Előadást letölteni
1
Fejezetek a sejtbiológiából
Dr. Darvas Zsuzsa Dr. László Valéria Dr. Tóth Sára Genetikai, Sejt- és Immunbiológiai Intézet

2
Előadások hétfő , NET Barna terem 1. hét (Szept.6.) A sejten belüli transzport folyamatok fő típusai (Ea.: László V.) 2. hét (szept. 13.) Endoplazmás retikulum. A vezikuláris transzport 3. hét (szept. 20.) Szekréció és a Golgi 4. hét (szept. 27.) Az endocitózis. Az endoszómális – lizoszómális rendszer 5. hét (okt. 4.) A sejtmagmembrán, nukleáris lamina és a magpórus; kaputranszport (Ea.: Darvas Zs.) 6. hét (okt. 11.) A sejtmag egyes komponensei, ezek működése, ės azok hibái (Ea.: Tóth S.) 7. hét (okt. 18.) Az rRNS szintézis; a telomeráz 8. hét (okt. 25.) Az endoszimbionta eredetű sejtalkotók: A mitokondrium 9. hét nov. 1.) szünet 10. hét (nov. 8.) A peroxiszóma 11. hét (nov. 15.) A citoszkeleton: Mikrotubulusok 12. hét (nov. 22.) Mikrofilamentumok és intermedier filamentumok 13. hét (nov. 29.) A sejt – sejt, valamint a sejt – mátrix kapcsolódása 14. hét (dec. 6.) A sejtciklus szabályozása
 2. hét. (szept. 13.) Endoplazmás retikulum. A vezikuláris transzport. 3. hét. (szept. 20.) Szekréció és a Golgi. 4. hét. (szept. 27.) Az endocitózis. Az endoszómális – lizoszómális rendszer. 5. hét. (okt. 4.) A sejtmagmembrán, nukleáris lamina és a magpórus; kaputranszport. (Ea.: Darvas Zs.) 6. hét. (okt. 11.) A sejtmag egyes komponensei, ezek működése, ės azok hibái. (Ea.: Tóth S.) 7. hét. (okt. 18.) Az rRNS szintézis; a telomeráz. 8. hét. (okt. 25.) Az endoszimbionta eredetű sejtalkotók: A mitokondrium. 9. hét. nov. 1.) szünet. 10. hét. (nov. 8.) A peroxiszóma. 11. hét. (nov. 15.) A citoszkeleton: Mikrotubulusok. 12. hét. (nov. 22.) Mikrofilamentumok és intermedier filamentumok. 13. hét. (nov. 29.) A sejt – sejt, valamint a sejt – mátrix kapcsolódása. 14. hét. (dec. 6.) A sejtciklus szabályozása.")
3
Az eukarióta sejt eredete

