Előadást letölteni
1
jelátvitel az immunrendszerben
Túl hosszú, utolsó dia lemaradt. Src aktiválás nem tudom kell-e

3
Ras Raf Mek1

4
A Fas ligand transzkripciójának szabályozása

7
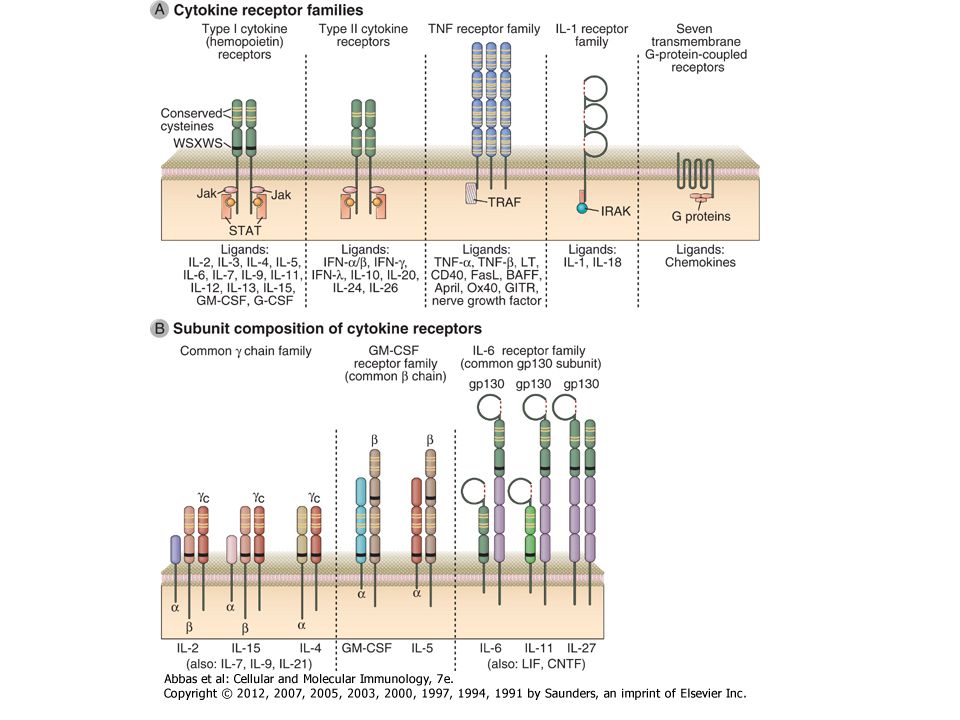
Receptor típusok az immunrendszerben
D vitamin Glucocorticoid PPARy BCR TCR FcRs Integrinek Citokinek Kemokin integrin Leukotrien Prosztaglandin C3a, C5a hisztamin EGFR InsulinR

8
Aktivitás Lokalizáció Interakció Posztranszlációs módosítások: Szerin/treonin foszforiláció Tirozin foszforiláció Ubiquitináció Palmitoiláció… Acetiláció Metiláció Hasítás .

9
Adaptor fehérjék

13
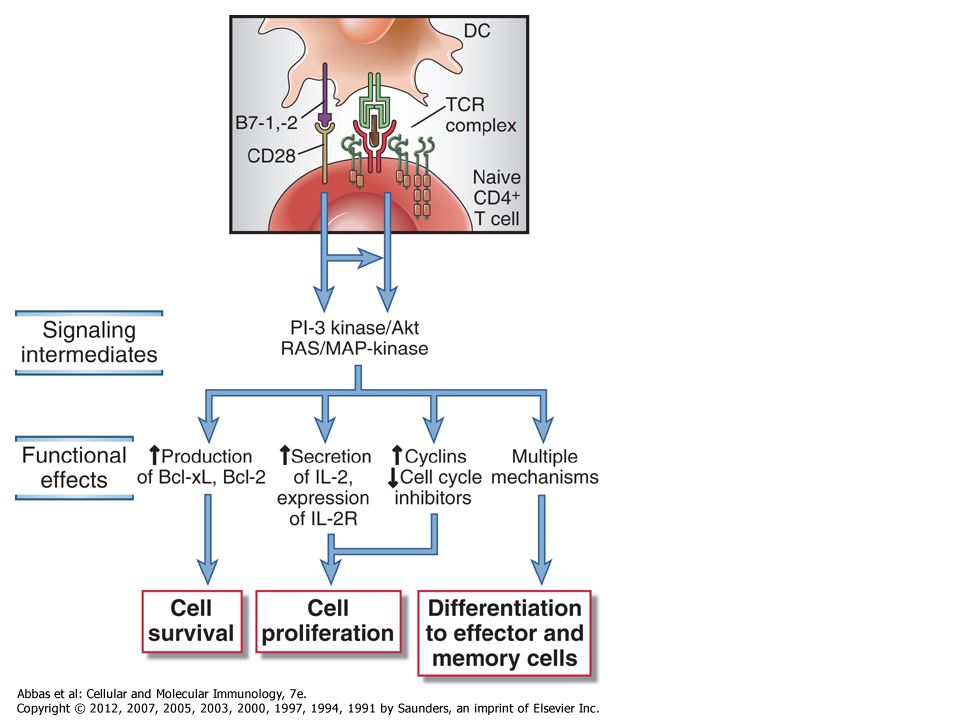
Aktív/nyugvó Elindulás/ ne induljon el Ligand mennyiség-jel intenzitás

