Előadást letölteni
Az előadás letöltése folymat van. Kérjük, várjon

1
Nem-receptor protein tirozin kinázok által elindított jelpályák
RECEPTOR TIROZIN KINÁZOK, Nem-receptor protein tirozin kinázok által elindított jelpályák Protein foszforiláció, protein foszfatázok

2
Tirozin kinázokhoz kapcsolt receptorok:
TCR, BCR, FcR, citokin receptorok

3
MA: Foszforilációs hely-specifikus ellenanyagok
Nem-receptor protein tirozin kinázok: vírusok transzformáló génjeiként azonosították Fehérjék tirozin foszorilációja és a malignus transzformáció összefüggése MA: Foszforilációs hely-specifikus ellenanyagok Polyoma middle-sized T antigen can be phosphorylated on tyrosine at multiple sites in vitro. T Hunter, M A Hutchinson, and W Eckhart Abstract The polyoma middle-sized T antigen (MT antigen) is associated with a protein kinase activity which phosphorylates tyrosine residues in polyoma T antigens in vitro. We have studied the sites of tyrosine phosphorylation of MT antigens phosphorylated in immunoprecipitates or in soluble form after partial purification by immunoaffinity chromatography. By analyzing the amino acid sequences of tryptic peptides of MT antigen, and by analyzing deletion mutant MT antigens, we have identified two major sites of phosphorylation in MT antigen, tyrosines 250 and 315. Additional sites were phosphorylated under some conditions. A synthetic peptide (Glu.Glu.Glu.Glu.Tyr.Met.Pro.Met.Glu), corresponding to the sequence around tyrosine 315, was phosphorylated when added to immunoprecipitates containing MT antigen.
 is associated with a protein kinase activity which phosphorylates tyrosine residues in polyoma T antigens in vitro. We have studied the sites of tyrosine phosphorylation of MT antigens phosphorylated in immunoprecipitates or in soluble form after partial purification by immunoaffinity chromatography. By analyzing the amino acid sequences of tryptic peptides of MT antigen, and by analyzing deletion mutant MT antigens, we have identified two major sites of phosphorylation in MT antigen, tyrosines 250 and 315. Additional sites were phosphorylated under some conditions. A synthetic peptide (Glu.Glu.Glu.Glu.Tyr.Met.Pro.Met.Glu), corresponding to the sequence around tyrosine 315, was phosphorylated when added to immunoprecipitates containing MT antigen.")
4
Tumor terápiában alkalmazható gyógyszerek

5
Nem receptor típusú protein tirozin kináz családok
Src, lyn, fyn, blk, yes, hck, lck Zap70, syk Receptorok az immunrenszerben pl.: BCR, TCR, citokin R, IgE, IgG receptorok

6
Hogyan kapcsolódnak a nem-receptor kinázok a receptorhoz?
Hogy aktiválódnak? Az acilezett fehérjék (pl. mirisztil csoporton keresztül a membránhoz kapcsolódó src –kinázok) és a GPI -kapcsolt fehérjék a koleszterinben gazdag lipid raftokban helyezkednek el
 és a GPI -kapcsolt fehérjék a koleszterinben gazdag lipid raftokban helyezkednek el.")
7
Glycosylphosphatidylinisotol (GPI)
")
8
Kináz célpontok: ITAM: Immunreceptor Tyrosine based Activation Motif

10
A receptor aktiváció citoszolban levő fehérjéket toborozhat a receptor közelébe.

11
Immunoreceptor Tirozin alapú Aktivációs Motívum
ITAM: Immunoreceptor Tirozin alapú Aktivációs Motívum D/E-x2- YxxL/I-x6-9Yx2L/I Az aktiváció előfeltétele a receptor oligomerizáció!

12
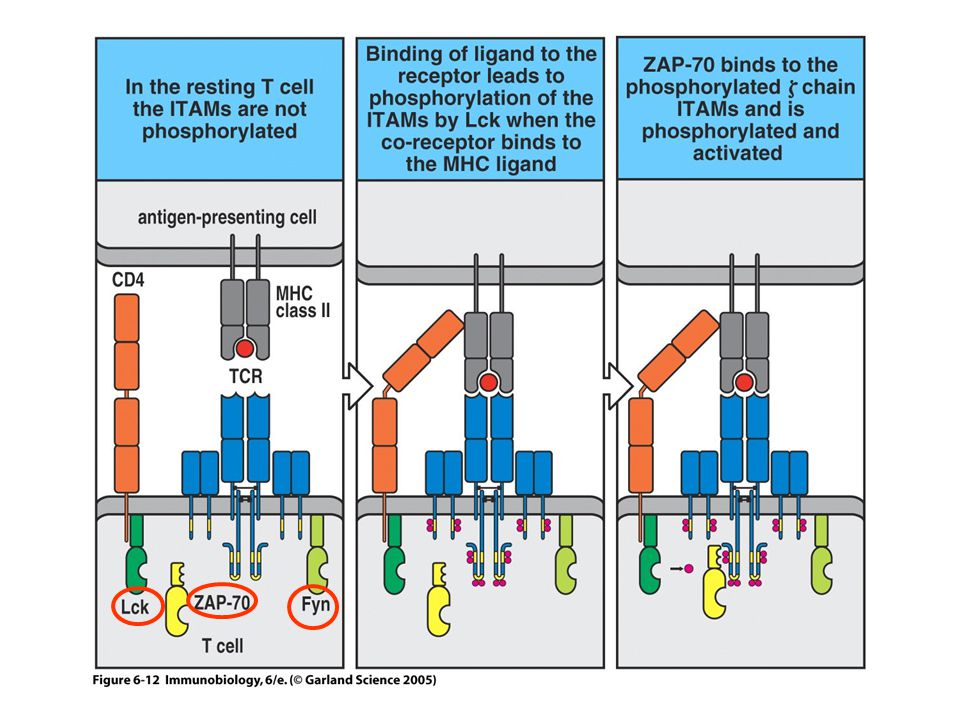
A TCR által indukált jelpálya kezdeti tirozin foszforilációs lépései
fyn