6
Centrális dogma Információ hordozó (DNS vagy fehérje) végrehajtó
 végrehajtó")
7
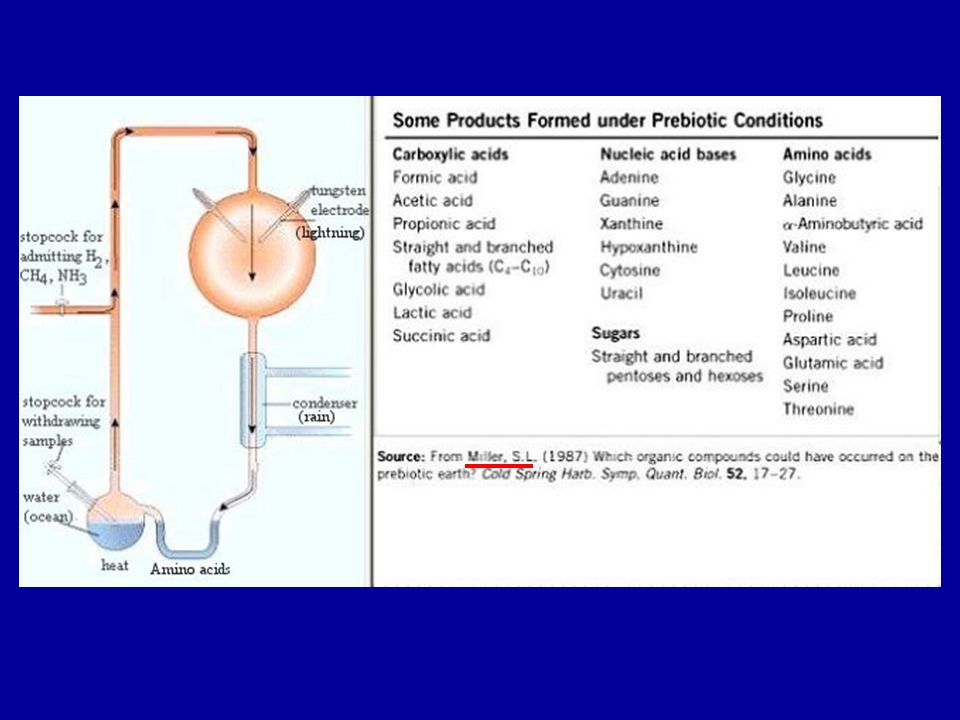
„RNS világ” Az RNS rendelkezik két fontos tulajdonsággal: templát
enzim (ribozimok) (splicing és transzláció)
 (splicing és transzláció)")
10
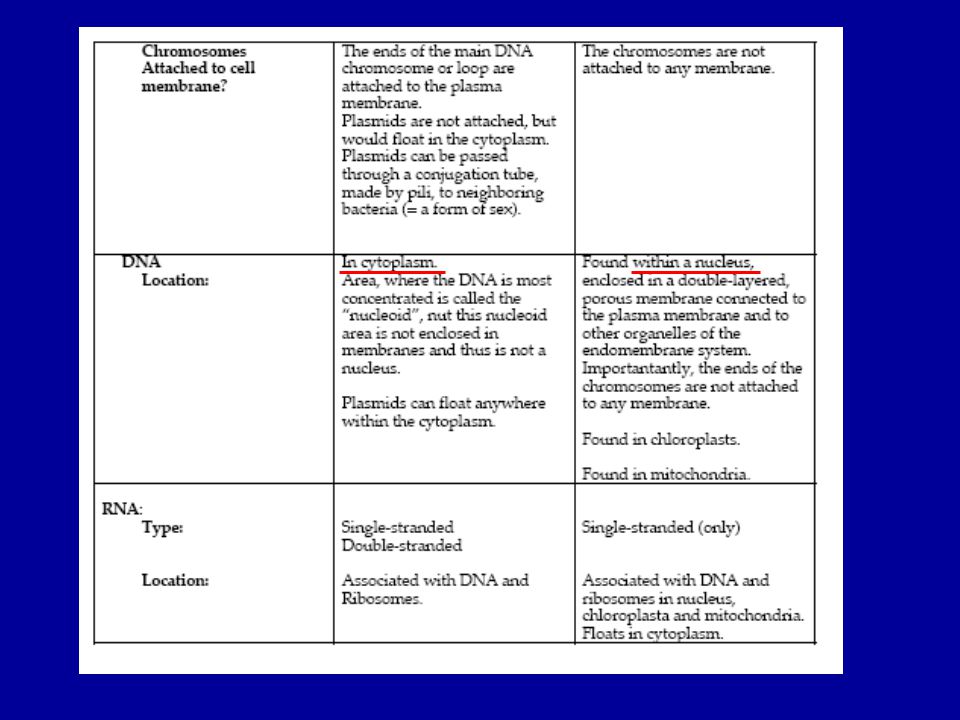
(EU)- (BACTERIA) Prokaryota
Archaea: cell membrane contains ether linkages; cell wall lacks peptidoglycan; genes and enzymes behave more like Eukaryotes; have three RNA polymerases like eukaryotes; and extremophiles Bacteria: cell membrane contains ester bonds; cell wall made of peptidoglycan; have only one RNA polymerase; react to antibiotics in a different way than archea do. (BACTERIA) Prokaryota Extrém körülmények között élnek (pl.Thermoacidophiles: Thermus aquaticus – PCR reakcióban a Taq polimerázt alkalmazzák)
 Prokaryota. Extrém körülmények között élnek (pl.Thermoacidophiles: Thermus aquaticus – PCR reakcióban a Taq polimerázt. alkalmazzák)")
11
2,7 milliárd éve 3,8 milliárd éve
Az eukarióták jobban „hasonlítanak” az archaeabaktériumokhoz, mint az eubaktériumokhoz.