14
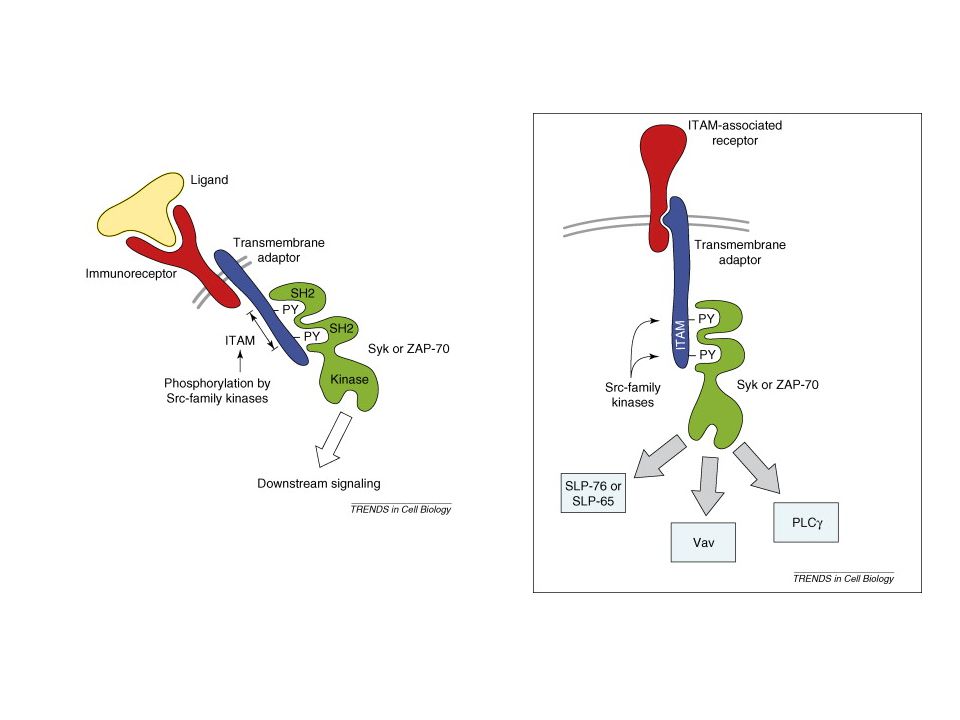
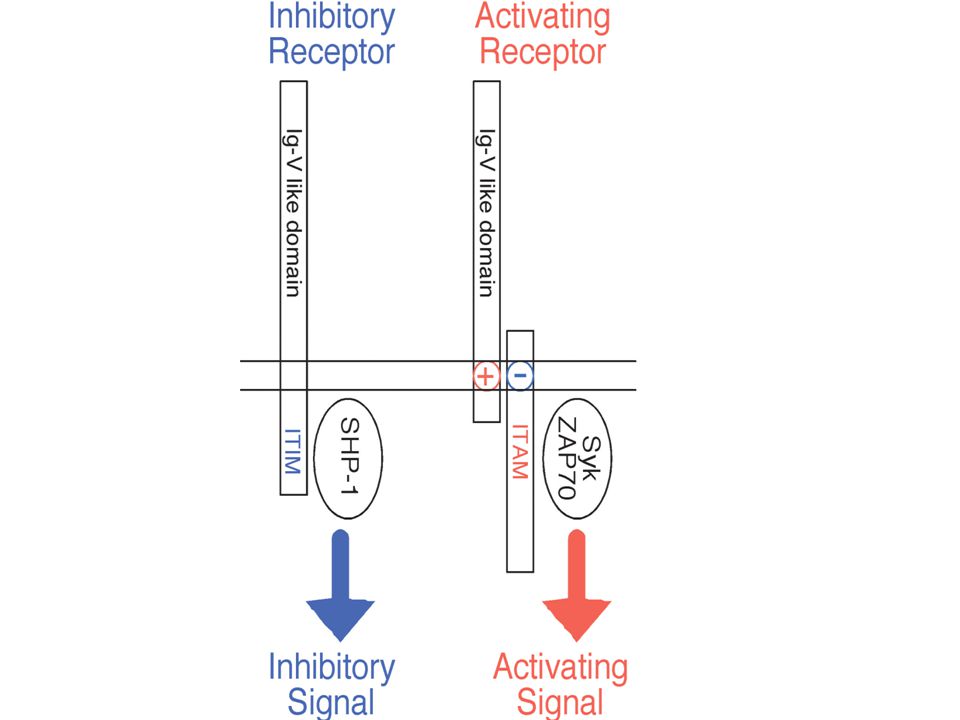
ITAM-ITIM motivum ITAMs (immunoreceptor tyrosine-based activating motifs) are found on receptors involved in cell activation and have the sequence YxxL/I(x)6-8YxxL/I, where Y represents a tyrosine residue, L represents leucine, I represents isoleucine, and x refers to any amino acid. ITIM (immunoreceptor tyrosine-based inhibitory motif), which has the consensus sequence V/L/IxYxxL, where V refers to valine
 are found on receptors involved in cell activation and have the sequence YxxL/I(x)6-8YxxL/I, where Y represents a tyrosine residue, L represents leucine, I represents isoleucine, and x refers to any amino acid. ITIM (immunoreceptor tyrosine-based inhibitory motif), which has the consensus sequence V/L/IxYxxL, where V refers to valine.")
16
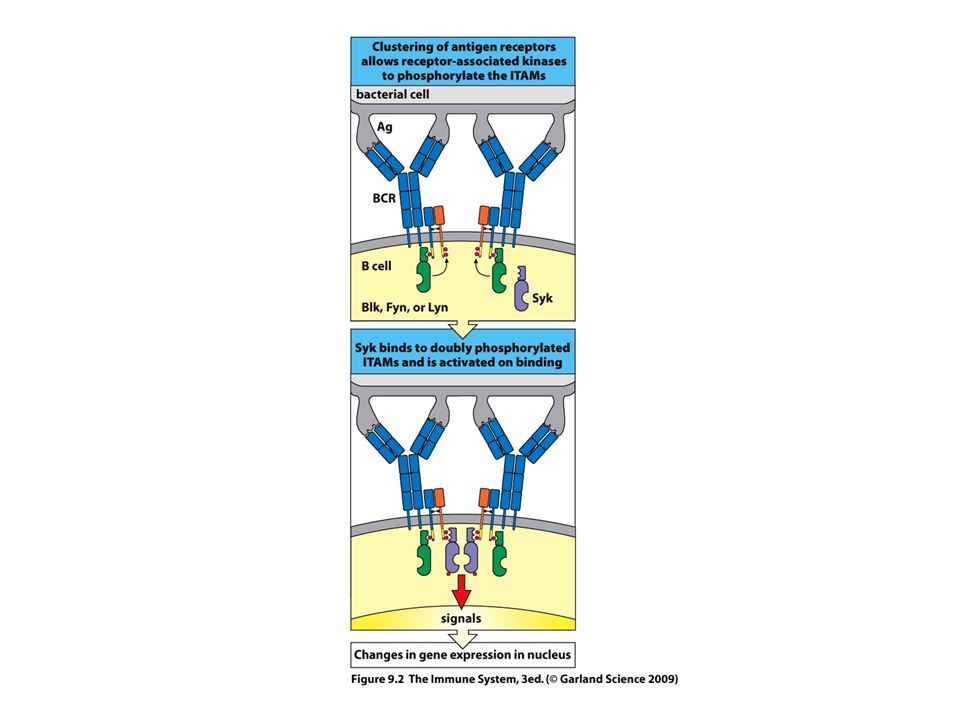
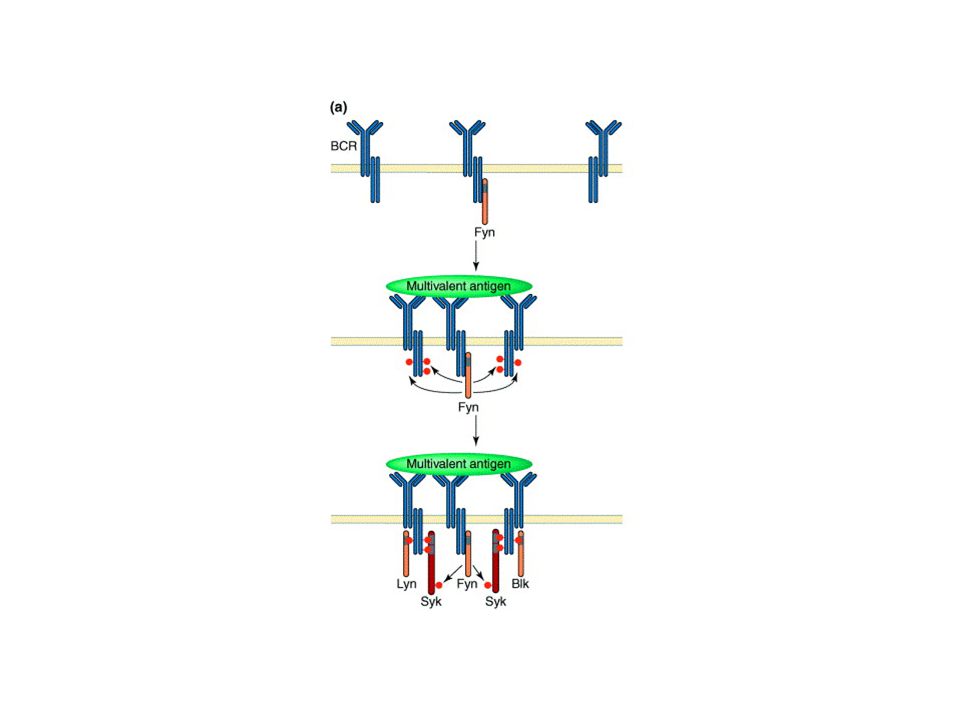
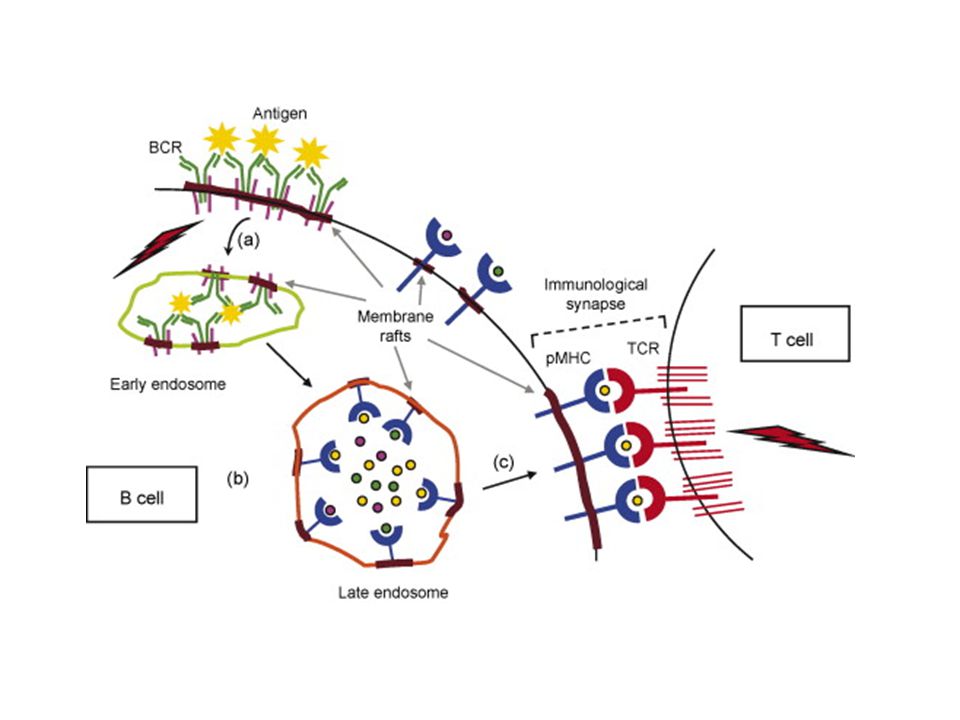
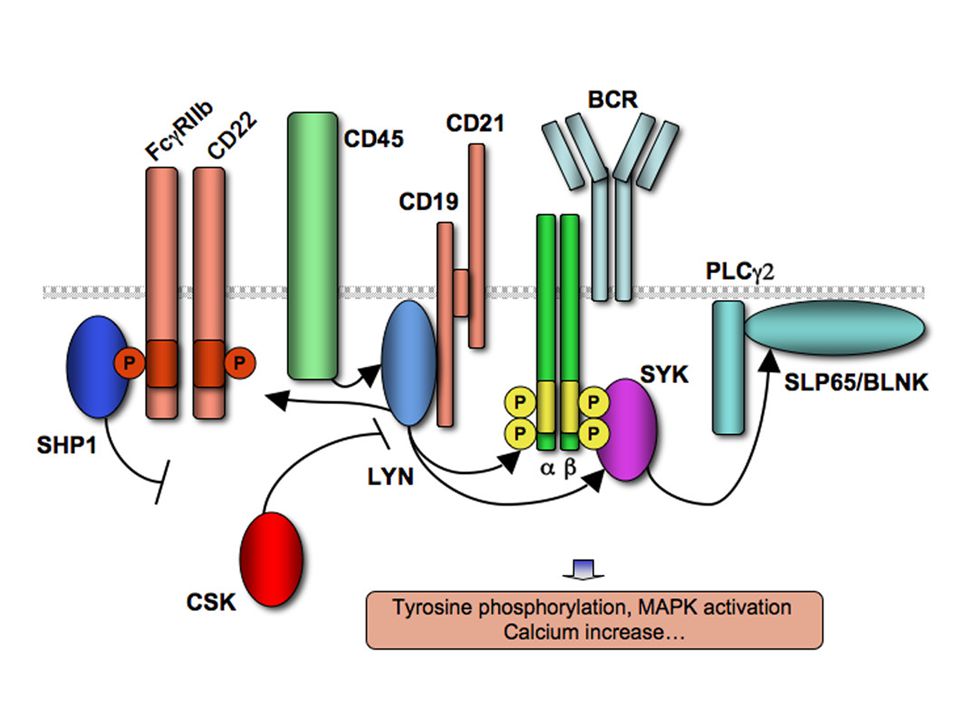
BCR-Igα/Igβ komplex The same signaling pathways are used by membrane IgM and IgD on naive B cells and by IgG, IgA, and IgE on B cells that have undergone isotype switching because all these membrane isotypes associate with Igα and Igβ.

