Előadást letölteni
Az előadás letöltése folymat van. Kérjük, várjon

1
Funkcionális neuroanatómia
12. Az alvás és ébrenlét, aktiválódás, figyelem és az elhatározás neuroanatómiája Neuromorfológia PhD program

2
Johann Friedrich Blumenbach (1752-1840).
Blumenbach examined a young man with an opening in his skull. He observed that his brain seemed to press upon his skull when he was awake and seemed to shrink when he was asleep. Blumenbach concluded that sleep was caused by a diminished flow of blood to the brain. William Howell ( ). Late in the nineteenth century, Howell proposed that sleep could have multiple interacting causes. He looked upon cerebral anemia and blood flow as being especially important factors. Constantin von Economo ( ), who studied patients with viral encephalitis and concluded sleep might be caused by damage to the periventricular gray matter of the brainstem. Finger_page247_fig17-3.jpg + Finger_page247_fig17-3.jpg + Finger_page250_fig17-5.jpg 2
. Late in the nineteenth century, Howell proposed that sleep could have multiple interacting causes. He looked upon cerebral anemia and blood flow as being especially important factors. Constantin von Economo ( ), who studied patients with viral encephalitis and concluded sleep might be caused by damage to the periventricular gray matter of the brainstem. Finger_page247_fig17-3.jpg + Finger_page247_fig17-3.jpg + Finger_page250_fig17-5.jpg. 2.")
3
Reticular formation

4
mesencephalic reticular formation promotes the walking state
Brodal_page334_fig dpi.jpg medullary reticular formation inhibits the walking state (sleep induction)
")
5
kis kép: Brodal_page335_fig16-4-200dpi.jpg

6
Brodal_page339_fig dpi.jpg 6

7
Kandel_page940_fig dpi.jpg The major regions of the brain stem and forebrain involved in sleep control. Stimulation of neurons in the nucleus reticularis pontis oralis/caudalis (RPO/RPC) region produces various characteristics of sleep. Depending on their exact size and location, bilateral lesions within this region completely block REM sleep or block components of REM sleep. CG – central gray; LC – locus ceruleus; LDT – lateral-dorsal tegmental nucleus; PPN – pedunculopontine nucleus. 7
 region produces various characteristics of sleep. Depending on their exact size and location, bilateral lesions within this region completely block REM sleep or block components of REM sleep. CG – central gray; LC – locus ceruleus; LDT – lateral-dorsal tegmental nucleus; PPN – pedunculopontine nucleus. 7.")
8
ébrenlét wakefulness

9
Beckstead_page155_fig13_2-200dpi.jpg 9

10
Spinoreticulothalamic tract „ascending reticular activating system”
A thalamus intralaminaris magjaiból eredő thalamo-corticalis rostok számos kérgi területen végződnek: a prefrontalis kéregben, az elülső cinguláris kéregben, a premotoros areában, az insularis kéregben és a parietális asszociációs area neuronjain. középen: O:\A_PM\Előadások-ppt\ September\Kandel_page342-5valtozat1-ures.jpg + Kandel_page342-5valtozat2-piros.jpg (méret: 17,53x7,12 cm; pozíció: 9,1; 1,47 cm) jobb felső: O:\A_PM\Előadások-ppt\Human brain \rajz sarga.jpg + rajz pirossal.jpg (méret: 6,19x6,6 cm; pozíció: 17,48; 1,74) jobb középső: O:\A_PM\Előadások-ppt\_Fájdalom, 2010\Duvernoy_page23.jpg jobb alsó: 2Babra.jpg 10 10
 jobb felső: O:\A_PM\Előadások-ppt\Human brain \rajz sarga.jpg + rajz pirossal.jpg (méret: 6,19x6,6 cm; pozíció: 17,48; 1,74) jobb középső: O:\A_PM\Előadások-ppt\_Fájdalom, 2010\Duvernoy_page23.jpg. jobb alsó: 2Babra.jpg")
11
The ascending arousal system sends projections from the brainstem and posterior hypothalamus throughout the forebrain. Aminergic nuclei (green circles) diffusely project throughout much of the forebrain, regulating the activity of cortical and hypothalamic targets directly. Neurons of the tuberomammillary nucleus (TMN) contain histamine (HIST), neurons of the raphé nuclei contain 5-HT and neurons of the locus coeruleus (LC) contain noradrenaline (NA). Saper_fig1.jpg (Trends Neurosci. 2001 Dec;24(12): ) Saper CB, Chou TC, Scammell TE. The sleep switch: hypothalamic control of sleep and wakefulness.
: ) Saper CB, Chou TC, Scammell TE. The sleep switch: hypothalamic control of sleep and wakefulness.")
12
Cholinerg sejtcsoportok
Phd Neurotranszmitterek 25. dia Cholinerg sejtcsoportok medial septal Ch1 nucleus of the diagonal band Ch2-Ch3 nucleus basalis (Meynert) Ch4 laterodorsal tegmental nucleus Ch5 pedunculopontine nucleus Ch6 Ch5 and Ch6 cholinerg neurons - coexpress substance P, ANF and CRF, - fibers ascend in the dorsal tegmental bundle, - innervate intralaminar, anterolateral and mediodorsal thalamic nuclei. In wakefulness and REM sleep: cholinerg neurons fire rapidly.
 Ch4. laterodorsal tegmental nucleus Ch5. pedunculopontine nucleus Ch6. Ch5 and Ch6 cholinerg neurons. - coexpress substance P, ANF and CRF, - fibers ascend in the dorsal tegmental bundle, - innervate intralaminar, anterolateral and mediodorsal thalamic nuclei. In wakefulness and REM sleep: cholinerg neurons fire rapidly.")
13
D:\A_PM\Előadások-ppt\071031-Reticular formation\Haines-page092_2012
D:\A_PM\Előadások-ppt\ Reticular formation\Haines-page092_2012.jpg (bal felső) + Haines-page094_2012.jpg (jobb felső) + Haines-page106_2012.jpg (bal alsó) + Haines-page110_2012.jpg (jobb alsó) 13
 + Haines-page094_2012.jpg (jobb felső) + Haines-page106_2012.jpg (bal alsó) + Haines-page110_2012.jpg (jobb alsó) 13.")
14
sleep

