Előadást letölteni
1
A sejtciklus és szabályozása

2
A sejt teória kialakulása
A sejt szerkezeti, fiziológiai és szerveződési egysége minden élő rendszernek. A sejt létezhet önálló egységként vagy szervezetek felépítő egységeként. A sejtek más sejtek közreműködése nélkül a kristályok keletkezéséhez hasonlóan jöhetnek létre. (spontán kialakulás) 1838 Matthias Schleiden Theodor Schwann Omnis cellula e cellula (minden sejt egy másik hasonló létező sejtből származik) 1858. Omnis cellula e cellula (minden sejt egy másik hasonló létező sejtből származik) 1858. A modern sejt teória Minden ismert élő rendszer sejtekből áll. A sejt minden élő rendszernek strukturális és funkcionális egysége. Minden sejt osztódással keletkezik egy másik sejtből. A sejtek örökítő információja sejtről sejtre adódnak át az osztódás során. Minden sejt alapvetően azonos kémiai felépítésű. Az élő rendszereket jellemző energia áramlás és átalakítás a sejtekben történik. Robert Virchow
 Matthias Schleiden Theodor Schwann. Omnis cellula e cellula (minden sejt egy másik hasonló létező sejtből származik) Omnis cellula e cellula (minden sejt egy másik hasonló létező sejtből származik) A modern sejt teória. Minden ismert élő rendszer sejtekből áll. A sejt minden élő rendszernek strukturális és funkcionális egysége. Minden sejt osztódással keletkezik egy másik sejtből. A sejtek örökítő információja sejtről sejtre adódnak át az osztódás során. Minden sejt alapvetően azonos kémiai felépítésű. Az élő rendszereket jellemző energia áramlás és átalakítás a sejtekben történik. Robert Virchow.")
3
A sejtciklus = két periódus szabályos váltakozása a sejtek életében
- a sejttartalom megkettőzése (interfázis) - kettéosztódás (M-fázis) M-fázis interfázis
 - kettéosztódás (M-fázis) M-fázis. interfázis.")
4
A sejtciklus áttekintése I.
Pontosan szabályozott DNS tartalom - replikáció + szétválás és (kromatin) (kromoszóma) centroszóma S fázisban mitózis alatt genetikailag azonos sejtek Nem pontosan szabályozott citoplazma - növekedés + osztódás G1 és G citokinézis M-fázis = mitózis + citokinézis G2 G1 S Interfázis = G1+ S+ G2
 (kromoszóma) centroszóma S fázisban mitózis alatt. genetikailag azonos sejtek. Nem pontosan szabályozott. citoplazma - növekedés + osztódás. G1 és G2 citokinézis. M-fázis = mitózis + citokinézis. G2. G1. S. Interfázis = G1+ S+ G2.")
5
A sejtciklus áttekintése II.
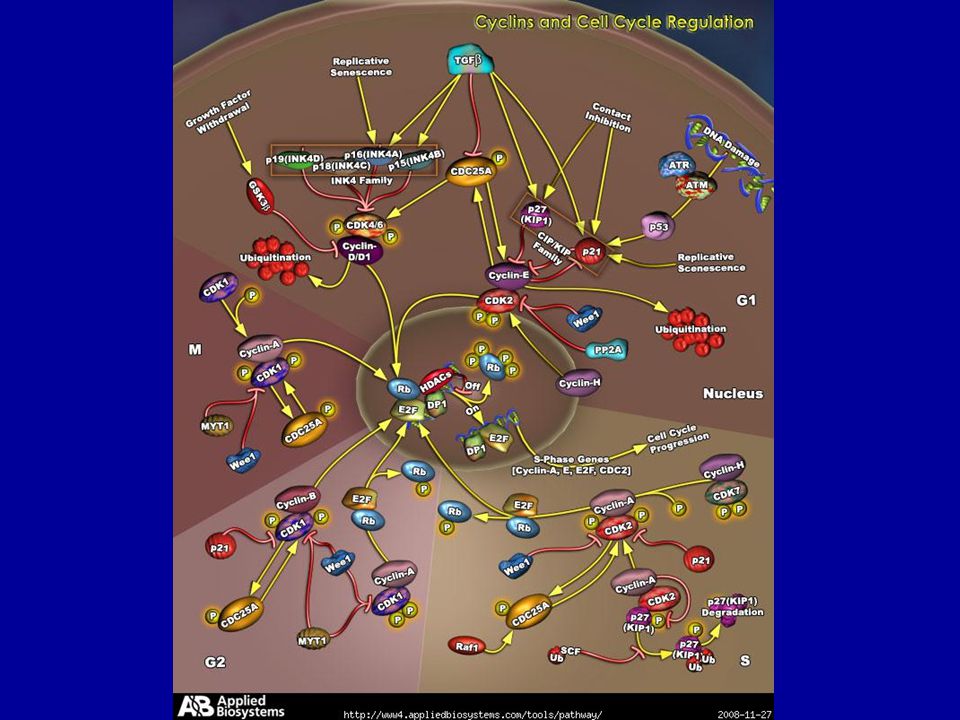
események, folyamatok jól ismertek hogy regulálódik? sejtciklus szabályozó rendszer ciklin dependens kinázok (Cdk-s) ciklinek foszfatázok egyéb kinázok (Aurora, Polo) ciklin dependens kináz gátlók (CKI) ubiquitin ligáz ellenőrzési pontok: G1,(S),G2,M univerzális és konzervatív 3 típus: korai embrionális standard emlős (soksejtű) M G2 G1
 ciklinek. foszfatázok. egyéb kinázok (Aurora, Polo) ciklin dependens kináz gátlók (CKI) ubiquitin ligáz. ellenőrzési pontok: G1,(S),G2,M. univerzális és konzervatív. 3 típus: korai embrionális. standard. emlős (soksejtű) M. G2. G1.")
6
Kinázok és foszfatázok működése
A foszforilálás aktiválja vagy inaktiválja a fehérjéket = szabályozza a működésüket Egy fehérjének több különböző hatású foszforilációs helye lehet. Poszttranszlációs szintű szabályozás.