13
Figure 6-16

14
BCR

15
Figure 6-15 part 1 of 2

16
STAT aktiváció az interferon receptor jelátvitel során
IRF: interferon regulatory factor ISGF: interferon-a stimulated gene complex 3 ISRE: interferon stimulated response element

17
Az FceRI jelátvitel kezdeti lépései
mastoparan Mastoparan is a peptide toxin from wasp venom. It has the chemical structure Ile-Asn-Leu-Lys-Ala-Leu-Ala-Ala-Leu-Ala-Lys-Lys-Ile-Leu-NH2.[2] The net effect of mastoparan's mode of action depends on cell type, but seemingly always involves exocytosis. Inmast cells, this takes the form of histamine secretion, while in platelets and chromaffin cells release serotoninand catecholamines are found, respectively. Mastoporan activity in the anterior pituitary gland leads to prolactinrelease. In the case of histamine secretion, the effect of mastoparan takes place via its interference with G proteinactivity. By stimulating the GTPase activity of certain subunits, mastoparan shortens the lifespan of active G protein. At the same time, it promotes dissociation of any bound GDP from the protein, enhancing GTP binding. In effect, the GTP turnover of G proteins is greatly increased by mastoparan. These properties of the toxin follow from the fact that it structurally resembles activated G protein receptors when placed in a phospholipidenvironment. The resultant G protein-mediated signaling cascade leads to intracellular IP3 release and the resultant influx of Ca2+.

18
Az FcgR által aktivált kinázok

19
Mire jó a foszforiláció?
Megváltoztatja a konformációt P Intramolekuláris kölcsönhatások P P VÁLTOZIK: * Enzim/Faktor aktivitás * Lokalizáció * Stabilitás P Intermoleculáris kölcsönhatások

20
Protein foszfatázok ATP ADP Kinázok Foszfatázok P PO4 2-
* A foszforiláció reverzibilis * Enzimek aktívan katalizálják * Különböző enzimcsaládok ellentétes reakciókat katalizálnak FONTOS: Protein defoszforiláció független, fiziológiailag releváns folyamat Protein defoszforiláció ki és bekapcsolhat mechanizmusokat

21
Protein defoszforiláció
Kinázok P Foszfatázok pTyrosine pSerine pThreonine C H C H C H CH3 OPO3 2- OPO3 2- OPO3 2-

22
Tirozin specifikus (PTP) Receptor-típusú alcsalád
A foszfatázok fajtái: Tirozin specifikus (PTP) Receptor-típusú alcsalád Nem receptor-típusú alcsalád Kettős specifitású foszfatázok (Tyr és Ser/Thr) (PTEN) Kis molekulatömegű foszfatázok (LMW) Szerin/Treonin-specifikus foszfatázok PPP alcsalád PPM alcsalád Egyéb foszfatázok (eg. Hisztidine) Nem-specifikus foszfatázok (alkaline, savas foszfatázok) CX5R
 Receptor-típusú alcsalád. Nem receptor-típusú alcsalád. Kettős specifitású foszfatázok (Tyr és Ser/Thr) (PTEN) Kis molekulatömegű foszfatázok (LMW) Szerin/Treonin-specifikus foszfatázok. PPP alcsalád. PPM alcsalád. Egyéb foszfatázok (eg. Hisztidine) Nem-specifikus foszfatázok (alkaline, savas foszfatázok) CX5R.")
23
Protein tirozin foszfatáz családok
Tirozin-specifikus kettős specifitású PTEN LMW HCSAGXGRXG HCXXGXXR CXXXXXR CXXXXXR

24
Protein tirozin foszfatázok
Rövid története : * 1970-es évek vége - tirozin kinázok felfedezése * 1980-as évek: megfigyelték a tirozin defoszforilációt . fiziológiai folyamat farmakológiai sajátságok – vanadát gátol. * Nyitott kérdések: Hogy néznek ki a foszfatázok? (klónozás) Hány PTPáz van? Specifikus/nem-specifikus? Reguláció? Hatásmechanizmus? Fiziológia jelentőség?
 Hány PTPáz van Specifikus/nem-specifikus Reguláció Hatásmechanizmus Fiziológia jelentőség")
25
PTP specificitás és diverzitás: két extrém model
PTKs PTPs PTKs PTPs kevés PTP alacsony specificitás sok PTP nagy szubsztrát specificitás