15
Kétféle alvás és szinonímái
slow-wave sleep NREM sleep non-REM sleep synchronized sleep S sleep rapid eye movement sleep REM sleep paradoxical sleep desynchronized sleep D sleep

16
brain areas involved in sleep
ascending reticulothalamic system (NREM)
")
17
slow-wave sleep NREM sleep

18
Neurons of the laterodorsal tegmental nuclei and pedunculopontine tegmental nuclei (LDT and PPT) (blue circles) send cholinergic fibers (ACh) to many forebrain targets, including the thalamus (LG+VLP), which then regulate cortical activity. Aminergic nuclei (green circles) diffusely project throughout much of the forebrain, regulating the activity of cortical and hypothalamic targets directly. Neurons of the tuberomammillary nucleus (TMN) contain histamine (HIST), neurons of the raphé nuclei contain 5-HT and neurons of the locus coeruleus (LC) contain noradrenaline (NA). Saper_fig1.jpg (Trends Neurosci. 2001 Dec;24(12): ) Saper CB, Chou TC, Scammell TE. The sleep switch: hypothalamic control of sleep and wakefulness.
 diffusely project throughout much of the forebrain, regulating the activity of cortical and hypothalamic targets directly. Neurons of the tuberomammillary nucleus (TMN) contain histamine (HIST), neurons of the raphé nuclei contain 5-HT and neurons of the locus coeruleus (LC) contain noradrenaline (NA). Saper_fig1.jpg (Trends Neurosci Dec;24(12): ) Saper CB, Chou TC, Scammell TE. The sleep switch: hypothalamic control of sleep and wakefulness.")
19
Non-REM sleep Midbrain reticular formation neurons inhibited.
The generated rhythmic firing of thalamic relay neurons is depleted(a result of GABAergic inhibitory neurons in the nucleus reticularis) = hyperpolarization of thalamo-cortical neurons. EEG spindles, low waves produced by synchronized synaptic potentials in cortical neurons = NREM sleep.
 = hyperpolarization of thalamo-cortical neurons. EEG spindles, low waves produced by synchronized synaptic potentials in cortical neurons = NREM sleep.")
20
NREM sleep jellemzői neuronal activity is low, parasympathetic activity predominates, slightly reduced pulmonary (alveolar) ventillation, declined heart rate and blood pressure, increased gastrointestinal motility, deminished kidney filtration, declined basal metabolic rate, falled body temperature, decreased sympathetic outflow, intact muscle tone and reflexes