7
Ubiquitin – függő proteolízis
aktiváló konjugáló ligáló enzim (specifikus)
")
8
Cdk szabályozása proteolízis proteolízis kinase foszfatáz kinase

9
A fő ellenőrzési pontok
G1, G2, M Is all DNA intact? A sejt akkor lépi át az ellenőrzési pontot, ha az azt megelőző folyamatok hibátlanul megtörténtek

10
2001 Orvosi Nobel díj Leland Hartwell Paul Nurse Tim Hunt
ellenőrzési pontok Cdk ciklin

11
Standard sejtciklus M-fázis egysejtűek (élesztő) M reguláló rendszer
Cdk + G1 ciklin SPF vagy Start kináz Cdk + M ciklin MPF 3 ellenőrzési pont M G2 G1 Az első Cdk = cdc2 vagy p34

12
MPF Cdk SPF Cdk szabályozása a standard sejtciklusban M-fázis G2 G1 S
M ciklin M-fázis MPF G2 Cdk M ciklin G1 G1 ciklin SPF S G1 ciklin - szintézis - degradation

13
M-fázis promoting factor aktiváció
ciklin Aktiváló kináz MPFi MPFi Cdk Gátló kináz foszfatáz MPFa

14
A standard sejtciklus szabályozása
Gátló kináz foszfatáz proteolízis Szubsztrát-PO4 + Ciklin(ek) Cdk MPF szubsztrát transzláció transzláció G1 és M ciklinek lebontása MPF aktivációja (aktiváló kináz és foszfatáz) és gátlása (gátló kináz) Aktiváló kináz
 Cdk. MPF. szubsztrát. transzláció. transzláció. G1 és M ciklinek lebontása. MPF aktivációja (aktiváló kináz és foszfatáz) és gátlása (gátló kináz) Aktiváló kináz.")
15
Soksejtűek sejtciklusa

16
A sejtciklus eltérő szerepe egy- és soksejtűekben
A sejtek addig osztódnak, amíg valamely kedvezőtlen környezeti körülmény le nem állítja (negatív) a sejtciklust. Felnőtt szervezetben a sejtek addig nem osztódnak, amíg a környezetükből stimulust (pozitív) nem kapnak.
 a sejtciklust. Felnőtt szervezetben a sejtek addig. nem osztódnak, amíg a környezetükből. stimulust (pozitív) nem kapnak.")
17
Soksejtűekben a sejtek sorsa jórészt a környezetüktől (más sejtektől függ)
Survival