18
BCR-TCR aktiválódás --ligand kötődés --receptor aggregáció (?)
--src-k foszforilálják az ITAM motívumot --syk/Zap70 kötődik a duplán foszforilált ITAM-hez --syk/Zap70 aktiváció—downsteram lépések

19
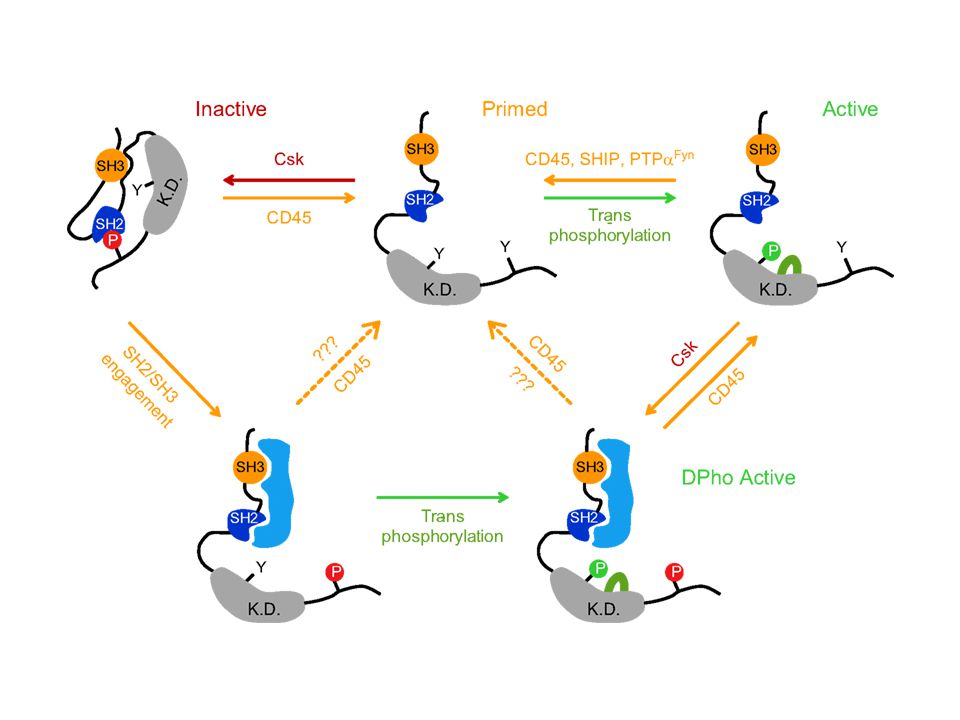
Jel első? lépése az ITAM foszforilációja az src kinázok által