21
Non-REM sleep stages stage 1) Transition from wakefulness to sleep
mixed frequencies – low voltage no rapid eye movements some skeletal muscle activities stage 2) Sleep spindles (sinusoidal waves) high amplitude low voltage stage 3) High amplitude – (reflects activity in the prefrontal cortex) slow delta waves stage 4) Increased slow-wave activities loss of muscle tone
 Sleep spindles (sinusoidal waves) high amplitude. low voltage. stage 3) High amplitude – (reflects activity in the prefrontal cortex) slow delta waves. stage 4) Increased slow-wave activities. loss of muscle tone.")
22
NREM sleep EEG hullámai
felső: Beckstead_page158_fig13_5-200dpi.jpg alsó: Muzur_fig1.jpg (Trends Cogn Sci. 2002 Nov 1;6(11): ) Muzur A, Pace-Schott EF, Hobson JA. The prefrontal cortex in sleep.
: ) Muzur A, Pace-Schott EF, Hobson JA. The prefrontal cortex in sleep.")
23
REM sleep

24
brain areas involved in sleep
ascending reticulothalamic system (NREM) pontine-geniculo-occipital (PGO) axis (REM)
 pontine-geniculo-occipital (PGO) axis (REM)")
25
REM sleep jellemzői neuronal activity is high in the pontine reticular formation, lateral geniculate body, occipital cortex neurons (PGO spikes), overall increase in neuronal activity during REM sleep, muscle tone is generally reduced, except eye movements, breathing, dreaming, changes of heart rate and blood pressure, EEG enters a desynchronized pattern, - higher metabolic rate and body temperature, pupils become highly constricted (miosis), rapid eye movements, respiration is relatively unresponsible to blood CO2, reduced responses to heat and cold (amibent body temperature), penile erections.
, rapid eye movements, respiration is relatively unresponsible to blood CO2, reduced responses to heat and cold (amibent body temperature), penile erections.")
26
REM sleep sejttípusai non-REM-on cells - anterior hypothalamus - sleep generator REM-waking-on cells - brain stem reticular formation - active in waking and REM (excite motor neurons) PGO-on cells - pontine reticular formation REM-off cells - biogenic amine cells REM-on cells - pontine reticular formation - very active in REM stages
 PGO-on cells - pontine reticular formation. REM-off cells - biogenic amine cells. REM-on cells - pontine reticular formation - very active in REM stages.")
27
Kandel_page940_fig dpi.jpg

28
NREM – REM sleep cycle NREM 1-4 (70-80 min)
NREM 3-2 (15-20 min) min x 6/night REM (5-10 min) adults: 50-60% in NREM stage 2 REM sleep: 20-25% 15-20% in NREM stage 3+4 5 % in NREM stage 1
 min x 6/night. REM (5-10 min) adults: 50-60% in NREM stage 2 REM sleep: 20-25% 15-20% in NREM stage % in NREM stage 1.")
29
REM-off cells. Aminergic nuclei (green circles) diffusely project throughout much of the forebrain, regulating the activity of cortical and hypothalamic targets directly. Neurons of the tuberomammillary nucleus (TMN) contain histamine (HIST), neurons of the raphé nuclei contain 5-HT and neurons of the locus coeruleus (LC) contain noradrenaline (NA). Saper_fig1.jpg (Trends Neurosci. 2001 Dec;24(12): ) Saper CB, Chou TC, Scammell TE. The sleep switch: hypothalamic control of sleep and wakefulness.
: ) Saper CB, Chou TC, Scammell TE. The sleep switch: hypothalamic control of sleep and wakefulness.")
30
brain areas involved in sleep
ascending reticulothalamic system (NREM) pontine-geniculo-occipital (PGO) axis (REM) circadian rhythm
 pontine-geniculo-occipital (PGO) axis (REM) circadian rhythm.")
31
Suprachiasmatic nucleus embedded in the upper surface of the optic chiasm in the hypothalamus. It participates in setting the normal sleep-wake cycle through connections with the pineal gland. Suprachiasmatic neurons works as an internal cirdadian pacemaker. It receives input from the retina (retino-hypothalamic tract). The suprachiasmatic nucleus regulates the timing of sleep, it is not responsible for sleep itself. Turlough_page294_Sleeping.jpg 31
. The suprachiasmatic nucleus regulates the timing of sleep, it is not responsible for sleep itself. Turlough_page294_Sleeping.jpg. 31.")
32
Regul_food_int_fig2.jpg 32