18
Soksejtűek sejtciklusa
Go fázis növekedési faktor G1 fázis letapadási függőség G1 fázis denzitás függő sejtosztódás gátlás (kontakt gátlás) (kevés növekedési faktor) M-fázis M G1 G2 Go G2 Restrikciós pont -Növekedési faktor -letapadás S
 (kevés növekedési faktor) M-fázis. M. G1. G2. Go. G2. Restrikciós pont. -Növekedési. faktor. -letapadás. S.")
19
A soksejtűek sejtciklusát
számos Cdk és ciklin szabályozza

20
Soksejtűek Cdk-i és ciklinjei
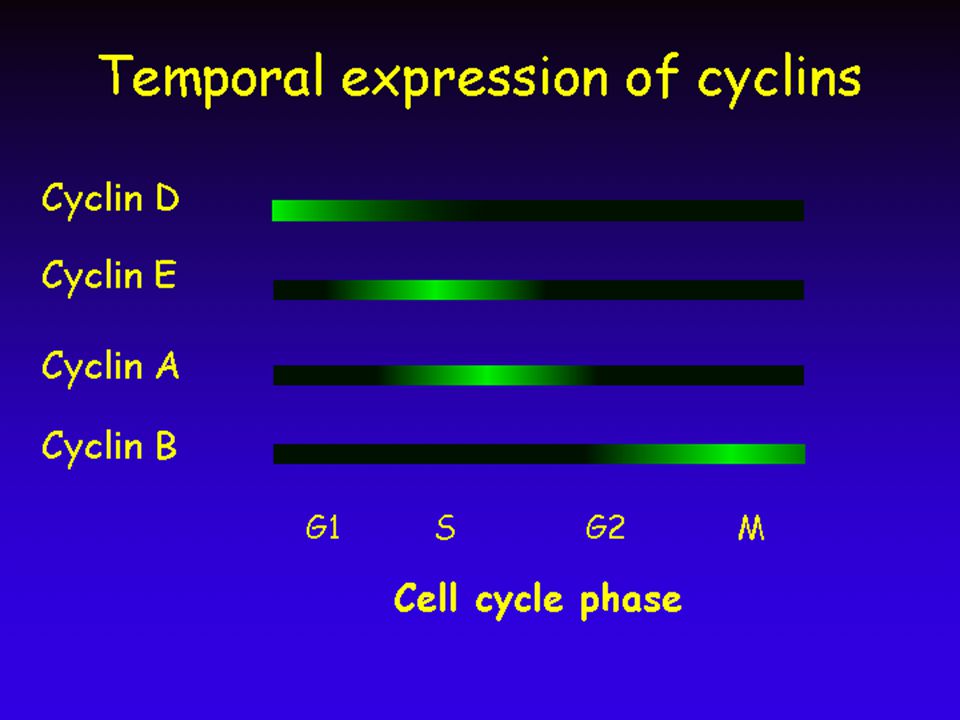
Cdk – ciklin komplex ciklin Cdk G1-Cdk Ciklin D (G1 ciklin) Cdk 4, 6 G1/S-Cdk Ciklin E (G1/S ciklin) Cdk 2 S-Cdk (SPF) Ciklin A (S ciklin) M-Cdk (MPF) Ciklin B (M ciklin) Cdk 1
 Cdk 4, 6. G1/S-Cdk. Ciklin E. (G1/S ciklin) Cdk 2. S-Cdk. (SPF) Ciklin A. (S ciklin) M-Cdk. (MPF) Ciklin B. (M ciklin) Cdk 1.")
21
Növekedési faktorok Go G1 átmenethez szükségesek
fehérjék ( 50); enzim kapcsolt receptoruk van parakrin szekréció nagyon specifikusak; esetleg kombinációban hatásosak hatásaik koncentráció függőek a pontos hatásuktól függően csoportjaik: mitogének, növekedési faktorok, túlélési faktorok pl. EGF, FGF, PDGF, NGF stb.
; enzim kapcsolt receptoruk van. parakrin szekréció. nagyon specifikusak; esetleg kombinációban. hatásosak. hatásaik koncentráció függőek. a pontos hatásuktól függően csoportjaik: mitogének, növekedési faktorok, túlélési faktorok. pl. EGF, FGF, PDGF, NGF stb.")
22
A növekedési faktorok hatásmechanizmusa
1. Aktivált tirozinkináz kaszkád Aktivált Ras (monomer G protein) Aktivált szerin/treonin kináz kaszkád (MAP-ok=mitogén aktivált proteinek) 2. Transzkripciós faktorok 3. Korai válaszgének transzkripciója (c-jun,c-myc,c-fos) 4. Jun,Myc,Fos transzkripciós faktorok szintézise 5. Késői válaszgének transzkripciója 6. Cdk-k és Dc (és sok egyéb) szintézise 7. Visszalépés G1-be
 Aktivált szerin/treonin kináz kaszkád (MAP-ok=mitogén aktivált. proteinek) 2. Transzkripciós faktorok. 3. Korai válaszgének. transzkripciója. (c-jun,c-myc,c-fos) 4. Jun,Myc,Fos transzkripciós. faktorok szintézise. 5. Késői válaszgének. transzkripciója. 6. Cdk-k és Dc (és sok egyéb) szintézise. 7. Visszalépés G1-be.")
23
A letapadás is lehet sejtciklus indukáló
Autofoszforilációs hely Extracelluláris mátrix Integrin Plazmamembrán Kináz domén FAK MAP kináz Src Sos Grb FAK = fokális adhéziós kináz