21
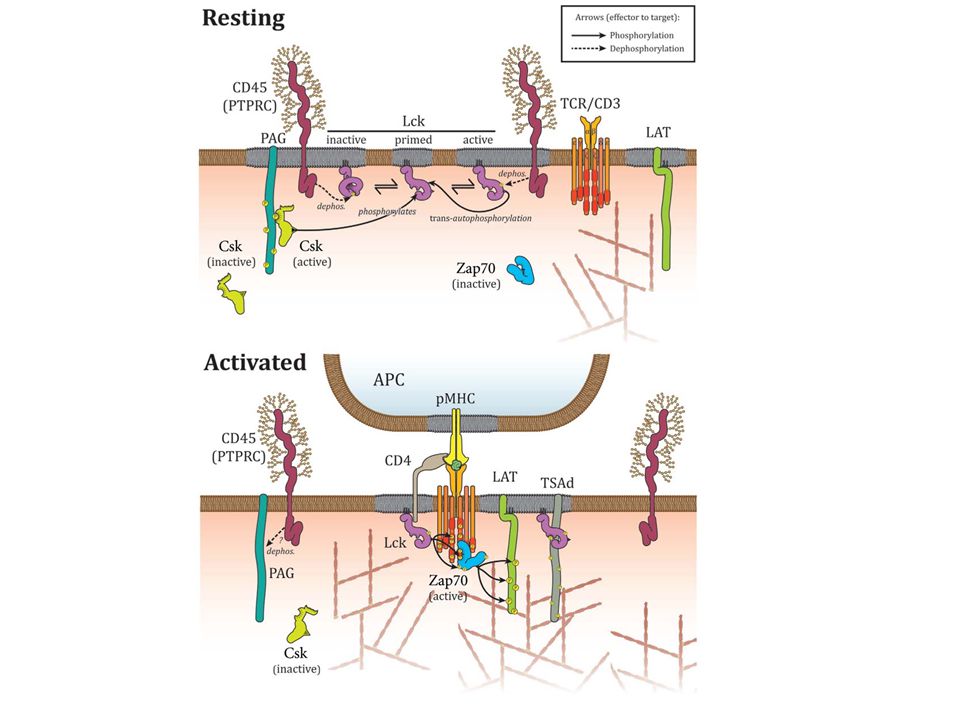
Jel első? lépése az src defoszforilációja
Jel első lépése a PAG (Csk kötő fehérje) defoszforilációja
 defoszforilációja.")
23
Syk autofoszforilálódik+ src is foszfoilálja

24
T Cell Receptor (TCR) Immunoglobulin (Ig) Components α and β chains Heavy and light chains Number of Ig domains One V domain and one C domain in each chain Heavy chain: one V domain, three or four C domains Light chain: one V domain and one C domain Number of CDRs Three in each chain for antigen binding Three in each chain Associated signaling molecules CD3 and ζ Igα and Igβ Affinity for antigen (Kd) M M (secreted Ig) Changes after cellular activation Production of secreted form No Yes Isotope switching Somatic mutations
 M M (secreted Ig) Changes after cellular activation. Production of secreted form. No. Yes. Isotope switching. Somatic mutations.")
25
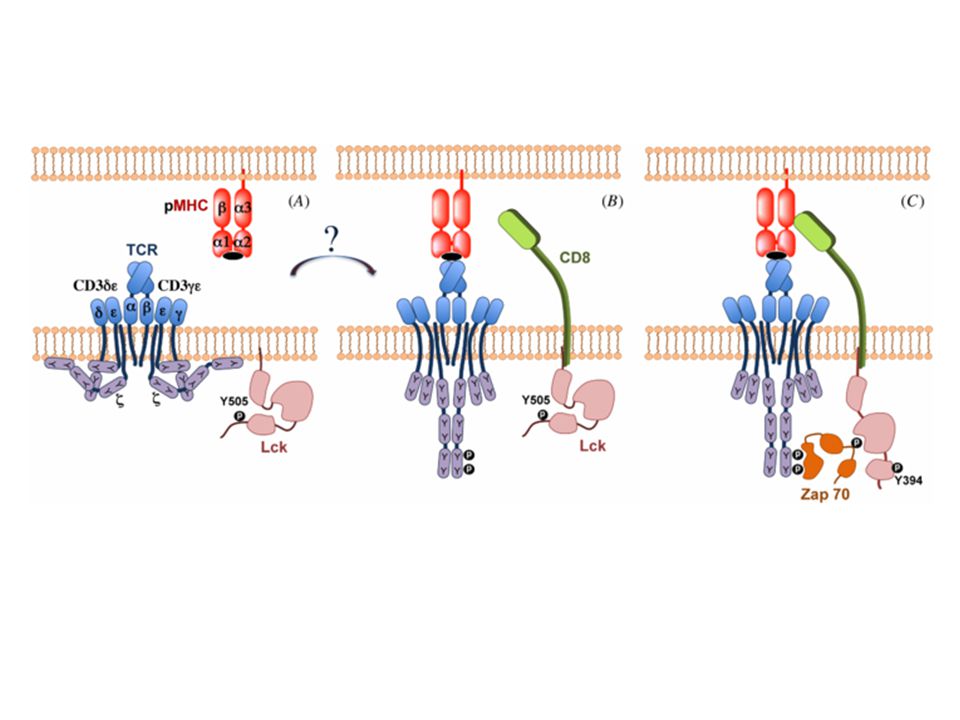
TCR-CD3 komplex The CD3 γ, δ, and Ε proteins are homologous to each other. The N-terminal extracellular regions of the γ, δ, and Ε chains each contains a single Ig-like domain, and therefore these three proteins are members of the Ig superfamily. The transmembrane segments of all three CD3 chains contain a negatively charged aspartic acid residue that binds to positively charged residues in the transmembrane domains of the TCR α and β chains. Each TCR complex contains one TCR αβ heterodimer associated with one CD3 γΕ heterodimer, one CD3 δΕ heterodimer, and one disulfide-linked ζζ homodimer. The CD3 proteins are required not only for signaling in T cells but for surface expression of the functionally complete receptor complex on T cells. The ζ chain is also associated with signaling receptors on lymphocytes other than T cells, such as the Fcγ receptor (FcγRIII) of NK cells.
 of NK cells.")
26
T-sejt MHC MHC sok féle peptid– hogyan aggregálhat?? CD 4-8 koreceptor/szinapszis CD 4-8 nem polimorf MHC résszel asszociál A CD 4-8-hoz kapcsolódó Lck (src kináz) a CD3 ITAM motívumjainak közelébe kerül
 a CD3 ITAM motívumjainak közelébe kerül.")
29
Immunológia szinapszis SMAC—supramolecular activation complex
TCR, CD3,CD4-8, CD28 a központban, inegrinek szélen, egyes molekulák pl. foszfatázok ‘kiszorítva’ Megszabja a T-sejt irányát, granulum ürülés, CD40-CD40L interakció kialakulás.. Figure 7-13 The immunologic synapse. A, This figure shows two views of the immunologic synapse in a T cell-APC conjugate (shown as a Nomarski image in panel c). Talin, a protein that associates with the cytoplasmic tail of the LFA-1 integrin, was revealed by an antibody labeled with a green fluorescent dye, and PKC-, which associates with the TCR complex, was visualized by antibodies conjugated to a red fluorescent dye. In panels a and b, a two-dimensional optical section of the cell contact site along the x-y axis is shown, revealing the central location of PKC- and the peripheral location of talin, both in the T cell. In panels d-f, a three-dimensional view of the entire region of cell-cell contact along the x-z axis is provided. Note, again, the central location of PKC- and the peripheral accumulation of talin. (Reprinted with permission of Macmillan Publishers Ltd. from Monks CRF, BA Freiburg, H Kupfer, N Sciaky, and A Kupfer. Three dimensional segregation of supramolecular activation clusters in T cells. Nature 395:82-86, copyright 1998.) B, A schematic view of the synapse, showing talin and LFA-1 in the p-SMAC (green) and PKC- and the TCR in the c-SMAC (red).
. Talin, a protein that associates with the cytoplasmic tail of the LFA-1 integrin, was revealed by an antibody labeled with a green fluorescent dye, and PKC-, which associates with the TCR complex, was visualized by antibodies conjugated to a red fluorescent dye. In panels a and b, a two-dimensional optical section of the cell contact site along the x-y axis is shown, revealing the central location of PKC- and the peripheral location of talin, both in the T cell. In panels d-f, a three-dimensional view of the entire region of cell-cell contact along the x-z axis is provided. Note, again, the central location of PKC- and the peripheral accumulation of talin. (Reprinted with permission of Macmillan Publishers Ltd. from Monks CRF, BA Freiburg, H Kupfer, N Sciaky, and A Kupfer. Three dimensional segregation of supramolecular activation clusters in T cells. Nature 395:82-86, copyright 1998.) B, A schematic view of the synapse, showing talin and LFA-1 in the p-SMAC (green) and PKC- and the TCR in the c-SMAC (red).")
33
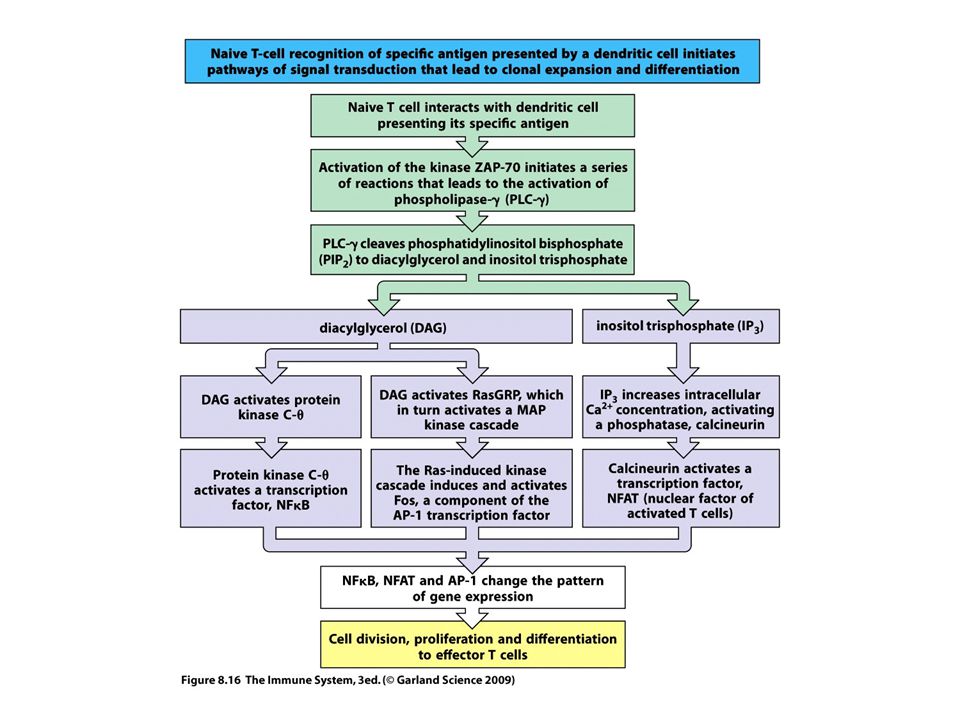
Jeltovábbítás

