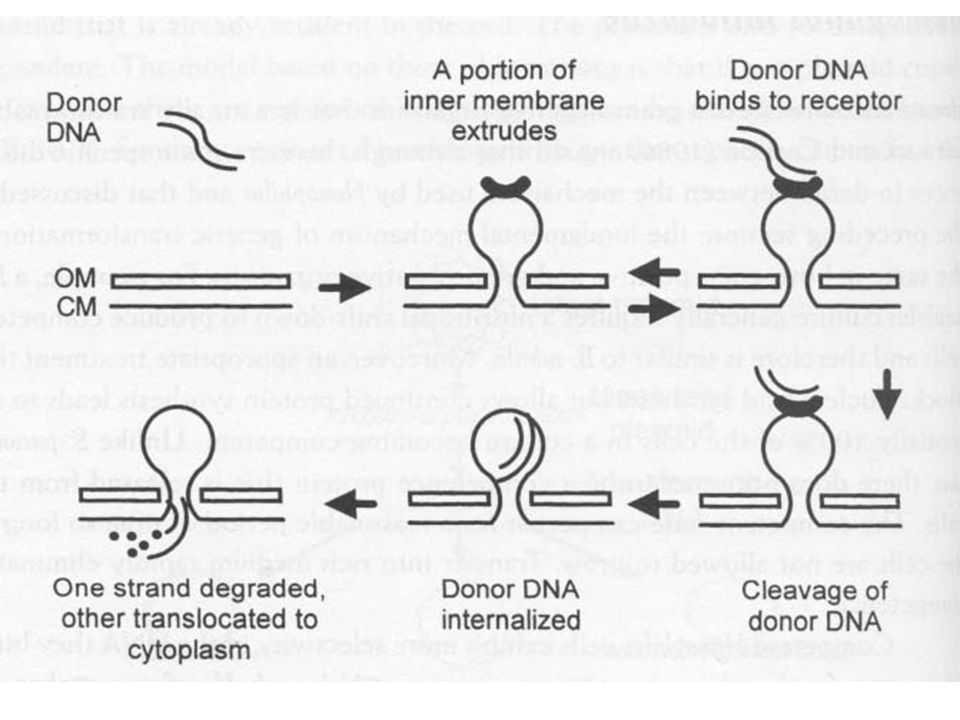
61
DNS bejutása más Belső membrán kitüremkedik, kialakul a transzformaszóm Donor DNS kapcsolódik a receptorhoz Plazmid transzformáció nagyon kis gyakorisággal Noha ds DNS jut be a transzformaszómba a rekombináció egyszálú DNS-el jön létre

63
E. coli transzformáció Természetes körülmények között (spontán) nem transzformálódik Különleges esetekben kompetenssé válik Szferoplaszt Kalcium koncentráció, hősokk (90% sejthalál) Kobalt klorid és DMSO befolyásolja Subtilis-hez hasonlóan PHB halmozódik a membránban (kompetens sejtnél) Szabályozott folyamat, PHB szintézist a Ca++ elősegíti Plazmid DNS-el simán transzformálható, mert nem linearizálódik ezért fontos!!!! (lineáris DNS kisebb hatékonyság –1000*)
 Kobalt klorid és DMSO befolyásolja. Subtilis-hez hasonlóan PHB halmozódik a membránban (kompetens sejtnél) Szabályozott folyamat, PHB szintézist a Ca++ elősegíti. Plazmid DNS-el simán transzformálható, mert nem linearizálódik ezért fontos!!!! (lineáris DNS kisebb hatékonyság –1000*)")
64
Lineáris DNS-el nem transzformálható
Exonukleáz V bontja a DNS-t (ezt a rolling circle replikációhoz ki kell kapcsolni, lambdánál) Legtöbb transzformált coli törzs recB, recC (ExoV inaktív) és sbcB (rekombináció hiány szupresszor) Természetes transzformáció gátjai Nincs kompetencia (spontán) ExoV jelenléte Mindkettőt ki lehet küszöbölni laboratóriumi körülmények között
 Legtöbb transzformált coli törzs recB, recC (ExoV inaktív) és sbcB (rekombináció hiány szupresszor) Természetes transzformáció gátjai. Nincs kompetencia (spontán) ExoV jelenléte. Mindkettőt ki lehet küszöbölni laboratóriumi körülmények között.")
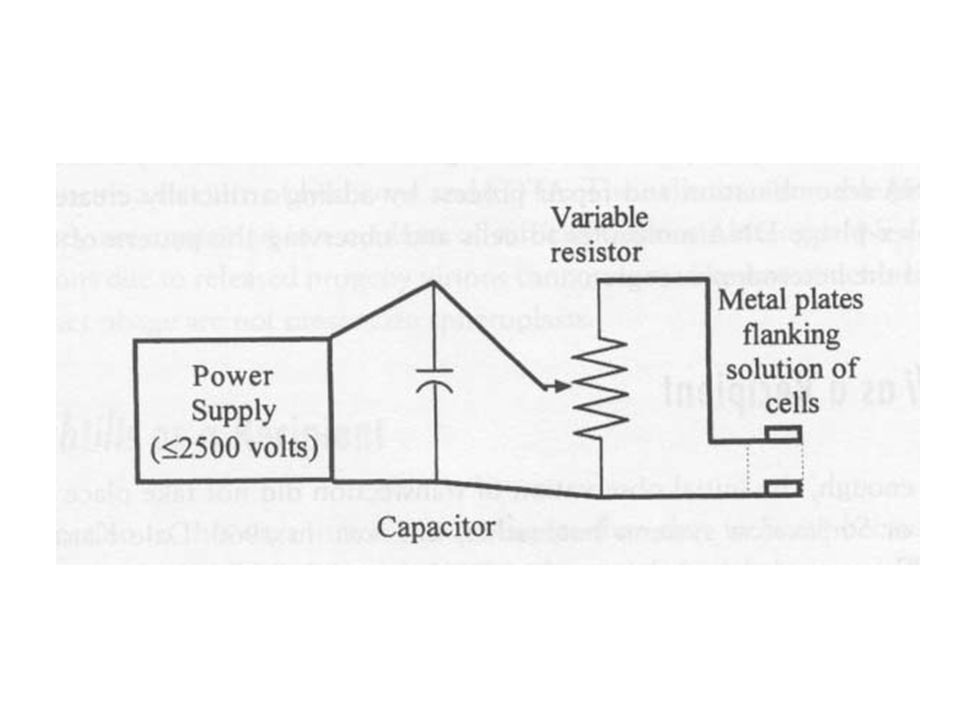
65
Elektroporáció Elektro transzformáció
Nagy feszültségű áram baktérium szuszpenzión keresztül Kis lyukak keletkeznek a sejtfalon Kis méretű plazmidok, vektor konstrukciók átjutnak Bejutás után hasonló, mint a többi Nagy sejtpusztulás, de sok baktériumsejtet lehet alkalmazni, elegendő a legtöbb kísérlethez E. coli esetén a leghatékonyabb (nagyobb transzformációs gyakoriság, nagyobb méretű DNS)
")
67
Transzfekció A donor DNS forrása nem baktérium sejt, hanem bakteriofág
Az eredmény infekciós centrum, azaz plakk A hatékonyság plakk vizsgálattal végezhető Előnye A DNS populáció egységes Így követhető a DNS sorsa a transzformáció lépéseiben Vizsgálható a transzformáció jelensége Hasznos volt a rekombináció és repair tanulmányozásához (mesterséges heteroduplexeket hoztak létre fág és más DNS-el és vizsgálták a rekombinációt, javítást)
")
68
Térképezés transzformációval
Gének kapcsoltságának megállapítása transzformációval Transzdukcióhoz hasonló elv Ha két távoli gént veszünk, azok egyszerre csak akkor transzformálhatnak, ha egy sejt egyszerre két DNS darabot épít be (egyik az egyik, másik a másik génnel) A közeli géneknél (kapcsolt) az együttes transzformánsok úgy is létrejöhetnek, hogy mindkét gén egy fragmensen van
 A közeli géneknél (kapcsolt) az együttes transzformánsok úgy is létrejöhetnek, hogy mindkét gén egy fragmensen van.")
69
A csökkenő mennyiségű DNS-el történő transzformáció során kapcsolt gének esetén a recipiens egyszeres és kétszeres transzformánsainak száma megegyezik (mert a két gén ugyanazon a transzformáló donor DNS-en helyezkedik el) Távoli gének esetén, nagyobb a valószínűsége annak, hogy külön DNS-en vannak, azaz, ha higítjuk a transzformáló DNS-t előbb utóbb, csak az egyik, vagy csak a másik gént tartalmazó DNS-el történik a transzformáció