13
Size (bázispár) ~ ~
 ~106 ~")
15
yes Cytoskeleton yes yes

16
Pro és eukarióta sejt szerkezete
A méretbeli különbség kb. 10-szeres!!

17
Az eukarióta sejt belső membránjainak eredete

18
A belső membránok invaginációval endoszimbiózissal keletkeztek.

19
Endoszimbionta elmélet
Lynn Margulis – 1966 1980 – bizonyítékok Anaerob eukarióták – nincs mitokondriumuk –főleg paraziták (Giardia intestinalis, Entamoeba histolytica, Microsporidia) Helyette mitosoma, hidrogenoszóma (dupla membrán, de nincs genom - degenerált mitokondrium)
 Helyette mitosoma, hidrogenoszóma (dupla membrán, de nincs genom - degenerált mitokondrium)")
20
(eu) A csillók eredete (endoszimbiózis) (pro)
 A csillók eredete (endoszimbiózis) (pro)")
21
(proteobaktérium)
")
22
Az eukarióta sejt belső tereinek topológiája

23
A sejten belüli fehérje és lipid
transzport mechanizmusok

24
Az eukarióta sejt fehérje (és foszfolipid)
transzportja

25
Alap kérdések a fehérje transzportoknál
Szignál Receptor Transzlokációs csatorna (ha membránon keresztül történik) energia
 energia.")
26
Fehérje szignál(A) és folt(B)=irányító szám
 és folt(B)=irányító szám")
27
Néhány szignál szekvencia
A szignál szekvencia általában nincsen rajta az érett fehérjén. Esetleg több szignál is kell (pl. mitokondrium) Szénhidrát is lehet szignál – M-6-PO4 (lizoszómális fehérjék)
 Szénhidrát is lehet szignál – M-6-PO4 (lizoszómális fehérjék)")