34
RAS MAPK Figure 7-14 The Ras-MAP kinase pathway in T cell activation. ZAP-70 that is activated by antigen recognition phosphorylates membrane-associated adaptor proteins (such as LAT), which then bind another adaptor, Grb-2, that provides a docking site for the GTP/GDP exchange factor SOS. SOS converts Ras·GDP to Ras·GTP. Ras·GTP activates a cascade of enzymes, which culminates in the activation of the MAP kinase ERK. A parallel Rac-dependent pathway generates another active MAP kinase, JNK (not shown).
, which then bind another adaptor, Grb-2, that provides a docking site for the GTP/GDP exchange factor SOS. SOS converts Ras·GDP to Ras·GTP. Ras·GTP activates a cascade of enzymes, which culminates in the activation of the MAP kinase ERK. A parallel Rac-dependent pathway generates another active MAP kinase, JNK (not shown).")
35
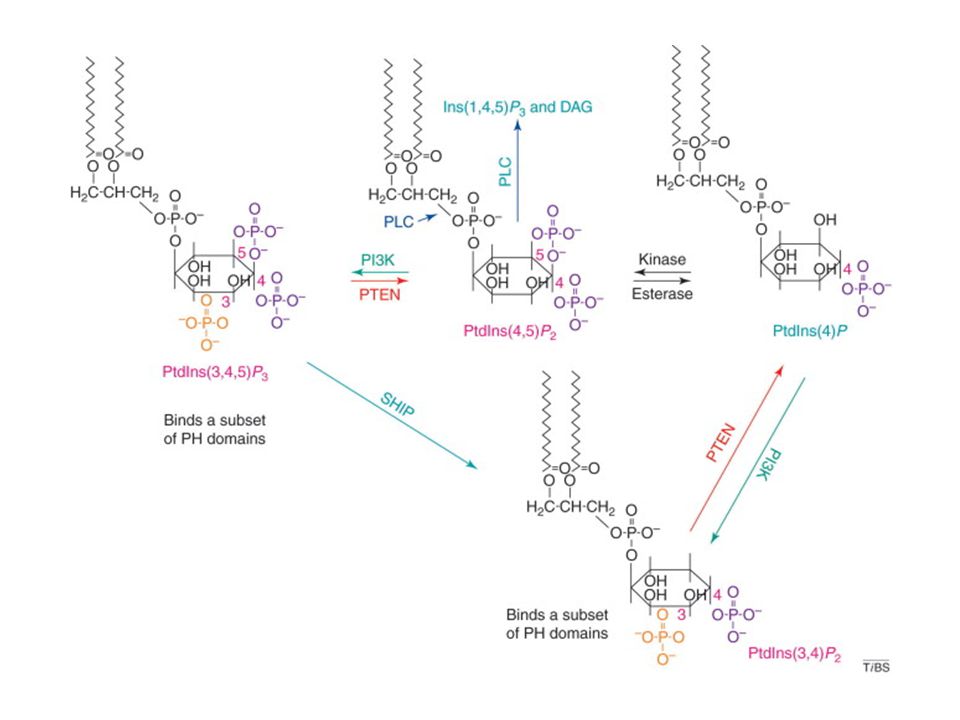
PLCγ PKC-Ca++ Figure 7-15 T cell signaling downstream of PLCγ1. A, The LAT adaptor protein that is phosphorylated on T cell activation binds the cytosolic enzyme PLCγ1, which is phosphorylated by ZAP-70 and other kinases, such as Itk, and activated. Active PLCγ1 hydrolyzes membrane PIP2 to generate IP3, which stimulates an increase in cytosolic calcium, and DAG, which activates the enzyme PKC. B, Depletion of endoplasmic reticulum calcium is sensed by STIM1. C, STIM1 which induces the opening of the CRAC channel that facilitates entry of extracellular calcium into the cytosol. Orai is a component of the CRAC channel. Increased cytosolic calcium and PKC then activate various transcription factors, leading to cellular responses.

38
Transzkripciós faktorok

40
PI3K

41
Koreceptorok: T-sejt, CD4, CD8, B-sejt CD19 (CR2-CD21) Kostimulátor molekulák:pl.CD28 (kostimulátor nem ismeri fel ugyanazt a komplexet mint az antigén receptor- szemeben a koreceptorral)
")
42
Figure 7-20 Role of complement in B cell activation
Figure 7-20 Role of complement in B cell activation. B cells express a complex of the CR2 complement receptor, CD19, and CD81. Microbial antigens that have bound the complement fragment C3d can simultaneously engage both the CR2 molecule and the membrane Ig on the surface of a B cell. This leads to the initiation of signaling cascades from both the BCR complex and the CR2 complex, because of which the response to C3d-antigen complexes is greatly enhanced compared with the response to antigen alone.