33
brain areas involved in sleep
ascending reticulothalamic system (NREM) pontine-geniculo-occipital (PGO) axis (REM) circadian rhythm hypothalamic flip-flop switch ventrolateral preoptic nucleus
 pontine-geniculo-occipital (PGO) axis (REM) circadian rhythm. hypothalamic flip-flop switch. ventrolateral preoptic nucleus.")
34
non-REM-on cells. Sleep-promoting neurons of the ventrolateral preoptic nucleus (VLPO, red circle) contain GABA and galanin (Gal). These cells produce sleep by inhibition histaminergic cells in the posterior hypothalamus and cells in the midbrain reticular formation. Saper_fig1.jpg (Trends Neurosci. 2001 Dec;24(12): ) Saper CB, Chou TC, Scammell TE. The sleep switch: hypothalamic control of sleep and wakefulness.
: ) Saper CB, Chou TC, Scammell TE. The sleep switch: hypothalamic control of sleep and wakefulness.")
36
The projections from the ventrolateral preoptic nucleus (VLPO) to the main components of the ascending arousal system and block their activity (non-REM-on cells). Axons from the VLPO directly innervate the cell bodies and proximal dendrites of neurons in the major monoamine arousal groups. Within the major cholinergic groups, axons from the VLPO mainly innervate interneurons, rather than the principal cholinergic cells. Abbreviations: LC, locus coeruleus; LDT, laterodorsal tegmental nuclei; PPT, pedunculopontine tegmental nuclei; TMN, tuberomammillary nucleus; VLPO, ventrolateral preoptic nucleus. The blue circle indicates neurons of the LDT and PPT; green circles indicate aminergic nuclei; and the red circle indicates the VLPO. Saper_fig2.jpg

37
brain areas involved in sleep
ascending reticulothalamic system (NREM) pontine-geniculo-occipital (PGO) axis (REM) circadian rhythm hypothalamic flip-flop switch ventrolateral preoptic nucleus tuberomamillary nucleus
 pontine-geniculo-occipital (PGO) axis (REM) circadian rhythm. hypothalamic flip-flop switch. ventrolateral preoptic nucleus. tuberomamillary nucleus.")
38
Lesions of the posterior hypothalamic area may cause hypersomnolence or even coma. This area contains the tuberomammillary nucleus, housing hundreds of histaminergic neurons, which project widely to the gray matter of the brain and spinal cord. Histaminergic fibers destined for the cerebral cortex. They branch within the superficial layers of the frontal cortex, and run back to supply the cortex of the parietal, occipital, and temporal lobes. Tuberomammillary histaminerg neurons activate H1 receptors on cortical neurons. The tuberomammillary nucleus play an important role in the arousal mechanism, and activated during the awake state by orexin. Turlough_page294_Sleeping.jpg 38

40
brain areas involved in sleep
ascending reticulothalamic system (NREM) pontine-geniculo-occipital (PGO) axis (REM) circadian rhythm hypothalamic flip-flop switch ventrolateral preoptic nucleus tuberomamillary nucleus orexin (and MCH) neurons
 pontine-geniculo-occipital (PGO) axis (REM) circadian rhythm. hypothalamic flip-flop switch. ventrolateral preoptic nucleus. tuberomamillary nucleus. orexin (and MCH) neurons.")
41
Orexin neurons in the lateral hypothalamic area innervate all of the components of the ascending arousal system, as well as the cerebral cortex (CTX) itself. Excitatory, they may help maintain wakefulness by increasing the activity of the ascending arousal system Orexin neurons are wake-active Orexin enurons might influence both sides of the flip-flop circuit by direct projections to both sides Abbreviations: BF, basal forebrain cholinergic nuclei; LC, locus coeruleus; LDT, laterodorsal tegmental nuclei; PPT, pedunculopontine tegmental nuclei; TMN, tuberomammillary nucleus. Blue circles indicate cholinergic neurons of the BF, LDT and PPT; green circles indicate monoaminergic nuclei. Saper_fig4.jpg