26
Konklúzió: * PTP család, számos taggal, 1 v. 2 foszfatáz doménnel
2001-ben: * PTP szupercsalád 38 humán PTP gén 17 különböző szerkezetű család (8 receptor-szerű, 9 nem receptor-szerű). több tucat homológ fehérje
. több tucat homológ fehérje.")
27
(From: Andersen et al., MCB 21, 7117-7136, 2001)
HCSAGXGRXG

28
9Å A foszfatáz katalitikus domén vázlatosan:
Asp Cys * Bemélyedő zsebben van a kötőhely * A kritikus aminosavak ennek az aljában vannak Mélysége (9Å) túl mély a pSer, pThr maradékoknak -> specificitás További specificitást a hidrofób oldalláncok biztosítanak, amelyek stabilizálják a tirozin fenolgyűrűjét
 túl mély a pSer, pThr maradékoknak -> specificitás. További specificitást a hidrofób oldalláncok biztosítanak, amelyek stabilizálják a tirozin fenolgyűrűjét.")
29
Két lépésban defoszforilál!
PTP-Cys - S - 215 P -O O Asp181 OH A. Kovalens cisztein-foszfát kötés, a tirozin felszabadul. PTP-Cys - S- H O- (From Barford et al., Nature Structural Biology 2, , 1995) B. A cisztein-foszfor kötés hidrolízise, az enzim regenerálódása. Két lépésban defoszforilál!
 B. A cisztein-foszfor kötés hidrolízise, az enzim regenerálódása. Két lépésban defoszforilál!")
30
PTP-hez kapcsolódó szabályozás:
PTP aktivitás szabályozása: receptor típusú PTP-k dimerizálódása (CD45, RPTPa). SH2 domén kötődés H2O2-tól függő szabályozás SRC családba tartozó kinázok aktiválása: tirozin defoszforiláció útján
. SH2 domén kötődés. H2O2-tól függő szabályozás. SRC családba tartozó kinázok aktiválása: tirozin defoszforiláció útján.")
31
CD45 PTP1B hasonló a CD45-höz PTP1B PTP1B T-sejt PTP CD45 LAR 74%
37% % CD45 44% % LAR

32
Ligandum közvetített dimerizáció
Probléma: kevés ligandum ismert. a ligandum kötődés következményei nem ismertek. Hogyan tanulmányozható a dimerizálódás ligandum nélkül?? Megoldás: EGFR-CD45 kiméra molekula EGFR membrán CD45

33
A ligandum kötődése és a dimerizáció gátló hatású!
A CD45 elengedhetetlen a T sejtek aktivációjához (Jurkat sejt ) CD45-deficiens Jurkat T-sejtek (plusz stimuláció TCR-en keresztül) EGFR-CD45 expresszió plusz EGF Nincs TCR signaling (nem transzfektált sejtek) Nincs TCR signaling CD45 expresszió TCR signaling EGFR-CD45 expresszió TCR signaling Conclusion: Ligand binding and dimerization is inhibitory! A ligandum kötődése és a dimerizáció gátló hatású! From: Desai et al., Cell 73, , 1993
 CD45-deficiens Jurkat T-sejtek. (plusz stimuláció TCR-en keresztül) EGFR-CD45. expresszió. plusz EGF. Nincs TCR signaling. (nem transzfektált. sejtek) Nincs TCR signaling. CD45. expresszió. TCR signaling. EGFR-CD45. expresszió. TCR signaling. Conclusion: Ligand binding and dimerization is inhibitory! A ligandum kötődése és a dimerizáció gátló hatású! From: Desai et al., Cell 73, ,")
34
A ligandum kötődése és a dimerizáció gátló hatású!
Hogyan? 1996: röntgenszerkezet : RPTPa, D1 domén: 1. Dimerizálódás 2. A “wedge” domén az N-terminuson, gátolja a katalitikus doménhez való hozzáférést “wedge” domain Based on: Bilwes et al., Nature 382, , 1996 Catalytic cleft

35
Wedge domén gátolja a PTP activitását a dimerben.
A mutációt hordozó wedge domén : megakadályozza az inaktivációt (foszfatáz aktivitás nő): E613R CD45 hiper-aktív “knock-in” egerekben in vivo; Egerekben limfocita proliferáció, ellenanyag termelés halál (Majeti et al., Cell 103, , 2000)
: E613R CD45 hiper-aktív knock-in egerekben in vivo; Egerekben limfocita proliferáció, ellenanyag termelés halál. (Majeti et al., Cell 103, , 2000)")
36
PTK és PTP ligandum által indukált aktiválódásának összehasonlítása
Active Inactive RPTK Inactive Ligand RPTP Active After: Weiss and Schlessinger,Cell 94, , 1998

37
A foszforiláció szabályozásának módjai:
SH2 domén által közvetített H2O2 által közvetített

38
Foszforiláció szabályozás pY SH2 doménhez való kötődése által
Kinase Y 416 527 p Src activity: Src phosphorylation, part I autofoszforiláció