43
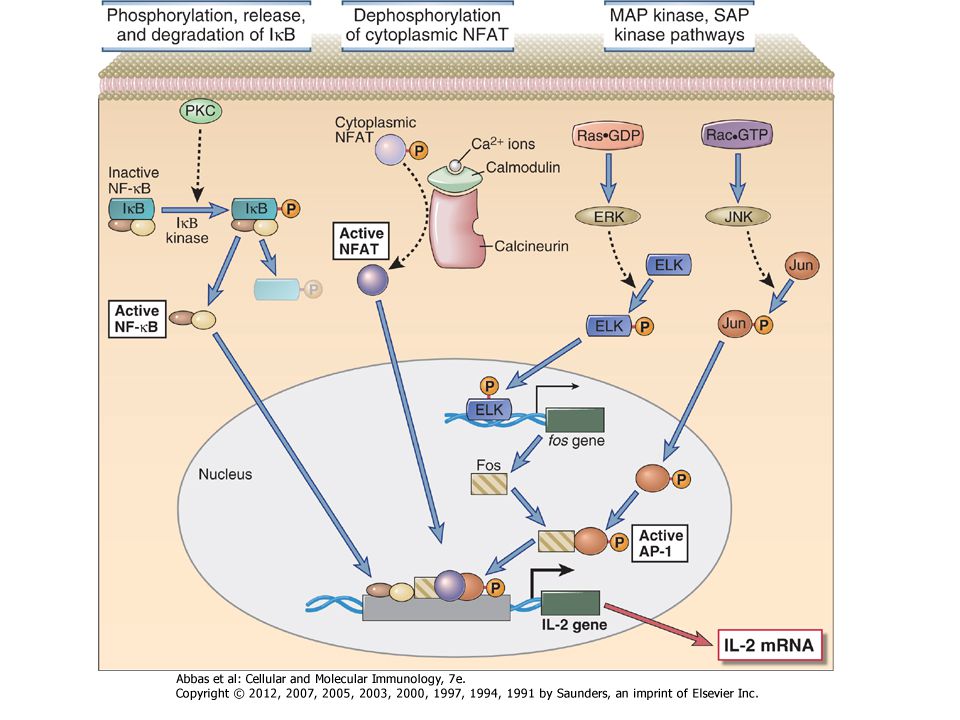
Figure 7-16 Activation of transcription factors in T cells
Figure 7-16 Activation of transcription factors in T cells. Multiple signaling pathways converge in antigen-stimulated T cells to generate transcription factors that stimulate expression of various genes (in this case, the IL-2 gene). The calcium-calmodulin pathway activates NFAT, and the Ras and Rac pathways generate the two components of AP-1. Less is known about the link between TCR signals and NF-κB activation. (NF-κB is shown as a complex of two subunits, which in T cells are typically the p50 and p65 proteins, named for their molecular sizes in kilodaltons.) PKC is important in T cell activation, and the PKC- isoform is particularly important in activating NF-κB. These transcription factors function coordinately to regulate gene expression. Note also that the various signaling pathways are shown as activating unique transcription factors, but there may be considerable overlap, and each pathway may play a role in the activation of multiple transcription factors.
. The calcium-calmodulin pathway activates NFAT, and the Ras and Rac pathways generate the two components of AP-1. Less is known about the link between TCR signals and NF-κB activation. (NF-κB is shown as a complex of two subunits, which in T cells are typically the p50 and p65 proteins, named for their molecular sizes in kilodaltons.) PKC is important in T cell activation, and the PKC- isoform is particularly important in activating NF-κB. These transcription factors function coordinately to regulate gene expression. Note also that the various signaling pathways are shown as activating unique transcription factors, but there may be considerable overlap, and each pathway may play a role in the activation of multiple transcription factors.")
46
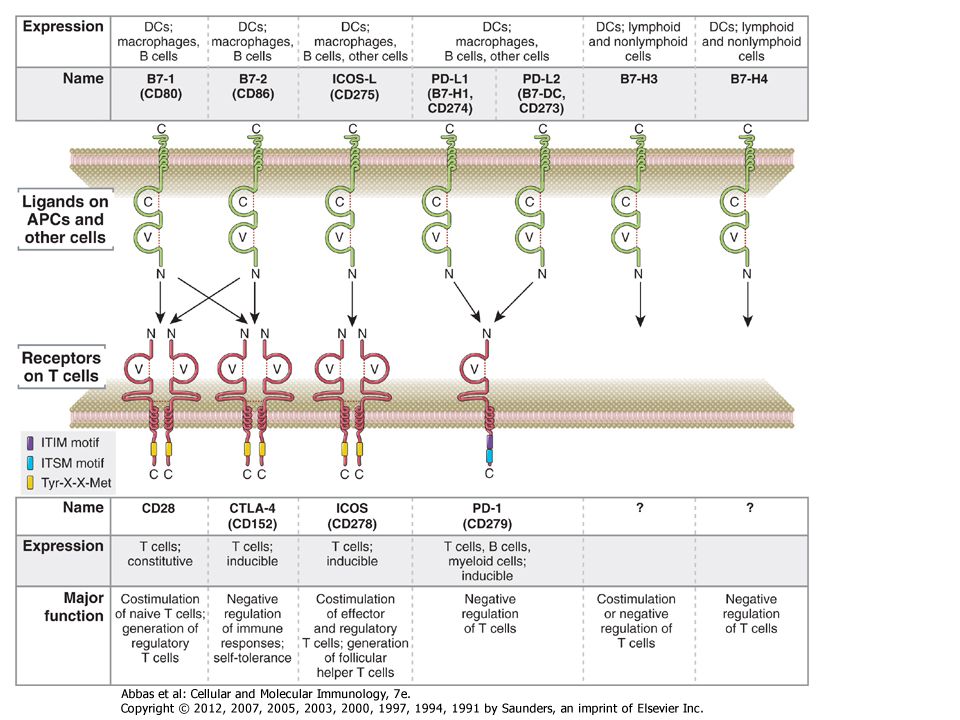
Jel gátlás Gátló receptorok: T-sejt :CTLA-4 ,PD-1 B-sejt: CD22, FcγRIIB, Ubiquitináció foszfatázok

48
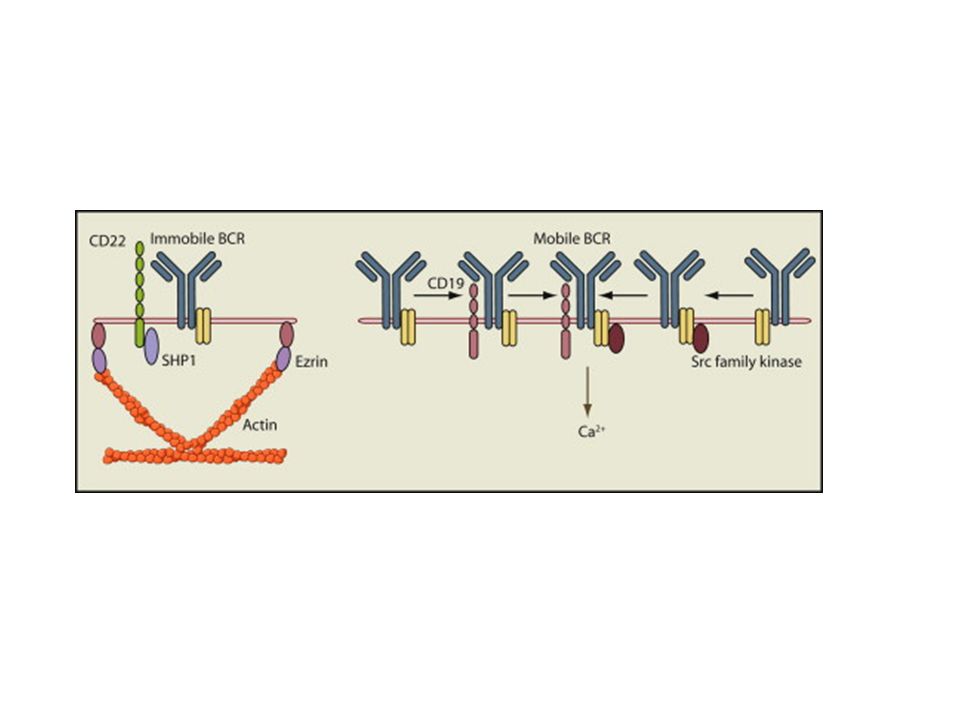
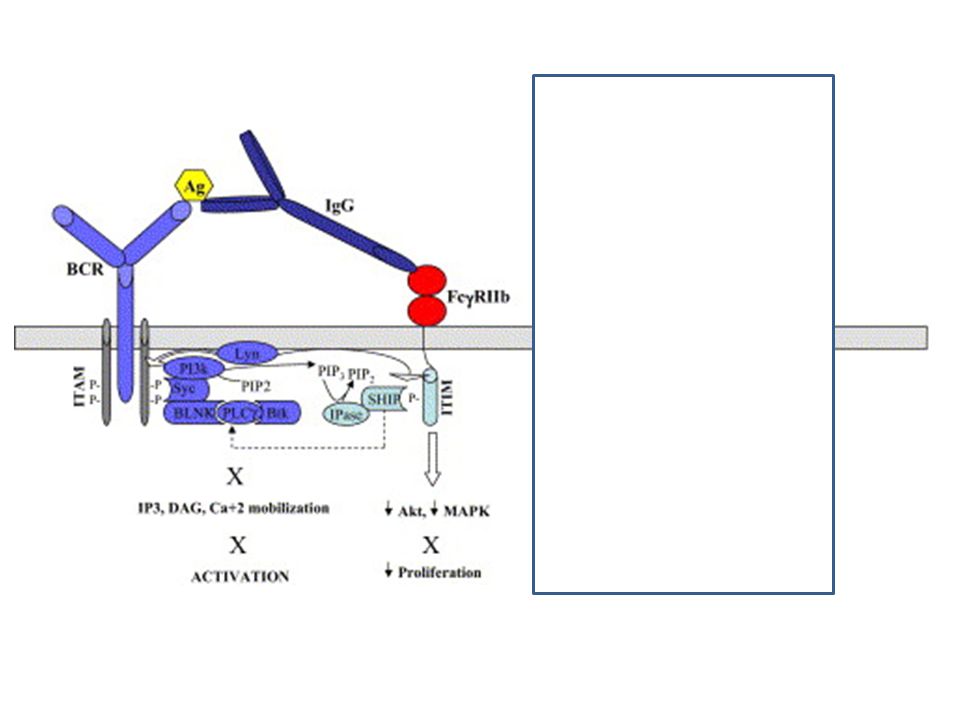
Az ITIM közvetített gátlás az src kinázok aktivációját igényli

