Előadást letölteni
1
2. A víz felvétele és transzportja, transzspiráció

2
2.1. A sejtmembrán vízvezető képessége A hidraulikus konduktivitás

3
A sejtmembrán ellenállásának nyomásfüggése, a „kritikus nyomásérték”

4
A transzcelluláris ozmózis és a poláris permeabilitás

6
A diffúziós permeabilitás és a hidraulikus konduktivitás viszonya

7
A reflexiós koefficiens

8
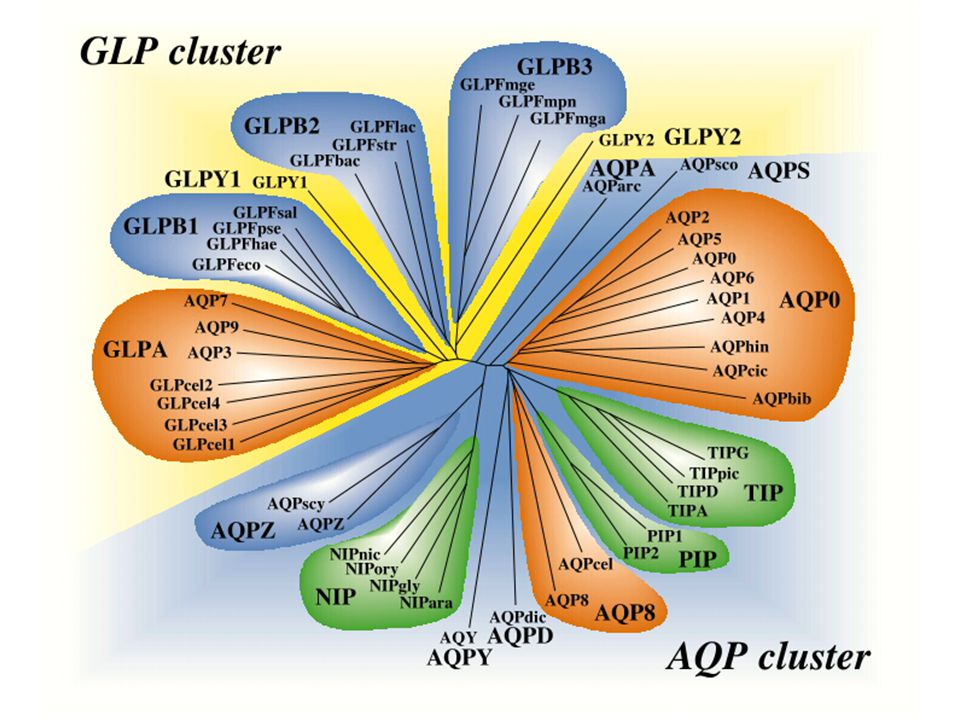
Aquaporinok

10
Struktúra és funkció: elsődleges struktúra -a legtöbb MIP tag: 250-300 aminosav, Mwt: 27-31 kDa -összehasonlítva egyes organizmusok elsődleges struktúráját MIP-jeik tekintetében néhány szakasz erősen konzervált -MIP-ek 46 típusa mutat filogenetikai rokonságot -szerkezete: 8 szomszédos struktúra mely tartalmaz 6 hidrofób transzmembrán hélixet, és 2 funkcionális B és E hurkot -B és E-ben az aszparagin-prolin- alanin (NPA) motívum erősen konzervált Aquaporin struktúra a AQP-2 példáján
 motívum erősen konzervált Aquaporin struktúra a AQP-2 példáján")
11
Felépítés és funkciók -Jung: CHIP28-ról 3D óraüvegmodell -B és E hurkok feltekeredtek és a memb.- ban találkoztak, az NPA motívumok átfedésekkel egy szűk csatornát alakítanak ki -aquaporinok szelektivitása: nemcsak a térbeli korlátoktól függ -AQP1 3A átmérőjű így akadályozza a nagyobb mol. áthaladását -pórusok legszűkebb része hidrofób kivéve a két központi aszparagin maradványt az NPA motívumon belül

12
-vízmolekula keresztülhaladása: az O és aszparagin közti kölcsönhatás úgy orientálja a 2 H atomot, hogy megakadályozza a H-kötések kialakulását a szomsz. vízmolekulák közt, ezalatt protonok átvitele korlátozott ebben a régióban -ionokra vonatkoztatva: pórusok térbeli korlátozottsága létrehoz egy dielektromos gátat, ami eltaszítja az ionokat -AQP1 tetramerikus formájú -in vitro növ.-i aggregátumokban oligomerek lehetnek

14
-a legtöbb aqv. igazoltan szenzitív a Hg komponensekre, ezek reverzibilisen blokkolják a víztranszportot, kötődve a cisztein maradványok SH csoportjaihoz -léteznek Hg inszenzitív aqv.-ok is -tonoplaszt vízpermeabilitása nagyobbá válhat, ezt igazolja a gyökér növekedésekor vagy vízpotenciál csökkenéskor a TIP- ek expressziója -a PIP-ek szerepe a víztranszport regulációja, optimalizálja a vízcserét az apoplaszt és a vakuolum közt Aquaporin aktivitás