39
CSK Src phosphorylation, part II C terminális src kináz p Y 416 Y 527
SH3 SH2 Kinase Src activity:

40
SRC aktiválódása Tyr defoszforiláció útján
SH3 P P P SH2 PTK pY527 Inactive conformation (“closed”) SH3 PTK Y527 P pY416 Active conformation (“open”) SH2
 SH3. PTK. Y527. P. pY416. Active conformation. ( open ) SH2.")
41
H2O2-től függő szabályozás
ROS felszabadulás biológiai jelekre reactive oxygen species Biológiai stimulusok, amelyek indukálják a ROS keletkezését: Fig. 1. Biological stimuli associated with the generation of reactive oxygen species (ROS). The main sources of intracellular ROS are illustrated and the specific stimuli that elicit the response are listed [57]. (a) The mitochondrial generation of ROS represents a relevant by-product of electron flow through the respiratory chain (complexes 1–4 are indicated in blue). Oxygen is converted to O2 z2 by ubiquinone (UQ)-mediated one-electron reduction, which is then converted to H2O2 by spontaneous or enzymatic dismutation that is mediated by superoxide dismutase. Mitochondrial ROS production is itself modulated by a variety of stimuli (listed), and, at least for integrin signaling, is reported to be Rac dependent [7]. (b) Upon activation, both NADPH oxidase and lypoxygenase (or cycloxygenase) are translocated to the membrane and, therefore, the small GTPase Rac is activated. Both oxidases produced O2 z2, which is then converted to H2O2 by spontaneous or enzymatic-dismutation mediated by superoxide dismutase. Abbreviations: AA, arachidonic acid; HPETE, hydroperoxyeicosatetraenoaic acid. Légzési elektrontranszport lánc
. The main sources of intracellular ROS are illustrated and the specific stimuli that elicit the response are listed [57]. (a) The mitochondrial generation of ROS represents a relevant by-product of electron flow through the respiratory chain (complexes 1–4 are indicated in blue). Oxygen is converted to O2 z2 by ubiquinone (UQ)-mediated one-electron reduction, which is then converted to H2O2 by spontaneous or enzymatic dismutation that is mediated by superoxide dismutase. Mitochondrial ROS production is itself modulated by a variety of stimuli (listed), and, at least for integrin signaling, is reported to be Rac dependent [7]. (b) Upon activation, both NADPH oxidase and lypoxygenase (or cycloxygenase) are translocated to the membrane and, therefore, the small GTPase Rac is activated. Both oxidases produced O2 z2, which is then converted to H2O2 by spontaneous or enzymatic-dismutation mediated by superoxide dismutase. Abbreviations: AA, arachidonic acid; HPETE, hydroperoxyeicosatetraenoaic acid. Légzési elektrontranszport lánc.")
42
H2O2 által történő PTP szabályozás
pTyr defoszforiláció PTPs: pKa= Cys-SH Cys-S- Cys-S-OH H2O2 (Sulfenic acid) DTT Thioredoxin Magasabb oxidációs állapot, inaktív, irreverzibilis (vanadate ions) Acid dissociation constant, Ka, (also known as acidity constant, or acid-ionization constant) is a quantitative measure of the strength of an acid in solution
 DTT. Thioredoxin. Magasabb oxidációs állapot, inaktív, irreverzibilis. (vanadate ions) Acid dissociation constant, Ka, (also known as acidity constant, or acid-ionization constant) is a quantitative measure of the strength of an acid in solution.")
43
Konklúzió: H2O2 képes enzimeket megtámadni, ha azok rendelkeznek
kritikus helyzetben levő ciszteinnel H2O2 különösen befolyásolhat olyan fehérjéket, amelyekben a Cys alacsony pKa –val rendelkezik, mint amilyenek a foszfatázok H2O2 relatív gyenge oxidáns, amelynek aktivitását a sejtben meglévő mechanizmusok szabályozzák. Reverzibilis hatású. H2O2 keletkezhet fiziológiás jelátviteli folyamatok során a sejtekben * H2O2 mint másodlagos hírvivő fejtheti ki hatását.

44
(From: Andersen et al., MCB 21, 7117-7136, 2001)
")
45
SH2 doméneket tartalmazó PTP-k:
SHP1 64% 53% 57% SHP2