52
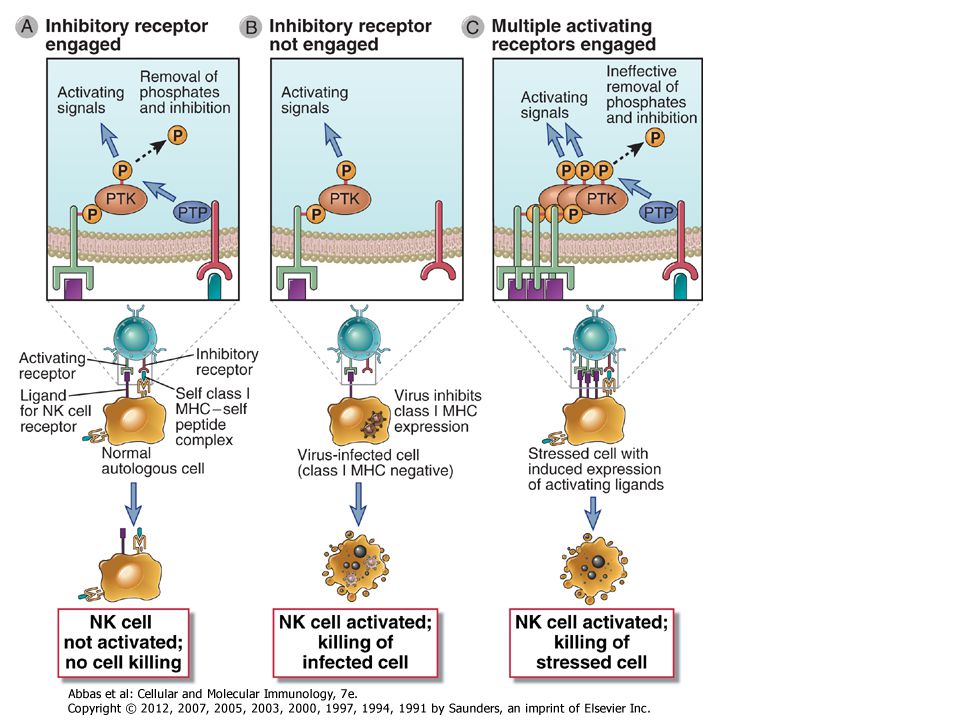
In human NK cells, the key inhibitory receptors can broadly be divided into three groups: KIRs or killer Ig-like receptors (see Chapter 4); ILT (Ig-like transcript) family proteins that are closely related to KIRs; and C-type lectins, the major one being a heterodimer consisting of the NKG2A C-type lectin and CD94. These inhibitory receptors are not restricted to NK cells and may also be present on some activated T cells. KIRs contain extracellular Ig domains that can recognize class I HLA molecules, and a subset of these receptors contains cytosolic ITIM motifs. ILT-2, part of an evolutionarily older family of inhibitory receptors than KIRs, also has extracellular Ig domains that bind HLA class I and cytosolic ITIM motifs. The CD94/NKG2A dimer binds to an atypical class I MHC molecule called HLA-E, and the NKG2A chain of this dimer contains cytosolic ITIM motifs.

54
Intracelluláris foszfatáz

55
Figure 7-22 Role of the ubiquitin ligase Cbl-b in terminating T cell responses. Cbl-b is recruited to the TCR complex, where it facilitates the monoubiquitination of CD3, ZAP-70, and other proteins of the TCR complex. These proteins are targeted for proteolytic degradation in lysosomes and other organelles (not shown). . Recruitment of Cbl-b to the TCR complex and associated adaptor proteins leads to the monoubiquitination, endocytosis, and lysosomal degradation of the TCR complex, and this may be a mechanism for the attenuation of TCR signaling (Fig. 7-22). CD28 signals block the inhibitory activity of Cbl-b, and this is one mechanism by which costimulation augments TCR signals. In knockout mice lacking Cbl-b, the T cells respond to antigen even without CD28-mediated costimulation and produce abnormally high amounts of IL-2. These mice develop autoimmunity as a result of the enhanced activation of their T cells.
. CD28 signals block the inhibitory activity of Cbl-b, and this is one mechanism by which costimulation augments TCR signals. In knockout mice lacking Cbl-b, the T cells respond to antigen even without CD28-mediated costimulation and produce abnormally high amounts of IL-2. These mice develop autoimmunity as a result of the enhanced activation of their T cells.")
56
Citokin szignaling

58
two different STATs may heterodimerize on phosphorylation
two different STATs may heterodimerize on phosphorylation. Therefore, there is a significant amount of combinatorial diversity in the signaling that can be generated from a limited number of JAK and STAT proteins. Gátlók SHP1, SHP2, SOCS ubiquitin ligáz…