42
A model for reciprocal interactions between sleep- and wake-promoting brain regions, which produces a flip–flop switch. Inhibitory pathways are shown in red, and the excitatory pathways in green. The blue circle indicates cholinerg neurons of the LDT and PPT; green boxes indicate aminergic nuclei; and the red box indicates the VLPO. Aminergic regions such as the TMN, LC and DR promote wakefulness by direct excitatory effects on the cortex and by inhibition of sleep-promoting neurons of the VLPO. During sleep, the VLPO inhibits amine-mediated arousal regions through GABAergic and galaninergic (GAL) projections. This inhibition of the amine-mediated arousal system disinhibits VLPO neurons, further stabilizing the production of sleep. The extended VLPO (eVLPO) might promote REM sleep by disinhibiting the PPT–LDT; its axons innervate interneurons within the PPT–LDT, as well as aminergic neurons that normally inhibit REM-promoting cells in the PPT–LDT. - Orexin/hypocretin neurons (ORX) in the lateral hypothalamic area (LHA) might further stabilize behavioral state by increasing the activity of aminergic neurons, thus maintaining consistent inhibition of sleep-promoting neurons in the VLPO and REM-promoting neurons in the PPT–LDT. Abbreviations: DR, dorsal raphé nucleus; HIST, histamine; LC, locus coeruleus; LDT, laterodorsal tegmental nuclei; PPT, pedunculopontine tegmental nuclei; REM, rapid eye movement; TMN, tuberomammillary nucleus; VLPO, ventrolateral preoptic nucleus. Saper_fig3.jpg
 projections. This inhibition of the amine-mediated arousal system disinhibits VLPO neurons, further stabilizing the production of sleep. The extended VLPO (eVLPO) might promote REM sleep by disinhibiting the PPT–LDT; its axons innervate interneurons within the PPT–LDT, as well as aminergic neurons that normally inhibit REM-promoting cells in the PPT–LDT. - Orexin/hypocretin neurons (ORX) in the lateral hypothalamic area (LHA) might further stabilize behavioral state by increasing the activity of aminergic neurons, thus maintaining consistent inhibition of sleep-promoting neurons in the VLPO and REM-promoting neurons in the PPT–LDT. Abbreviations: DR, dorsal raphé nucleus; HIST, histamine; LC, locus coeruleus; LDT, laterodorsal tegmental nuclei; PPT, pedunculopontine tegmental nuclei; REM, rapid eye movement; TMN, tuberomammillary nucleus; VLPO, ventrolateral preoptic nucleus. Saper_fig3.jpg.")
43
álom – dreams

44
Finger_page256_fig18-1.jpg The Sphinx at Giza showing the stone stela at its base telling how Thutmes IV listened to the god Hormokhu in a dream. Thutmes IV cleared the sand covering the great Sphinx, as requested by Hormokhu, and was granted prosperity. Thutmes IV lived approximately 1450 B.C. 44

45
Muzur_fig3.jpg (Trends Cogn Sci. 2002 Nov 1;6(11):475-481.)
Muzur A, Pace-Schott EF, Hobson JA. The prefrontal cortex in sleep.

46
Dreaming Dreaming only in REM sleep Strong activation of the
1) anterior cingulate cortex (dream movement and emotion) 2) premotor areas 3) motor cortex (rapid eye movement, sleep-behavior) 4) basal ganglia (fictive motion of dreams) Reciprocal inhibition between frontal and limbic areas (inhibitory functions are deactivated) Prefrontal cortex deactivation → bizarreness of dreams
 anterior cingulate cortex (dream movement and emotion) 2) premotor areas. 3) motor cortex (rapid eye movement, sleep-behavior) 4) basal ganglia (fictive motion of dreams) Reciprocal inhibition between frontal and limbic areas (inhibitory functions are deactivated) Prefrontal cortex deactivation → bizarreness of dreams.")
47
felébredés – kezdeményezés arousal – response initiation
Prefrontal cortex: - initiation of goal-directed behavior delayed responses - attention distractibility (reduced ability to continuous attention and activity) - selection of specific behavior among several others lesions lesions
 - selection of specific behavior among several others. lesions. lesions.")
48
② ① ① Elülső cingularis és dorsomedialis prefrontális kéreg
Spinoreticulothalamic tract „ascending reticular activating system” ① Elülső cingularis és dorsomedialis prefrontális kéreg ② Középső cingularis kéreg → akcióra irányított figyelem, megelőzés ② ① bal: D:\A_PM\Előadások-ppt\Human brain \ rajz1_ jpg jobb: O:\A_PM\Előadások-ppt\ September\Kandel_page342-5valtozat1-ures.jpg + Kandel_page342-5valtozat2-piros.jpg (méret: 17,53x7,12 cm; pozíció: 9,1; 1,47 cm) 48 48
")
49
dorsolateral prefrontal cortex
Emotional labeling of stimuli, goal-directed behavior, movement Executive functions, logic planning bal: O:\A_PM\Előadások-ppt\Human brain \rajz-2 sorozat\ rajz2-prefrontal.jpg (71%, pozíció: 0,7; 3,22 cm) jobb: D:\A_PM\Előadások-ppt\ Siófok-Depression\scan080604\ Duvernoy-fig40B-sarga2.jpg (méret: 163%; pozíció: 12,61; 0,56 cm) 49
 jobb: D:\A_PM\Előadások-ppt\ Siófok-Depression\scan080604\ Duvernoy-fig40B-sarga2.jpg (méret: 163%; pozíció: 12,61; 0,56 cm) 49.")
50
téli alvás