46
Intramolekuláris kötés
SH3 P P P SH2 PTK pY527 Inactive conformation (“closed”) SH3 PTK Y527 P pY416 Active conformation (“open”) SH2 N-SH2 C-SH2 PTP P Active conformation (“open”) N-SH2 C-SH2 PTP pTyr binding site Inactive conformation (“closed”) Src SHP2 After: Weiss and Schlessinger,Cell 94, , 1998
 SH3. PTK. Y527. P. pY416. Active conformation. ( open ) SH2. N-SH2. C-SH2. PTP. P. Active conformation. ( open ) N-SH2. C-SH2. PTP. pTyr binding site. Inactive conformation. ( closed ) Src. SHP2. After: Weiss and Schlessinger,Cell 94, ,")
47
Az SHP-2 által történő pozitív jelpálya szabályozás modelljei
Fig. 2. Models for positive signaling by Src homology-2 (SH2) domain-containing phosphatase-2 (Shp2). (a) Regulation of Ras by controlling RasGap recruitment. Shp2 (and its orthologs) are recruited directly to some receptor tyrosine kinases (RTKs), such as the platelet-derived growth factor receptor (PDGFR) and Torso. Ligand binding results in receptor activation and phosphorylation of multiple receptor-tyrosyl residues. These phosphotyrosines (Tyr–P) recruit signal relay molecules containing SH2 and/or PTB domains, which in turn, lead to activation of downstream signaling cascades. One such pathway, the Ras/Raf/Erk cascade is shown. Ras is activated by recruitment of the adaptor Grb2, which binds to receptors either directly or indirectly via a second adaptor, Shc. Grb2 binds a guanine nucleotide exchange factor (GEF), which activates Ras by promoting GDP release, so that GTP can bind to Ras. Ras inactivation is catalyzed by GTPase activating proteins (GAPs), which promote hydrolysis of RasGTP to RasGDP. One such GAP, RasGap, has SH2 domains and binds to some receptors. Shp2 can dephosphorylate these RasGap binding sites, thereby preventing RasGap recruitment to receptors until appropriate signaling has occurred. (b) Regulation of SFK. Shown are several possible models by which Shp2, upon binding to scaffolding adapters such as Gab family proteins, might regulate Src family kinases (SFKs). Shp2 might directly dephosphorylate the inhibitory tyrosyl residues of a pool of SFKs (left). Alternatively, Shp2 might promote dephosphorylation of SFK inhibitory tyrosines indirectly. For example, Shp2 could dephosphorylate the transmembrane protein PAG/CBP, which recruits and activates Csk, the kinase that phosphorylates SFK inhibitory tyrosines (right). Alternatively, Shp2 might somehow regulate RPTPa, which directly dephosphorylates the inhibitory tyrosyl residues (?) Ultimately, SFK might contribute to Ras activation by as yet unclear mechanisms. Broken lines indicate unknown and/or unclear pathways; solid lines indicate established mechanisms. (c) Regulation of Sprouty and/or downstream pathways. Sprouty proteins inhibit Ras activation; this requires tyrosyl phosphorylation of Sprouty. Thus, dephosphorylation of Sprouty by Shp2 could promote Ras activation. This figure also shows that Shp2 has additional actions downstream of Ras. For example, acting through Gab2, Shp2 can regulate Elk-mediated transcription, by an as yet unknown mechanism (broken line with question mark). Initial work suggested an ‘adapter’ model, in which Shp2 becomes tyrosyl phosphorylated in response to RTK stimulation and then binds Grb2/Sos [3]. However, the C-terminal tyrosyl residues are dispensable in Xenopus FGFR [47] and Sevenless [48] signaling. By contrast, the PTP activity of Shp2 is essential for positive signaling, although the key target(s) remain(s) unclear. Shp2 preferentially dephosphorylates the RasGap binding site on the PDGFR. Az SHP-2 által történő pozitív jelpálya szabályozás modelljei
 domain-containing phosphatase-2 (Shp2). (a) Regulation of Ras by controlling RasGap recruitment. Shp2 (and its orthologs) are recruited directly to some receptor tyrosine kinases (RTKs), such as the platelet-derived growth factor receptor (PDGFR) and Torso. Ligand binding results in receptor activation and phosphorylation of multiple receptor-tyrosyl residues. These phosphotyrosines (Tyr–P) recruit signal relay molecules containing SH2 and/or PTB domains, which in turn, lead to activation of downstream signaling cascades. One such pathway, the Ras/Raf/Erk cascade is shown. Ras is activated by recruitment of the adaptor Grb2, which binds to receptors either directly or indirectly via a second adaptor, Shc. Grb2 binds a guanine nucleotide exchange factor (GEF), which activates Ras by promoting GDP release, so that GTP can bind to Ras. Ras inactivation is catalyzed by GTPase activating proteins (GAPs), which promote hydrolysis of RasGTP to RasGDP. One such GAP, RasGap, has SH2 domains and binds to some receptors. Shp2 can dephosphorylate these RasGap binding sites, thereby preventing RasGap recruitment to receptors until appropriate signaling has occurred. (b) Regulation of SFK. Shown are several possible models by which Shp2, upon binding to scaffolding adapters such as Gab family proteins, might regulate Src family kinases (SFKs). Shp2 might directly dephosphorylate the inhibitory tyrosyl residues of a pool of SFKs (left). Alternatively, Shp2 might promote dephosphorylation of SFK inhibitory tyrosines indirectly. For example, Shp2 could dephosphorylate the transmembrane protein PAG/CBP, which recruits and activates Csk, the kinase that phosphorylates SFK inhibitory tyrosines (right). Alternatively, Shp2 might somehow regulate RPTPa, which directly dephosphorylates the inhibitory tyrosyl residues ( ) Ultimately, SFK might contribute to Ras activation by as yet unclear mechanisms. Broken lines indicate unknown and/or unclear pathways; solid lines indicate established mechanisms. (c) Regulation of Sprouty and/or downstream pathways. Sprouty proteins inhibit Ras activation; this requires tyrosyl phosphorylation of Sprouty. Thus, dephosphorylation of Sprouty by Shp2 could promote Ras activation. This figure also shows that Shp2 has additional actions downstream of Ras. For example, acting through Gab2, Shp2 can regulate Elk-mediated transcription, by an as yet unknown mechanism (broken line with question mark). Initial work suggested an ‘adapter’ model, in which Shp2 becomes tyrosyl phosphorylated in response to RTK stimulation and then binds Grb2/Sos [3]. However, the C-terminal tyrosyl residues are dispensable in Xenopus FGFR [47] and Sevenless [48] signaling. By contrast, the PTP activity of Shp2 is essential for positive signaling, although the key target(s) remain(s) unclear. Shp2 preferentially dephosphorylates the RasGap binding site on the PDGFR. Az SHP-2 által történő pozitív jelpálya szabályozás modelljei.")
48
Tirozin specifikus (PTP) Receptor-típusú alcsalád
A foszfatázok fajtái: Tirozin specifikus (PTP) Receptor-típusú alcsalád Nem receptor-típusú alcsalád Kettős specifitású foszfatázok (Tyr és Ser/Thr) (PTEN, Myotubularin) Kis molekulatömegű foszfatázok (LMW) Szerin/Treonin-specifikus foszfatázok PPP alcsalád PPM alcsalád Egyéb foszfatázok (eg. Hisztidine) Nem-specifikus foszfatázok (alkaline, savas foszfatázok) CX5R
 Receptor-típusú alcsalád. Nem receptor-típusú alcsalád. Kettős specifitású foszfatázok (Tyr és Ser/Thr) (PTEN, Myotubularin) Kis molekulatömegű foszfatázok (LMW) Szerin/Treonin-specifikus foszfatázok. PPP alcsalád. PPM alcsalád. Egyéb foszfatázok (eg. Hisztidine) Nem-specifikus foszfatázok (alkaline, savas foszfatázok) CX5R.")
49
Kettős specifitású foszfatázok
* Tirozin ÉS Szerin/Treonin defoszforiláció. * Molekuláris mechanizmus a tirozin foszfatázokéhoz hasonló (cystein, etc.) * Sekélyebb katalitikus zseb * in vitro nagy szubsztrát specificitás
 * Sekélyebb katalitikus zseb. * in vitro nagy szubsztrát specificitás.")
50
MKP- MAP Kináz foszfatázok
Stimulus MAP kinase kinase kinase MAP kinase kinase MAP kinase Substrates