28
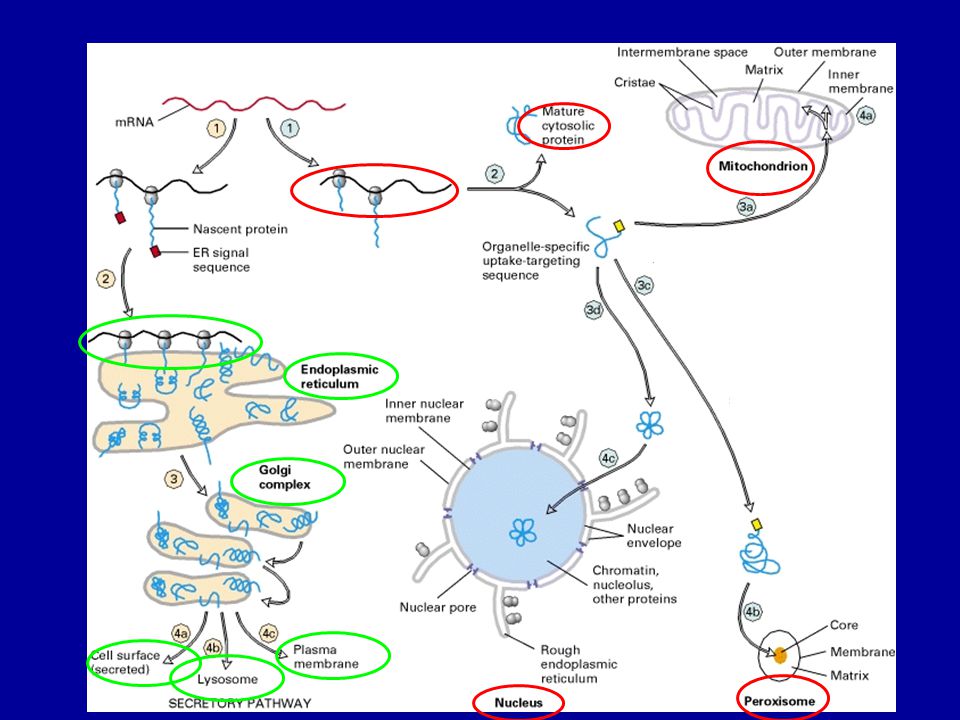
A fehérjék szintézise és szortírozása I.
szabad riboszóma (szintézis után) Sejtmag 1.Kapu transzport NLS szignál nélkül Citoszol
 Sejtmag. 1.Kapu transzport. NLS. szignál nélkül. Citoszol.")
29
A fehérjék szintézise és szortírozása II.
szabad riboszóma (szintézis után) Sejtmag 1.Kapu transzport NLS N-term. szignál nélkül C term. Mitokondrium Peroxiszóma 2. Poszttranszlációs transzmembrán transzport Citoszol
 Sejtmag. 1.Kapu transzport. NLS. N-term. szignál nélkül. C term. Mitokondrium. Peroxiszóma. 2. Poszttranszlációs. transzmembrán transzport. Citoszol.")
30
A fehérjék szintézise és szortírozása III.
SRP Membránhoz kötött riboszóma szabad riboszóma (szintézis közben) Endoplazmás retikulum 3.Ko-transzlációs transzmembrán transzport
 Endoplazmás. retikulum. 3.Ko-transzlációs. transzmembrán. transzport.")
31
A fehérjék szintézise és szortírozása IV.
Endoplazmás retikulum Szabad riboszóma (szintézis közben) 4. Vezikuláris transzport Golgi Transzport vezikulum Szekréciós vezikulum lizoszóma Reziduális test Plazmamembrán endoszóma
 4. Vezikuláris transzport. Golgi. Transzport. vezikulum. Szekréciós vezikulum. lizoszóma. Reziduális. test. Plazmamembrán. endoszóma.")
33
Kotranszlációs transzmembrán transzport (endoplazmás retikulum működése)
")
34
Szignál hipotézis Günter Blobel – Orvosi Nobel díj 1999 1971