51
PM631_JCompNeurol505_443.pdf \ Brati_fig2.jpg
c-Fos expression in the preoptic area during hibernation. In ART, low level of c-fos expression was detected in neurons throughout the entire preoptic area (B), while in ET the ventrolateral subdivision of the preoptic area exhibited stronger c-fos expression (C). In animals during torpor (LT) the area remained silent (D). During arousal (EA and LA) we found activity in the ventromedial subdivision of the preoptic area (E,F), and in IBA animals, strong c-fos activity was detected all throughout the area (G). PM631_JCompNeurol505_443.pdf \ Brati_fig2.jpg 51
, while in ET the ventrolateral subdivision of the preoptic area exhibited stronger c-fos expression (C). In animals during torpor (LT) the area remained silent (D). During arousal (EA and LA) we found activity in the ventromedial subdivision of the preoptic area (E,F), and in IBA animals, strong c-fos activity was detected all throughout the area (G). PM631_JCompNeurol505_443.pdf \ Brati_fig2.jpg. 51.")
52
PM631_JCompNeurol505_443.pdf \ Brati_fig3.jpg
c-Fos expression in the suprachiasmatic nucleus during hibernation. The SCN showed low activity in ART (B), while during torpor, c-fos expression in the neurons of the SCN became progressively stronger (C–E) peaking in EA (E) and returning to lower levels of expression in LA (F) and IBA (G). PM631_JCompNeurol505_443.pdf \ Brati_fig3.jpg 52
, while during torpor, c-fos expression in the neurons of the SCN became progressively stronger (C–E) peaking in EA (E) and returning to lower levels of expression in LA (F) and IBA (G). PM631_JCompNeurol505_443.pdf \ Brati_fig3.jpg. 52.")
53
PM631_JCompNeurol505_443.pdf \ Brati_fig4.jpg
c-Fos expression in the reticular thalamic nucleus and the somatomotor cortex during hibernation. The reticular thalamic nucleus showed no activity in awake animals (ART and IBA) (B,D), meanwhile during the torpor phase of the hibernation bout (LT), strong c-fos expression was detected in the nucleus (C). In contrast, activated neurons were found in the somatomotor cortex in only awake animals (ART and IBA) (E,G), but not during torpor (LT) (F). PM631_JCompNeurol505_443.pdf \ Brati_fig4.jpg 53
 (B,D), meanwhile during the torpor phase of the hibernation bout (LT), strong c-fos expression was detected in the nucleus (C). In contrast, activated neurons were found in the somatomotor cortex in only awake animals (ART and IBA) (E,G), but not during torpor (LT) (F). PM631_JCompNeurol505_443.pdf \ Brati_fig4.jpg. 53.")
54
PM631_JCompNeurol505_443.pdf \ Brati_fig5.jpg
c-Fos expression in the paraventricular nucleus during hibernation. The PVN showed no c-fos activity in ART (B), in ET (C), and in LT (D); however, in IBA strong c-fos expression was detected mainly in the parvocellular neurons of the nucleus (E). PM631_JCompNeurol505_443.pdf \ Brati_fig5.jpg 54
, in ET (C), and in LT (D); however, in IBA strong c-fos expression was detected mainly in the parvocellular neurons of the nucleus (E). PM631_JCompNeurol505_443.pdf \ Brati_fig5.jpg. 54.")
55
PM631_JCompNeurol505_443.pdf \ Brati_fig6.jpg
c-Fos expression in the choroid plexus and the ependymal cells of the lateral ventricle during hibernation. Neither the epithelial cells of the choroid plexus nor the ependymal cells of the lateral ventricle showed any c-fos expression in awake animals (ART and IBA) (B,G). During torpor, we detected increasing c-fos expression in the choroid plexus, starting in ET (C), peaking in EA (E), and diminishing in LA (F), while the ependymal cells of the lateral ventricle exhibited activity only in the arousal phases (EA and LA) (E,F). PM631_JCompNeurol505_443.pdf \ Brati_fig6.jpg 55