51
T X Y MAPKK T X Y P T X Y P Tyr PTPs (HePTP STEP PTP-SL) T X Y P
After: Camps et al., Faseb J. 14, 6-16, 1999 T X Y MAPK MAPKK T X Y P T X Y P Tyr PTPs (HePTP STEP PTP-SL) MAPK T X Y P Ser/Thr PPs (PP2A) T X Y Dual-specificity PPs
 MAPK. T. X. Y. P. Ser/Thr PPs. (PP2A) T. X. Y. Dual-specificity PPs.")
52
Tirozin specifikus (PTP) Receptor-típusú alcsalád
A foszfatázok fajtái: Tirozin specifikus (PTP) Receptor-típusú alcsalád Nem receptor-típusú alcsalád Kettős specifitású foszfatázok (Tyr és Ser/Thr) (PTEN, Myotubularin) Kis molekulatömegű foszfatázok (LMW) Szerin/Treonin-specifikus foszfatázok PPP alcsalád PPM alcsalád Egyéb foszfatázok (eg. Hisztidine) Nem-specifikus foszfatázok (alkaline, savas foszfatázok) CX5R
 Receptor-típusú alcsalád. Nem receptor-típusú alcsalád. Kettős specifitású foszfatázok (Tyr és Ser/Thr) (PTEN, Myotubularin) Kis molekulatömegű foszfatázok (LMW) Szerin/Treonin-specifikus foszfatázok. PPP alcsalád. PPM alcsalád. Egyéb foszfatázok (eg. Hisztidine) Nem-specifikus foszfatázok (alkaline, savas foszfatázok) CX5R.")
53
Kis molekula tömegű (LMW) tirozin foszfatázok
CXXXXXR motif “standard” PTP ~240 residues (minimal MW=~26-30 kDa) ~160 residues (MW=~18 kDa) Overall, sharp differences in 3-dimensional structure Low MW PTP (after: Ramponi & Stefani, BBA 1341, , 1997)
 ~160 residues. (MW=~18 kDa) Overall, sharp differences. in 3-dimensional structure. Low MW PTP. (after: Ramponi & Stefani, BBA 1341, , 1997)")
54
Major types of protein phosphatases
Tyrosine-specific (PTP) Receptor-type subfamily Non receptor-type subfamily Dual Specificity (Tyr and Ser/Thr) Low Molecular Weight (LMW) Serine/Threonine-specific PPP subfamily PPM subfamily Other phosphatases (eg. Histindine) Non-specific phosphatases (alkaline, acid phosphatases) CX5R Serine/Threonine-specific PPP subfamily PPM subfamily
 Receptor-type subfamily. Non receptor-type subfamily. Dual Specificity (Tyr and Ser/Thr) Low Molecular Weight (LMW) Serine/Threonine-specific. PPP subfamily. PPM subfamily. Other phosphatases (eg. Histindine) Non-specific phosphatases (alkaline, acid phosphatases) CX5R. Serine/Threonine-specific. PPP subfamily. PPM subfamily.")
55
Szerin-treonin foszfatázok
Sematikus szerkezet: A B regulátor scaffold C katalitikus