24
A Cdk inhibitorok két családja
INK4 család – p14, 16, 18, 19 – csak a Cdk4/6 –ot (kapcsolódást a ciklinnel és az aktivitást) gátolja CIP/KIP család – p21, 27, 57 – elsősorban a Cdk2-E/A aktivitását gátolják
 gátolja. CIP/KIP család – p21, 27, 57 – elsősorban a Cdk2-E/A aktivitását gátolják.")
25
Visszalépés a G1 fázisba
M G1 Restrikciós pont S Stimuláció Gátlás pRb-PO4 Cdk2 - Ec Cdk4/6- Dc Dc pRb-E2F p107,130 p21,27,57 Go p14,15,16,19 E2F p53 Ac Ec Növekedési faktor letapadás DNS sérülés Cdk2- Ac

26
Myc transzkripciós faktor
target génjei: Cyclin D SCF=ubiquitin ligáz E2F

28
S fázis

29
DNS szintézis iniciálása I.
ORC az origókhoz (sok ezer) kapcsolódik Más fehérjék bekötődése Pre-RC
 kapcsolódik. Más fehérjék. bekötődése. Pre-RC.")
30
DNS szintézis iniciáció II.
Cdk2-Ac aktivitás A DNS replikálódik, de csak egyszer (Cdk2-Ac) 1.A DNS replikáció iniciálódik 2. Az új iniciáció gátolt (ORC defoszforilálódás a mitózis után)
 1.A DNS replikáció iniciálódik. 2. Az új iniciáció gátolt. (ORC defoszforilálódás. a mitózis után)")
31
S fázis iniciáció Kromatin remodelling
Sok origó (replikáció start pontja) hosszú DNS ORC (origo felismerő komplex) bekötődése Cdc6 bekötődése Egyéb fehérjék bekötődése pre-RC SCF (ubiqutin ligáz) p27 degradáció Cdk2-Ec aktiváció Cdk2-Ec aktiválja a Cdk2-Ac-t Cdk2-Ac foszforilálja a pre-RC elemeit (pl. Cdc6-ot) DNS replikáció (de csak egyszer !)
 hosszú DNS. ORC (origo felismerő komplex) bekötődése. Cdc6 bekötődése. Egyéb fehérjék bekötődése pre-RC. SCF (ubiqutin ligáz) p27 degradáció Cdk2-Ec aktiváció. Cdk2-Ec aktiválja a Cdk2-Ac-t. Cdk2-Ac foszforilálja a pre-RC elemeit (pl. Cdc6-ot) DNS replikáció. (de csak egyszer !)")
32
DNS szintézis (egy replikációs origó)
5’ 3’ 3’ 1. 2. 3. 5’ 5’ 3’ 3’ 5’ RNS primer

33
S fázis (DNS szintézis)
2 replikációs villa (replikációs buborék) primáz RNS primer DNA polimeráz 5’ 3’ monomerek: dezoxinukleozid trifoszfátok leading (vezető) és lagging (elmaradó) szál primer kivágása ligáz szemi-konzervatív Azonos DNS molekulák
 primáz RNS primer. DNA polimeráz 5’ 3’ monomerek: dezoxinukleozid trifoszfátok. leading (vezető) és lagging (elmaradó) szál. primer kivágása. ligáz. szemi-konzervatív. Azonos DNS molekulák.")
34
A hisztonkorongok elrendeződésének két modellje
=