 (B,G). During torpor, we detected increasing c-fos expression in the choroid plexus, starting in ET (C), peaking in EA (E), and diminishing in LA (F), while the ependymal cells of the lateral ventricle exhibited activity only in the arousal phases (EA and LA) (E,F). PM631_JCompNeurol505_443.pdf \ Brati_fig6.jpg. 55.")
56
PM631_JCompNeurol505_443.pdf \ Brati_fig8.jpg
c-Fos expression in tanycytes and the ependymal cells of the third ventricle during hibernation. No c-fos activity was detected in ART (B) and ET (C) phases. Strong c-fos expression was observed in both tanycytes and third ventricular ependymal cells during LT (D) and EA (E), while this activity diminished in LA (F). Interestingly, neurons in the arcuate nucleus and in the dorsomedial hypothalamus exhibited strong activity in IBA (G). PM631_JCompNeurol505_443.pdf \ Brati_fig8.jpg 56
 and ET (C) phases. Strong c-fos expression was observed in both tanycytes and third ventricular ependymal cells during LT (D) and EA (E), while this activity diminished in LA (F). Interestingly, neurons in the arcuate nucleus and in the dorsomedial hypothalamus exhibited strong activity in IBA (G). PM631_JCompNeurol505_443.pdf \ Brati_fig8.jpg. 56.")
58
Formatio reticularis Az agytörzs integráló rendszere, ahol számos agyterületről (cortex, cerebellum, törzsdúcok, gerincvelő) érkező információ feldolgozása történik. Számos reflex (légzés, keringés, alvás) és projekciós pályarendszer útján a kérgi tevékenységek aktiválásával („felszálló reticuláris aktiváló rendszer”), agytörzsi autonóm központok, továbbá az izomtónus reakciókészségének emelésével az adekvát szomatikus válaszok kidolgozásáért és megvalósításáért felelős neuronok összessége.
 érkező információ feldolgozása történik. Számos reflex (légzés, keringés, alvás) és projekciós pályarendszer útján a kérgi tevékenységek aktiválásával („felszálló reticuláris aktiváló rendszer ), agytörzsi autonóm központok, továbbá az izomtónus reakciókészségének emelésével az adekvát szomatikus válaszok kidolgozásáért és megvalósításáért felelős neuronok összessége.")
59
Nucleus pedunculopontinus
A középagy és a híd határán, a colliculus inferiortól ventrálisan, a tegmentumban levő mag, mely magába foglal egy cholinerg sejtcsoportot (Ch6) is, mely a thalamus beidegzésében vesz részt. Szerepet tulajdonítanak neki a fázisos REM („rapid-eye-movement”) alvásban. Közvetlen rostokat kap a törzsdúcokból és a kisagyból. Részben átfed a „középagyi motoros régió” („mesencephalic locomotor region”) területével. A reticulospinalis pályához küld rostokat. A járás folyamatosságának fenntartásában van szerepe. Sérülése, illetve a törzsdúcokkal való kapcsolatának megszakadása lehet felelős a Parkinson-kórra jellemző járási zavaroknak és rigiditásnak.
 is, mely a thalamus beidegzésében vesz részt. Szerepet tulajdonítanak neki a fázisos REM („rapid-eye-movement ) alvásban. Közvetlen rostokat kap a törzsdúcokból és a kisagyból. Részben átfed a „középagyi motoros régió („mesencephalic locomotor region ) területével. A reticulospinalis pályához küld rostokat. A járás folyamatosságának fenntartásában van szerepe. Sérülése, illetve a törzsdúcokkal való kapcsolatának megszakadása lehet felelős a Parkinson-kórra jellemző járási zavaroknak és rigiditásnak.")
60
Nucleus reticularis thalami
Vékony, C-alakú mag, mely körbeveszi a thalamus felső és oldalsó részét, valamint elülső pólusát. Sejtjei a thalamus külső lemeze és a capsula interna között helyezkednek el. Számos axon-kollaterálist kap a cortico-thalamikus pályákon át szinte valamennyi kérgi areából. Kétirányú kapcsolatban van az elülső magcsoport kivételével a thalamus összes magjával, a törzsdúcokkal és a formatio reticularisszal. Sejtjei GABAerg gátló neuronok, s bár nincs közvetlen projekciója a kéregbe, a thalamus-magok gátlásával szinte egy funkcionális gát szerepét tölti be a thalamo-corticális kapcsolatokban.