56
Alapvető különbségek a ser/Thr és Tyr foszfatázok között :
C Ser/Thr Tyr Szerkezet: Holoenzimek egy láncú enzimek Variabilitás: kevés katalitikus alegység sok féle enzim Specificitás: mindhárom alegység szabja egyetlen enzim meg Katalitikus mechanizmus: Teljesen eltérő Farmakológia: Teljesen eltérő

57
(Protein phosphatase 2 (PP2))
Ser/Thr Phosphatases PPP subfamily PPM subfamily PP2C PP1 PP2A PP2B PPM family, which includes PP2C and pyruvate dehydrogenase phosphatase, are enzymes with Mn2+/Mg2+ metal ions that are resistant to classic inhibitors (Protein phosphatase 2 (PP2)) PPP1 PPP2A PPP5 Célpont: Raf MEK Akt So many functions, so few enzymes?
) PPP1. PPP2A. PPP5. Célpont: Raf. MEK. Akt. So many functions, so few enzymes")
58
Variability = több tucat molekuláris kapcsolat! PP2A PR55 B
(a, b, g, d) Variability Scaffold: PR65 (a or b) B’ A PR61 (a, b, g, d, e) C B’’ Mammalian protein phosphatase 2A (PP2A) holoenzymes. The catalytic subunit (C) is bound constitutively to a scaffold subunit (A or PR65). This heterodimer can further complex with any one of an array of B regulatory subunits, whose binding to the core dimer appears to be mutually exclusive. Two isoforms of the C subunit (a and b) and two isoforms of the PR65 (a and b) exist. Three isoforms of PR55 have been identified (a, and the brain-specific b and g isoforms). Five genes (a, b, g, d and e) encode members of the PR61 (also known as the B9 or B56) family, and some transcripts undergo alternative splicing to generate up to ~11 isoforms. The B99 family comprises 72-kDa (PR72) and 130-kDa (PR130) subunits, which are generated by alternative splicing of a single gene, as well as PR59, the product of a closely related gene. PR72, PR130 PR59, PR48 B’’’ Regulátor alegységek Catalytic: Ca or Cb Konstitutív kötődés PR93/SG2NA PR110/striatin
 Variability. Scaffold: PR65. (a or b) B’ A. PR61. (a, b, g, d, e) C. B’’ Mammalian protein phosphatase 2A (PP2A) holoenzymes. The catalytic subunit (C) is bound. constitutively to a scaffold subunit (A or PR65). This heterodimer can further complex with. any one of an array of B regulatory subunits, whose binding to the core dimer appears to be. mutually exclusive. Two isoforms of the C subunit (a and b) and two isoforms of the PR65 (a. and b) exist. Three isoforms of PR55 have been identified (a, and the brain-specific b. and g isoforms). Five genes (a, b, g, d and e) encode members of the PR61 (also known as. the B9 or B56) family, and some transcripts undergo alternative splicing to generate up to. ~11 isoforms. The B99 family comprises 72-kDa (PR72) and 130-kDa (PR130) subunits, which are generated by alternative splicing of a single gene, as well as PR59, the product of. a closely related gene. PR72, PR130. PR59, PR48. B’’’ Regulátor alegységek. Catalytic: Ca or Cb. Konstitutív kötődés. PR93/SG2NA. PR110/striatin.")
59
Ser/Thr foszfatázok PPP subfamily PP1 PP2A PP2B PP2C PPM subfamily calcineurin Gátlószer: (okadaic acid microcystin) Gátlószer (FK506 cyclosporin) Ca++ függő Aktív hely: Mn++, Fe++ Mn++,/ 2X Mg++ Hasonló: alapszerkezet, általános alak, alapvető katalitikus mechanizmus Eltérő: szekvencia, katalitikus mechanizmus részletei, biokémiája.
 Gátlószer. (FK506. cyclosporin) Ca++ függő. Aktív hely: Mn++, Fe++ Mn++,/ 2X Mg++ Hasonló: alapszerkezet, általános alak, alapvető katalitikus mechanizmus. Eltérő: szekvencia, katalitikus mechanizmus részletei, biokémiája.")
60
Példa a PP2A-függő szabályozásra fiziológiás folyamat során:
(from: Millward et al., TIBS 24, , 1999) Insulin receptor IRS 1/2 PI3-K p85 p110 PtdIns(3,4)P2 or PtdIns(3,4,5)P3 Active PKB ATP ADP PDK1 PH kinase Inactive PKB Active PKB (Downstream signaling events) Pi PP2A Regulation of protein kinase B (PKB) by PDK1 and protein phosphatase 2A (PP2A). Binding of insulin to its receptor causes activation of phosphoinositide 3-kinase (PI3-K), which generates phosphatidylinositol 3,4-bisphosphate [PtdIns(3,4) P2] and phosphatidylinositol 3,4,5-trisphosphate [PtdIns(3,4,5) P3] in the plasma membrane. These phospholipids are ligands for the pleckstrin homology (PH) domain of PKB, and bring about recruitment of PKB to the membrane, as well as a conformational change in PKB. Binding of PtdIns(3,4) P2 or PtdIns(3,4,5) P3 converts PKB into a substrate for two distinct kinase kinases: PDK1 and Ser473 kinase. Each of these phosphorylates PKB on a different site; together they cause its activation. Thr308 of PKB is phosphorylated by PDK1, which is itself membrane localized through a PH domain (not shown); the Ser473 kinase has not yet been identified. Activated PKB detaches from the membrane to phosphorylate various cytosolic and nuclear substrates. The phosphatase responsible for returning PKB to its inactive state is PP2A. Inactive PKB kinase PH (PKB=AKT, =Ser/Thr kinase)