35
End replication problem és a megoldás = telomeráz

37
Protoonkogének és tumor szupresszor gének termékei
A sejtciklust serkentik gátolják Cdk-k, ciklinek, pRb, CKI-k növekedési faktorok stb. (ras gyakori) (p21, p53 stb) Tumor szupresszor protoonkogén
 (p21, p53 stb) Tumor szupresszor. protoonkogén.")
38
A sejtciklust szabályozó gének szerepe a tumorok kialakulásában
protoonkogének Tumor szupresszor

39
A p53 központi reguláló szerepe

40
A p53 szabályozása és működése
DNA damage induces phosphorylation (P) of p53 (at the Mdm2 binding site) and Mdm2, preventing Mdm2 from binding to p53. As a result, the p53 level increases, and stops cells from entering cell cycle until the DNA is repaired. If repair fails, p53 initiates apoptosis (programmed cell death). Mechanisms resulting in a decrease in p53 steady-state levels are indicated with green arrows and those resulting in increases p53 levels are indicated with red arrows. Solid lines indicate active mechanisms and broken lines indicate inactivated mechanisms. Ubiquitin ligáz
 of p53 (at the Mdm2 binding site) and Mdm2, preventing Mdm2 from binding to p53. As a result, the p53 level increases, and stops cells from entering cell cycle until the DNA is repaired. If repair fails, p53 initiates apoptosis (programmed cell death). Mechanisms resulting in a decrease in p53 steady-state levels are indicated with green arrows and those resulting in increases p53 levels are indicated with red arrows. Solid lines indicate active mechanisms and broken lines indicate inactivated mechanisms. Ubiquitin ligáz.")
41
Protoonkogének és tumor szupresszor gének a humán kromoszómákon

42
A sejtciklust szabályozó gének konzervatívak, univerzálisak

43
Fehérjék csoportjai, amelyek a sejtek növekedését szabályozzák. Mutációjuk oka lehet a tumorok kialakulásának.

44
Knudson hipotézise a tumor szupresszor gének szerepéről a tumorok kialakulásában
Két találat teória, LOH = loss of heterozygosity

45
Retinoblasztoma Kisgyermekkorban a retinában kialakuló tumor
Oka az RB1 gén csíravonalbeli mutációja Kb. fele öröklődő és a fele sporadikus

46
Li-Fraumeni szindróma
Oka a p53 mutáció Öröklődő – fiatalkori tumorok (emlő, agy, sarcoma, akut leukémia)
")
47
A tumorok jellemző tulajdonságai

48
A vastagbél rák kialakulásához is sok mutáció vezet
Adenomatous polyposis coli

49
Wnt fehérjék – konzervatív morfogének, sejt differenciálódás, polaritás, embrionális fejlődés,
tumorok

50
Hepatocarcinoma kialakulása

51
Centroszóma ciklus

52
A centroszóma (MTOC) és a centriolumok szerkezete
mikrotubulusok centriolumok tubulin gyűrűk

53
Centroszóma ciklus M Cdk1+Bc (MPF) Cdk2+Ec G2 G1 eltávolodás szétválás
Cdk2+Ac S duplikálódás

54
Tumorokban gyakori az atípusos mitózis (tripoláris)
Három centroszóma

55
M fázis

56
Nagy változások az M fázisban
DNA (blue), microtubules (red) nuclear envelope marker (green) Nagy változások az M fázisban Interphase metaphase anaphase telophase
, microtubules (red) nuclear envelope marker. (green) Nagy változások az M fázisban. Interphase metaphase. anaphase. telophase.")
57
M-fázis promoting faktor aktiváció
ciklin Aktiváló kináz MPFi MPFi Cdk Gátló kináz foszfatáz MPFa

58
Néhány MPF szubsztrát nukleáris lamina laminjai magmembrán szétesése
Golgi membránban GM130 (COPI borított vezikulum dokkoló fehérje) Golgi is vezikulumokra esik szét kondenzin kromoszóma kondenzáció (H3 hisztont egy másik kináz foszforilálja) MAP-ok mitotikus orsó kialakulása foszfatáz (+ feedback) APC (anaphase promoting complex) aktiváció miozin inaktiváció
 Golgi is vezikulumokra esik szét. kondenzin kromoszóma kondenzáció (H3 hisztont. egy másik kináz foszforilálja) MAP-ok mitotikus orsó kialakulása. foszfatáz (+ feedback) APC (anaphase promoting complex) aktiváció. miozin inaktiváció.")
59
A mitózis fázisai Interfázis korai profázis késői profázis prometafázis Metafázis korai anafázis késői anafázis késői telofázis A metafázisban működik az M ellenőrzési pont.