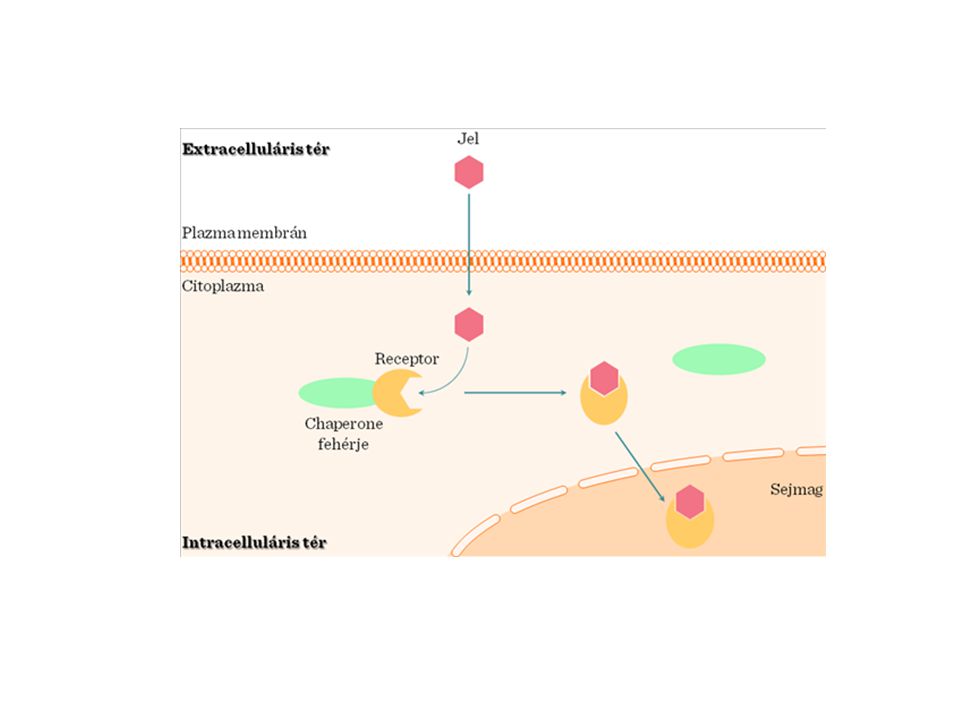
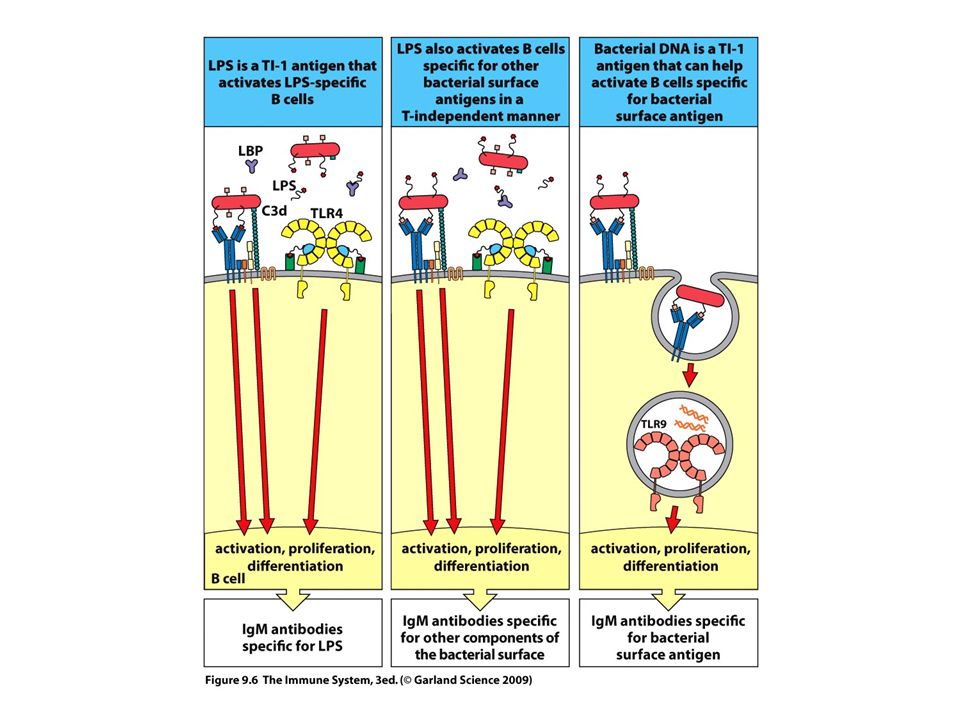
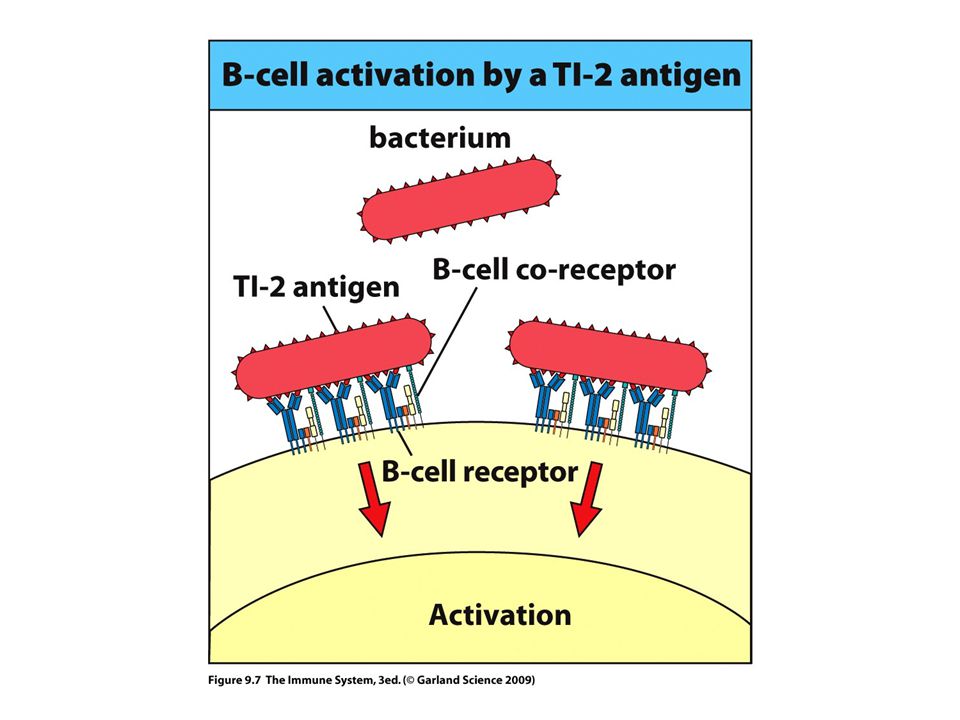
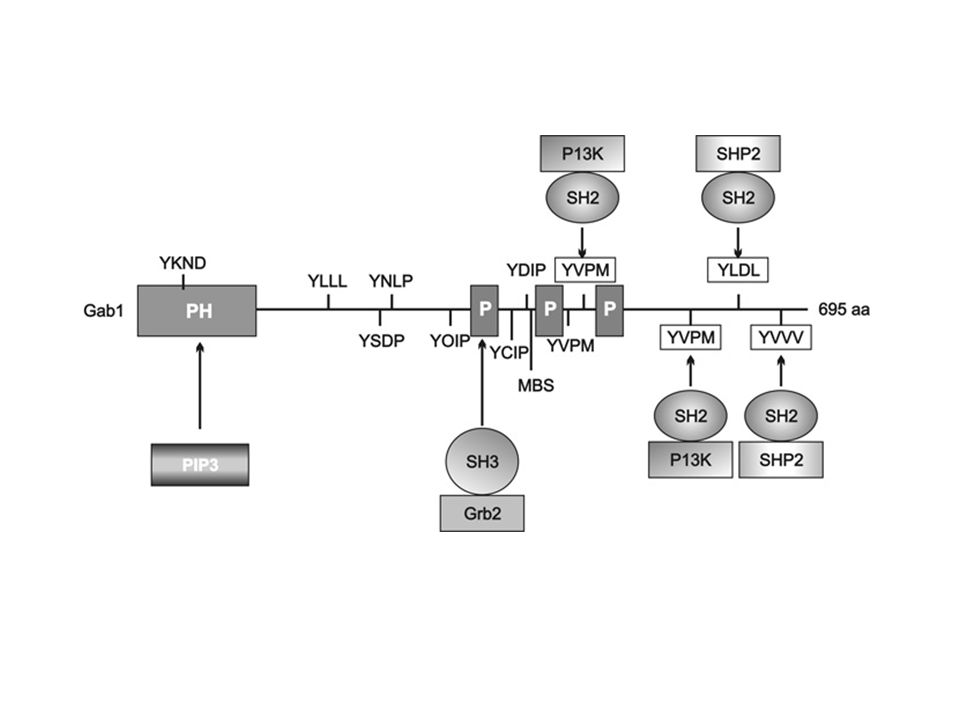









>")











>")
>")