35
A riboszómák a szintetizált fehérjétől függően szabadon
vagy membránhoz kötötten helyezkednek el Riboszóma alegységek a citoszolban Az SRP közreműködésével tudnak az ER membránhoz kötődni.

36
A szignál felismerő részecske (SRP)
Prokarióta SRP RNS Hidrofób oldalláncok – Szignál szekvencia megkötése P9, 14, 19, 54, 68, 72 - fehérjék 7S RNS (300 nukleotid) váz - nukleoluszban RNS polimeráz III, fehérjék részben a magban, részben a citoplazmában kapcsolódnak. Szignál: aminosav N-terminálisan (1+ majd hidrofób- szükséges és elégséges a transzlokációhoz)
 váz - nukleoluszban RNS polimeráz III, fehérjék részben. a magban, részben a citoplazmában kapcsolódnak. Szignál: aminosav N-terminálisan (1+ majd hidrofób- szükséges és elégséges. a transzlokációhoz)")
37
Monomer (kis) G-fehérjék
 G-fehérjék")
38
Bakteriális SRP és SRP receptor GTP kötés stabilizálja, a
GTP hidrolízise pedig destabilizálja az SRP és az SRP receptor kapcsolódását.

39
Kotranszlációs transzmembrán transzport

40
Transzmembrán protein transzport baktériumban
Intracellular Extracellular The chaperone protein SecB binds to the nascent polypeptide chain to prevent premature folding which would make transport across the plasma membrane impossible. SecE and SecY are transmembrane components which form a pore in the membrane through which the still unfolded polypeptide is threaded. The translocation process is energy-dependent (ATP) and is driven by SecA. Once the protein has passed through the pore, the signal sequence is cleaved off by an extracellular, membrane-bound protease.
 and is driven. by SecA. Once the protein has passed through the pore, the signal sequence is cleaved off by an extracellular, membrane-bound protease.")
41
Transzlokon (eukarióta)
Trimer - komponensei: Sec61 (10 membránt átérő helix) és Sec61 , Sec61 Hidrofób csatorna – fehérje számára átjárható, de a hidrofil ionok és molekuláknak nem A szintézis biztosítja az energiát Nyitott és zárt állapot (riboszóma felől) Oldalra is ki tud nyílni Szignál peptidáz – transzmembrán fehérje (a szignál szekvenciától C terminálisan található szekvenciát ismer fel)
 és Sec61 , Sec61 Hidrofób csatorna – fehérje számára átjárható, de a hidrofil ionok és molekuláknak nem. A szintézis biztosítja az energiát. Nyitott és zárt állapot (riboszóma felől) Oldalra is ki tud nyílni. Szignál peptidáz – transzmembrán fehérje (a szignál szekvenciától C terminálisan található szekvenciát ismer fel)")
42
(Poszttranszlációs transzmembrán transzlokáció az ER-on)
Gombákban általános (esetleg más eukariótákban is) BiP (Hsp70 csaperon) és ATP kell hozzá
 BiP (Hsp70 csaperon) és. ATP kell hozzá.")
43
Transzmembrán fehérjék elhelyezkedése
Az ER-ben, a kotranszlációs transzmembrán során alakul ki és a további transzport során már nem változik meg.

44
Type I: Signal sequence on amino terminus enters first and continues to elongate. Protein is threaded through the translocating channel (open area in rer membrane) until a hydrophobic stop sequence is reached. That hydrophobic stop sequence (seen as a hatched region in the protein) is then inserted in the membrane and forms the anchor for that protein. Signal is cleaved by protease inside the lumen.
 until a hydrophobic stop sequence is reached. That hydrophobic stop sequence (seen as a hatched region in the protein) is then inserted in the membrane and forms. the anchor for that protein. Signal is cleaved by protease inside the lumen.")
45
Type II: No cleavable signal sequence
Type II: No cleavable signal sequence. These proteins have rather long hydrophobic regions that will be anchored in the membrane. Type II proteins are threaded into the lumen with the C terminus leading. Protein continues to be inserted until it reaches the hydrophobic stop signal sequence.

46
What regulates the orientation of Type II and III proteins?

47
Membránt kétszer és többször átérő fehérjék

48
Mi történik a fehérjékkel az ER lumenében?
Cél: funkcióképes térszerkezet (harmadlagos, negyedleges) kialakítása Proteolízis (szignál peptidáz) Hajtogatás (PDI –protein diszulfid izomeráz, calnexin, calretikulin, BiP) N-glikoziláció (szintézis közben) Multimer protein összeszerelése Minőség ellenőrzés
 kialakítása. Proteolízis (szignál peptidáz) Hajtogatás (PDI –protein diszulfid izomeráz, calnexin, calretikulin, BiP) N-glikoziláció (szintézis közben) Multimer protein összeszerelése. Minőség ellenőrzés.")
49
N-glikoziláció N-oligoszacharid lánc N-oligoszacharid lánc módosítása
hozzákötése

50
Glikoziláció jelentősége
hajtogatódás (pl. hemagglutinin) Stabilitás (ECM fibronektin) Sejt adhézió (leukociták és endotél CAM-ja) Antigenitás (A,B,O vércsoportok)
 Stabilitás (ECM fibronektin) Sejt adhézió (leukociták és endotél CAM-ja) Antigenitás (A,B,O vércsoportok)")
51
A,B,0 vércsoport antigének
tonga.usip.edu/gmoyna/ biochem341/lecture29.html A,B,0 vércsoport antigének

52
Protein diszulfid izomeráz (PDI)
A PDI egyrészt létrehozza a diszulfid hidakat (a), részben a hibásakat felismeri és korrigálja (b).
, részben a hibásakat felismeri és korrigálja (b).")
53
ER lumen fő jellemzői: oxidatív magas Ca2+