 Insulin. receptor. IRS. 1/2. PI3-K. p85. p110. PtdIns(3,4)P2 or. PtdIns(3,4,5)P3. Active. PKB. ATP. ADP. PDK1. PH. kinase. Inactive. PKB. Active. PKB. (Downstream signaling events) Pi. PP2A. Regulation of protein kinase B (PKB) by PDK1 and protein phosphatase 2A (PP2A). Binding of insulin to its receptor causes activation of phosphoinositide 3-kinase (PI3-K), which. generates phosphatidylinositol 3,4-bisphosphate [PtdIns(3,4) P2] and phosphatidylinositol 3,4,5-trisphosphate [PtdIns(3,4,5) P3] in the plasma membrane. These phospholipids are ligands for the pleckstrin homology (PH) domain of PKB, and bring about recruitment of PKB to the membrane, as well as a conformational change in PKB. Binding of PtdIns(3,4) P2 or PtdIns(3,4,5) P3 converts PKB into a substrate for two distinct kinase kinases: PDK1 and Ser473 kinase. Each of these phosphorylates PKB on a different site; together they cause its activation. Thr308 of PKB is phosphorylated by PDK1, which is itself membrane localized through a PH domain (not shown); the Ser473 kinase has not yet been identified. Activated PKB detaches from the membrane to phosphorylate various cytosolic and nuclear substrates. The phosphatase responsible for returning PKB to its inactive state is PP2A. Inactive. PKB. kinase. PH. (PKB=AKT, =Ser/Thr kinase)")
61
Negatív visszacsatolás
Péda a PP2A-függő szabályozó hatására (2) T X Y MAPK MAPKK T X Y P T X Y P Tyr PTPs (HePTP STEP PTP-SL) MAPK PP2A T X Y P Ser/Thr PPs T X Y Dual-specificity PPs In vitro, PP2A can dephosphorylate and inactivate MEK1 and ERK-family kinases 33,34, and both kinases are activated after treatment of cells with OA35,36. Transient expression of SV40 small t antigen (which inhibits PP2A) activates the MEK1 and ERK, which might explain how small t antigen promotes transformation37; conversely, overexpression of casein kinase 2a (which activates PP2A) suppresses the activity of MEK1. Note that PP2A is not the only phosphatase that is important for the inactivation of ERKs. As mentioned above, PP2A operates in several cell types in conjunction with an unidentified tyrosine specific phosphatase to dephosphorylate the regulatory threonine and tyrosine residues in the activation loop of ERK. However, in other cell types, both phosphates are removed by a single dualspecificity phosphatase, MKP-1. MKP-1 is the product of an immediate-early gene that is transcriptionally upregulated following activation of the ERK MAP-kinase cascade, and therefore forms part of a negative feedback loop . MAPK cascade Negatív visszacsatolás
 T. X. Y. MAPK. MAPKK. T. X. Y. P. T. X. Y. P. Tyr PTPs. (HePTP. STEP. PTP-SL) MAPK. PP2A. T. X. Y. P. Ser/Thr PPs. T. X. Y. Dual-specificity PPs. In vitro, PP2A can dephosphorylate and inactivate MEK1 and ERK-family kinases 33,34, and both kinases are activated after treatment of cells with OA35,36. Transient expression of SV40 small t antigen (which inhibits PP2A) activates the MEK1 and ERK, which might explain how small t antigen promotes transformation37; conversely, overexpression of casein kinase 2a (which activates PP2A) suppresses the activity of MEK1. Note that PP2A is not the only phosphatase that is important for the inactivation of ERKs. As mentioned above, PP2A operates in several cell types in conjunction with an unidentified tyrosine specific phosphatase to dephosphorylate the regulatory threonine and tyrosine residues in the activation loop of ERK. However, in other cell types, both phosphates are removed by a single dualspecificity phosphatase, MKP-1. MKP-1 is the product of an immediate-early gene that is transcriptionally upregulated following activation of the ERK MAP-kinase cascade, and therefore forms part of a negative feedback loop . MAPK cascade. Negatív visszacsatolás.")
Hasonló előadás














>")
>")