61
Nucleus suprachiasmaticus
A hypothalamus elülső részében, a harmadik agykamra két oldalán alul, közvetlenül a chiasma opticum felett helyezkedik el. A napszaki ritmus fenntartásában van szerepe. „Biológiai órá”-nak nevezik – bár helyesebb lenne pacemaker-nek hivni. Rostokat kap a retinából (tractus retino-hypothalamicus), valamint a corpus geniculatum lateralis speciális sejtcsoportjából és a középagyi raphe magoktól. A nucleus suprachiasmaticus rostokat küld a hypothalamus paraventriculáris magjához, a thalamushoz, a törzsdúcokhoz és multiszinaptikus pályákon át a locus coeruleushoz (alvás-ébrenlét szabályozása) és a corpus pinealéhoz (melatonin szintézis).
, valamint a corpus geniculatum lateralis speciális sejtcsoportjából és a középagyi raphe magoktól. A nucleus suprachiasmaticus rostokat küld a hypothalamus paraventriculáris magjához, a thalamushoz, a törzsdúcokhoz és multiszinaptikus pályákon át a locus coeruleushoz (alvás-ébrenlét szabályozása) és a corpus pinealéhoz (melatonin szintézis).")
62
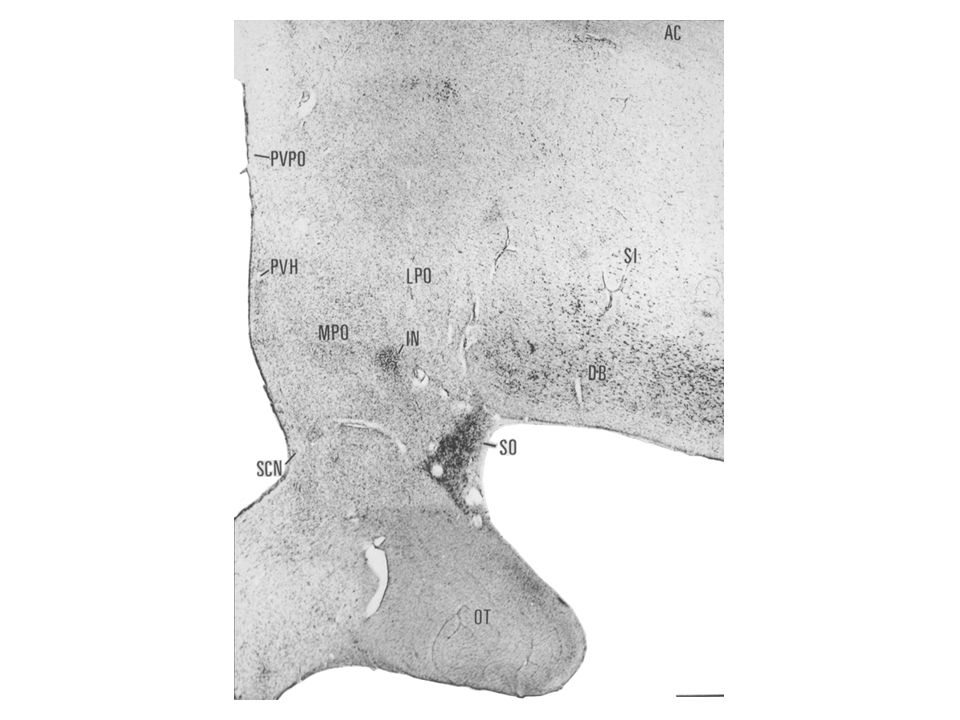
Regio preoptica – area preoptica
A hypothalamus elülső része, a commissura anterior kereszteződése előtti és alatti terület. Közvetlen idegi kapcsolatban van a hippocampusszal („precommissuralis fornix rostok”) és az amygdalával. Sejtjeinek egy része gonadotropin-releasing hormont termel, és az eminentia medianába projíciál. A régió mediális magjának (nucleus preopticus medialis) egyik szubdivíziója a szexuális dimorfizmus központja. A régióban helyezkedik el az agy thermoregulációs központja, valamint a terület ventrolaterális részében találhatók az alvás mechanizmusában szerepet vivő GABAerg neuronok (VLPO).
 és az amygdalával. Sejtjeinek egy része gonadotropin-releasing hormont termel, és az eminentia medianába projíciál. A régió mediális magjának (nucleus preopticus medialis) egyik szubdivíziója a szexuális dimorfizmus központja. A régióban helyezkedik el az agy thermoregulációs központja, valamint a terület ventrolaterális részében találhatók az alvás mechanizmusában szerepet vivő GABAerg neuronok (VLPO).")
63
Retino-hypothalamikus pálya
A retina ganglionsejtjeiből eredő pálya. Rostjai a nervus opticuson belül futnak, a koponyaüregbe való belépés után a chiasma opticumban kereszteződnek és zömmel a nucleus suprachiasmaticusban végződnek.

Hasonló előadás










 MORE?” symposium Washington.>")