54
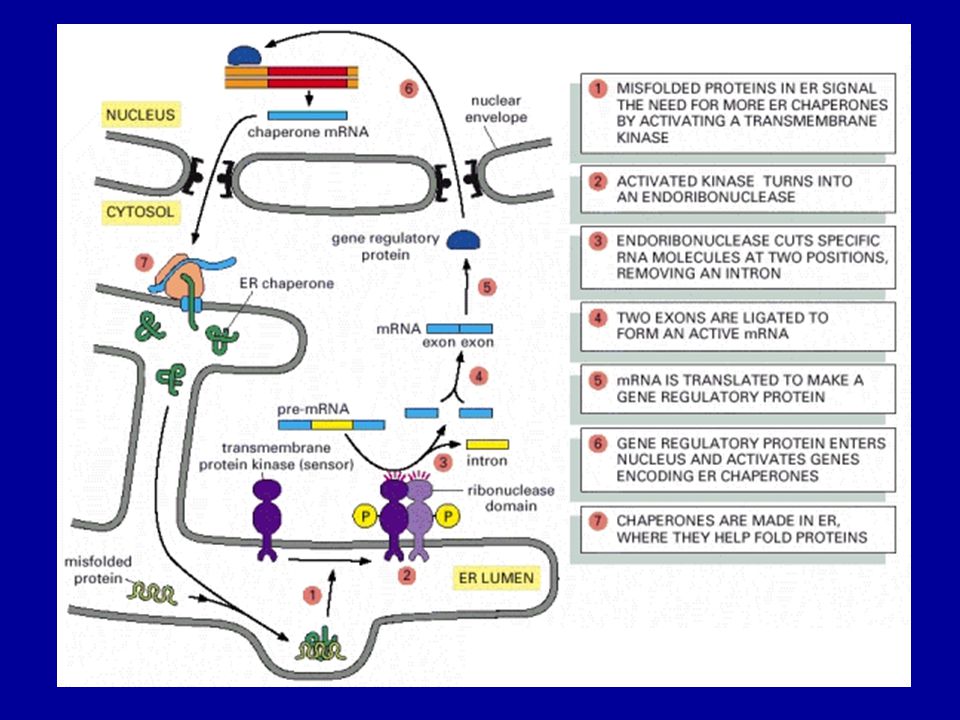
Unfolded protein response (UPR)
Csaperon expresszió A rosszul hajtogatott fehérjék indukálják a hajtogatást segítő csaperonok szintézisét.

56
Protein degradáció a proteaszómában

57
Multimer protein összeszerelése
Hemagglutinin

58
Sima felszínű endoplazmás retikulum

59
Sima felszínű endoplazmás retikulum funkciói
Általános membránlipidek (foszfolipidek koleszterin, ceramid) szintézise Ca ion tárolás Specifikus Szteroidok szintézise Méregtelenítés (máj) Glükóz metabolizmus
 szintézise. Ca ion tárolás. Specifikus. Szteroidok szintézise. Méregtelenítés (máj) Glükóz metabolizmus.")
60
SER szerepe a glukóz metabolizmusban
SER membrán

61
SER szerepe a glukóz metabolizmusban
SER membrán Glükóz 6 foszfatáz – glükoneogenesis and glükogenolízis utolsó lépése (máj, vese)
")
62
Glikogén tárolási betegség (GSD)
GSDIa – von Gierke – glükóz-6 foszfatáz (G6PC:17q21) hibája GSDIb – glükóz-6 foszfát transzporter (G6PT1:11q23) hibája GSDIc - pirofoszfát transzporter(11q23-q24.2) hibája-ritka GSDId?– glükóz transzporter hibája - nem ismert Súlyos tünetei: glikogén felhalmozódás a májban és a vesében Hipoglikémia, hepatomegália Lókusz heterogénia
 hibája. GSDIb – glükóz-6 foszfát transzporter (G6PT1:11q23) hibája. GSDIc - pirofoszfát transzporter(11q23-q24.2) hibája-ritka. GSDId – glükóz transzporter hibája - nem ismert. Súlyos tünetei: glikogén felhalmozódás a májban és a vesében Hipoglikémia, hepatomegália. Lókusz heterogénia.")













 prokariótában>")






>")