60
Magmembrán szétesése MPF Laminok foszforilációja Interfázis Profázis
Foszforilált A és C lamin Foszforilált B lamin a magmembrán vezikulumokon

61
DNS-től a kromoszómáig
A DNS kondenzációját a kondenzin foszforilációja indítja el. telomérák

62
Kohezin Kondenzin Cornelia de Lange szindróma

63
Kohesin és kondenzin Smc = structural maintance
of chromosome + egyéb proteinek

64
A centroméra és a kinetokor nem ugyanaz

65
szerkezete és kimutatása
A kinetokor régió szerkezete és kimutatása

66
Scleroderma Autoimmun betegség – kinetochor elleni autoantitestek

67
Mitotikus orsó (metafázis)
asztrális kinetokor poláris mikrotubulusok polar

68
A mitózis fázisai Interfázis korai profázis késői profázis prometafázis Metafázis korai anafázis késői anafázis késői telofázis Ha metafázisban (M ellenőrzési pont) minden kromatidához megfelelő módon kapcsolódnak kinetochor mikrotubulusok, a sejt folytatja a mitózist…
 minden kromatidához megfelelő módon. kapcsolódnak kinetochor mikrotubulusok, a sejt folytatja a mitózist…")
69
Anafázis promoting komplex (APC)
APC = ubiquitin ligáz poliubiquitinált fehérjék proteaszóma MPFa (M fázis elején) APC (anafázis elején) szekurin lebomlás B ciklin lebomlás szeparáz MPFi ? kohezin MPF szubsztrátok defoszforilálása Anafázis, citokinézis
 APC (anafázis elején) szekurin lebomlás B ciklin lebomlás. szeparáz MPFi. kohezin MPF szubsztrátok. defoszforilálása. Anafázis, citokinézis.")
70
APC hatása kohesin leválása testvér kromatidák szétválása (anafázis)
proteasome kohesin leválása testvér kromatidák szétválása (anafázis)
")
71
Anafázis (Kinetochor MT depolimerizáció – no ATP)
(Poláris MT polimerizáció és sliding)
")
72
Miozin II változása a sejtciklusban

73
Citokinézis (aktin és miozin II)
Cytokinesis signals the end of 1 round of the cell cycle and the generation of 2 daughter cells. -It begins shortly after the onset of anaphase and is triggered by the inactivation of cdk1. -In Animal cells, cytokinesis is mediated by a contractile ring of actin and myosin II filaments that forms beneath the plasma membrane. -The location of the ring is detemined by the location of the mitotic spindle. Cleavage proceeds as contraction of the actin and myosin filaments pulls the PM inward. This eventually results in pinching the cell in half causing the production of 2 daughter cells

74
Citokinézis – kontraktilis gyűrű
aktin miozin

75
Citokinézis (SEM,TEM)
")
76
Regulációs stratégiák az emlős sejtciklusban
Gátló kináz foszfatáz proteolízis Szubsztrát-PO4 CKI-k + ciklin Cdk CKI-k szubsztrát proteolízis transzkripció Aktiváló kináz transzláció