15
Aquaporinok regulációja -túlzott vízveszteség hatására a víz permeabilitása csökken és csökken az aquaporinok aktivitása - de: vízstressz hatására génexpresszió is történhet (ez a szárazságtűrésben játszhat szerepet) -biotikus és abiotikus stresszorok, környezeti változások, patogének okozta károk összefüggésbe hozhatók az aqv.-ok aktivációjával és inaktivációjával -gabona gyökerében só-stresszre drámai változás e tekintetben: Pos ozmotikus gradiens erősen csökkent, az aqv.-ok záródtak válaszolván a víz deficitre
 -biotikus és abiotikus stresszorok, környezeti változások, patogének okozta károk összefüggésbe hozhatók az aqv.-ok aktivációjával és inaktivációjával -gabona gyökerében só-stresszre drámai változás e tekintetben: Pos ozmotikus gradiens erősen csökkent, az aqv.-ok záródtak válaszolván a víz deficitre")
16
Génexpresszió transzkripciós regulációja -víz-stressz erős hatással van a MIP gének expressziójára -különböző növ.-ben különböző mintázatúak az egyes MIP gének, ezek lehetnek specifikusak Post-transzlációs reguláció -legegyszerűbb post-transzlációs modifikáció: foszforiláció, mely konformációváltozással járhat

17
2.2. Víztranszport a gyökérben

18
A gyökér szerkezete keresztmetszeti képen A Caspary pontos endodermisz elzárja a sejtfalon keresztüli víztranszportot a xilémbe

19
A víz transzportja a gyökérben történhet az 1.apoplasztban, az összefüggő sejtfalrendszerben 2.A szimplasztban, a sejtek összefüggő citoplazmájában 3.Sejtről sejtre, transzcellulárisan

20
2.2. Víztranszport a gyökérben

21
Az exudáció („könnyezés”) Jv =Lp σRT(Cx – Co) Jv = gyölér vízfelvételi sebessége Lp = vízvezető képesség Σ = szállított anyagok reflexiós koefficiense Cx = xilém elemekben lévő oldat koncentrációja Co = közeg koncentrációja
 Jv =Lp σRT(Cx – Co) Jv = gyölér vízfelvételi sebessége Lp = vízvezető képesség Σ = szállított anyagok reflexiós koefficiense Cx = xilém elemekben lévő oldat koncentrációja Co = közeg koncentrációja")
22
A gyökér hidraulikus konduktivitása (Lpr)
")
24
A gyökérzónák vízfelvétele a transzspiráció intenzitásától függően I-V, csúcsi – alapi zónák A-D, fokozódó transzspirációs intenzitás

25
2.3. Vízmozgás a fatestben

26
A víz a központi hengerben a fatest, a xilém vízszállító csöveibe kerül, amelyek elhalt sejtek

27
Vízáramlás a trachea elemeken keresztül Poiseuille egyenlet: V = r4/8 ( P/ x) r= rádiusz, = viszkozitás Következményei: 1 érett metaxilém elem (r= 100 m) = 560 metaxilém elem (r=23 m) A gyökérben a radiális víztranszport a limitáló és nem az axiális (1 membrán barrier ellenállása = 24 km xilémedény ellenállásával Vízáramlás a trachea elemeken keresztül Poiseuille egyenlet: V = r4/8 ( P/ x) r= rádiusz, = viszkozitás Következményei: 1 érett metaxilém elem (r= 100 m) = 560 metaxilém elem (r=23 m) A gyökérben a radiális víztranszport a limitáló és nem az axiális (1 membrán barrier ellenállása = 24 km xilémedény ellenállásával
 r= rádiusz, = viszkozitás Következményei: 1 érett metaxilém elem (r= 100 m) = 560 metaxilém elem (r=23 m) A gyökérben a radiális víztranszport a limitáló és nem az axiális (1 membrán barrier ellenállása = 24 km xilémedény ellenállásával Vízáramlás a trachea elemeken keresztül Poiseuille egyenlet: V = r4/8 ( P/ x) r= rádiusz, = viszkozitás Következményei: 1 érett metaxilém elem (r= 100 m) = 560 metaxilém elem (r=23 m) A gyökérben a radiális víztranszport a limitáló és nem az axiális (1 membrán barrier ellenállása = 24 km xilémedény ellenállásával")
29
A kavitáció

30
3. A transzspiráció

31
Gázcserenyílások

32
Szőrök a levélfelszínen

33
A kutikula fluoreszcencia és fénymikroszkópos képe

34
Sztómazáró sejtek

35
Kis pórusokon keresztül intenzívebb a párologtatás, mint közeli vagy összefüggő felületeken

36
Levélszerkezet és sztóma elhelyezkedés

37
A gázcserét befolyásoló tényezők

38
A negatív nyomás kialakulásának helye és mechanizmusa

39
A hajtásba irányuló víztranszportot is a vízpotenciál gradiens tartja fenn

40
A hidraulikus lift

41
3.2. A sztómamozgás élettana és mechanizmusa

42
A gázcserenyílások környezeti tényezőktől függő reakciói

43
ZÁRT SZTÓMA NYITOTT SZTÓMA Fény Kis széndioxid koncentráció Átlagos hőmérséklet Elegendő víz Auxin (indolecetesav) Sötétség Nagy széndioxid koncentráció Szélsőséges hőmérséklet Vízhiány Abszcizinsav
 Sötétség Nagy széndioxid koncentráció Szélsőséges hőmérséklet Vízhiány Abszcizinsav")
44
A molekuláris mechanizmus

46
A zeaxantin lehetséges szerepe

47
A molekuláris mechanizmus TEA =tetraetil ammónium klorid

48
A molekuláris mechanizmus Ld. „Anyagtranszport és mechanizmusai” – később!

49
Kálium csatorna

50
A „patch clamp” technika

51
Ca 2+ VK SV FV K+K+ Vacuole Ca 2+ cyt K+K+ Ca 2+ -függő Ca 2+ -független H + cyt ABA IP 3 VmVm cADPR K+K+ K+K+ K in K out t Cytosol A sztómazárósejt működésének szabályozási módjai

52
Köszönöm a figyelmet
























 és azt.>")