71
A két gén egyedi transzformációs gyakoriságának meghatározása után a két gyakoriságot összeszorozzuk (ez a várható transzformációs gyakoriság, ha a két gén nem kapcsolt) Ezt az értéket összehasonlítják a kétszeres transzformációs gyakorisággal, ha az utóbbi nagyobb, mint a várt érték, azaz az egyszeres transzformánsok gyakoriságához közelít, akkor a két gén kapcsolt

72
A kapcsoltság így megállapítható
Feltételezés A transzformáló keverékben ismert méretű és azonos nagyságú DNS darabok vannak A donor DNS átlagos nagyságától függ, hogy két gén kapcsolt-e Nagyobb DNS darabok esetén távolabbi gének is kapcsoltnak mutatkoznak A kapcsoltság így megállapítható Ha A gén kapcsolt B-vel, B gén kapcsolt C-vel, akkor a sorrend ABC

73
Géncsere/Genetikai térképezés baktériumokban
Transzformáció, transzdukció és konjugáció

75
Genetikai információ átvitel baktériumokban
Mindíg egy irányú: donorból a recipiensbe Gének sorrendjének meghatározása Kapcsoltság Szelektált/nem szelektált marker Két és három pontos térképezés Genetikai és fizikai térkép

76
B. subtilis transzformációja

79
Térképezés transzformációval:
A rekombinációs gyakoriságokból következtetünk a gének sorrendjére. p+ q+ o+ x p q o Ha p+ és q+ kotranszformációja gyakori, akkor a sorrend p-q-o. Ha p+ és o+ kotranszformációja gyakoribb, akkor p-o-q.

80
Genetikai térképezés transzformációval
a+ b+ Donor sejtből DNS darabolódás a+ a+ b+ b+ a-b- recipiens transzformációja Ha a+b+ kapcsoltak (fizikailag közel vannak egymáshoz), a+b+, a+b- és a-b+ rekombinánsokat kaphatunk. Ha nem szorosan kapcsoltak, akkor csak a+b- vagy a-b+ . Transzformált, transzformáns.
, a+b+, a+b- és a-b+ rekombinánsokat kaphatunk. Ha nem szorosan kapcsoltak, akkor csak a+b- vagy a-b+ . Transzformált, transzformáns.")
81
PROTOPLASZT Baktérium, gomba, növény, állat sejtekből protoplaszt
+ DNS + PEG Protoplaszt készítés sejtfalbontó enzimekkel: lizozim, cellulázok, proteázok, mihez mi kell

82
KÉMIAI KOMPETENCIA Li-acetát S. cerevisiae LiCl P. pastoris
KCl B. subtilis



 DNS-ből,>")







 prokariótában>")


 Bihari Péter.>")