77
Hogyan működnek az ellenőrzési pontok?
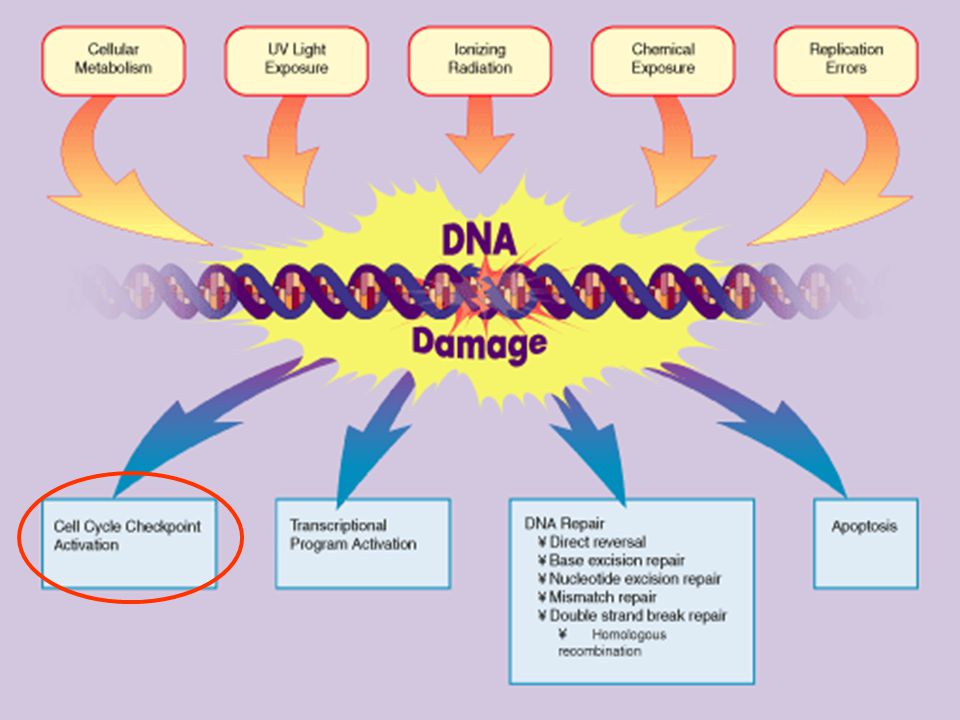
G G M DNS sérülés szabad kinetokor nem teljes replikáció érzékelő protein kinázok közvetítő effektor s e j t c i k l u s m e g á l l í t á s a

79
G1 ellenőrzési pont A DNS sérülés érzékelése p53 foszforiláció
Protein kináz p53 foszforiláció közvetítő effektor p21 transzkripció -In addition to CHK1 and CHK2, in mammalian cells G1 arrest is also mediated by a transcription factor and tumor suppressor gene called p53. -p53 is phosphorylated by both ATM and CHK2. In the case of the p53 protein, phosphorylation serves to stabilize a protein that would otherwise be rapidly degraded. -This stabilization, allows levels of p53 to increase so that p53 can activate transcription of the gene that encodes the Cdk inhibitor, p21. Activation of p21 leads to inhibition of Cdk2/cyclinE complexes. -If you recall from a few slides back, we said the Cdk2/cyclin E are necessary for progression from G1 to S. So as a result this interaction causes cell cycle arrest at the G1 checkpoint. -Another interesting note, it that p53 is the most commonly mutated protein in human cancers. Mutations that result in the loss of p53 function prevents G1 arrest in response to DNA damage. This allows damaged DNA to divide without being repaired. The inheritance of damaged DNA results in an increased frequency of mutations, general instability of the genome of those cells and tends to contribute to cancer development. Cdk2-Ec gátlás No S fázis

80
Ataxia telangiectasia (ATM)
Rendkívül sokféle, sok szervet érintő tünet együttes sugár érzékenység és tumorok kialakulása

81
G2 ellenőrzési pont ATR Protein kináz érzékelő közvetítő effektor

82
BRCA (breast cancer) szerepe a DNS sérüléseinek a kijavításában
Mutációjuk – emlő és petefészek tumorokban

83
M-ellenőrzési pont Van szabad kinetochor
Minden kinetochorhoz kapcsolódik MT APC Cdc20 APC

85
Egyéb kinázok a sejtciklus
szabályozásában

86
Polo szerű kinázok (Plk1) szerepe az M-fázisban
Plk1 lokalizáció Plk inhibitorok – (BI2536, GSK461364, ON-01910, HMN-214) – I-II fázisú klinikai kipróbálás
 – I-II fázisú klinikai kipróbálás.")
87
Aurora kináz A sötét kék, Aurora kináz B narancs



 DNS-ből,>")

 daganatkeltő.>")
 megváltozHAT Ebből visszakövetkeztethetünk a mutációt szenvedett gén funkciójára.>")












>")


 „késői” gének (late - L) 4.7 kb SV40 genom - kicsiny „tanulóvírus” fertőzést követően először.>")