Előadást letölteni
1
Kompetitív kizárás vagy együttélés?
Az interspecifikus kompetíció modelljei

2
Verhulst- féle logisztikus modell:
(Lotka-Volterra két fajos egyensúlyi modell) Egy faj esetén dN1/dt = rmax,1N1(K1-N1)/K1 Két faj esetén: dN1/dt = rmax,1N1[K1-(N1+α12N2)]/K1 dN2/dt = rmax,2N2[K2-(N2+α21N1)]/K2 Ahol, α12 az arányossági tényező azt mutatja, hogy a második faj egy egyede hányszorosra csökkenti a közös forrást az 1-es populáció egy egyedével arányosan
 Egy faj esetén. dN1/dt = rmax,1N1(K1-N1)/K1. Két faj esetén: dN1/dt = rmax,1N1[K1-(N1+α12N2)]/K1. dN2/dt = rmax,2N2[K2-(N2+α21N1)]/K2. Ahol, α12 az arányossági tényező azt mutatja, hogy a második faj egy egyede hányszorosra csökkenti a közös forrást az 1-es populáció egy egyedével arányosan.")
3
Verhulst- féle logisztikus modell:
zérus növekedési görbe két populáció esetén: rmax,1N1[K1-(N1+α12N2)]/K1=0 Mivel az rmax,1N1/K1=0 [K1-(N1+α12N2)]=0 Egy egyenes egyenlete az (N1, N2) koordináták esetén: N2= -N1/α12+K1/α12 Egyenlet a 2-es populációra: N2= -α12/N1+K2
]/K1=0. Mivel az rmax,1N1/K1=0. [K1-(N1+α12N2)]=0. Egy egyenes egyenlete az (N1, N2) koordináták esetén: N2= -N1/α12+K1/α12. Egyenlet a 2-es populációra: N2= -α12/N1+K2.")
4
Verhulst- féle logisztikus modell: egy faj növekedési görbéje
dN2/dt=0 K2/α21 N1

5
Verhulst- féle logisztikus modell: két faj zéró növekedési görbéje
● K2/α21 K1 N1 A kezdeti állapotól függően az egyik faj kihal.

6
Verhulst- féle logisztikus modell:
dN1/dt=0 N2 dN2/dt=0 N1

7
Verhulst- féle logisztikus modell: két faj zéró növekedési görbéje
● K1 K2/α21 N1 A két faj stabil együttélése

8
Verhulst- féle logisztikus modell:
K2>K1/α12; K2>K2/α21 “complete competitors cannot coexist” (Hardin, 1960) K2<K1/α12; K1<K2/α21 α12 α21 < 1 A kompetticó határt szab az együtt élő fajok hasonlóságának és számának. Elérhető tápanyag mennyisége, illetve annak tér- és időbeli eloszlása. K- és R- stratégista fajok
 K2<K1/α12; K1<K2/α21. α12 α21 < 1. A kompetticó határt szab az együtt élő fajok hasonlóságának és számának. Elérhető tápanyag mennyisége, illetve annak tér- és időbeli eloszlása. K- és R- stratégista fajok.")
9
Tilman (1982, 1986) forráshasználati modellje
 forráshasználati modellje")
10
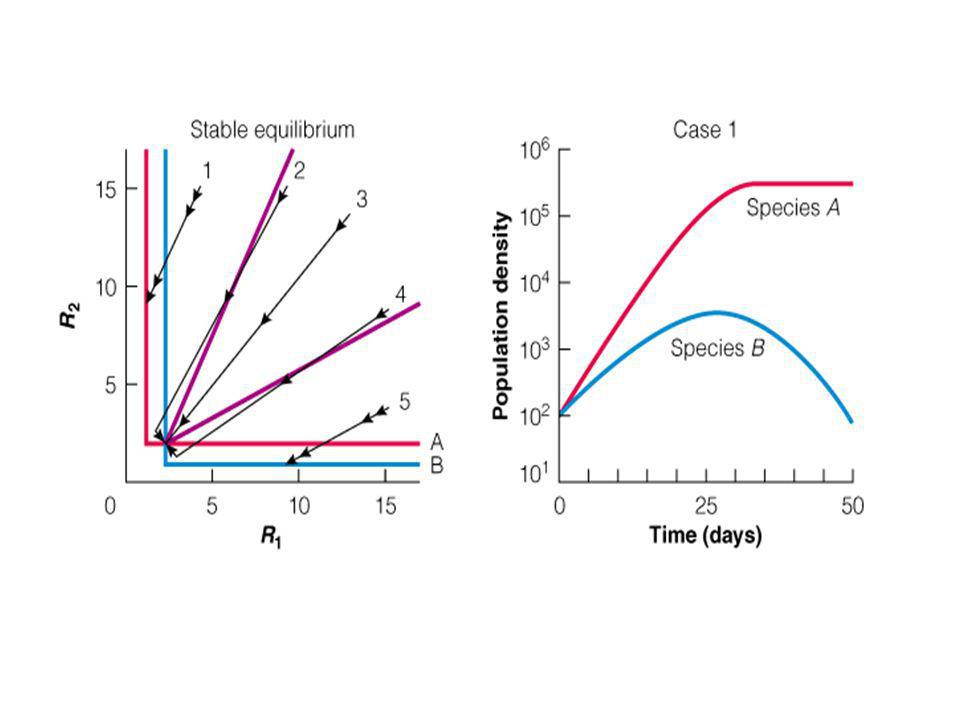
1 – nincs túlélő faj 2 – csak az “A” él túl 3 – az “A” kizárja a “B”-t. 4 – Stabil együttélés 5 – az “B” kizárja a “A”-t. 6 – csak az “B” él túl

12
Gazda-parazitoid kapcsolat
Paraziták: nem pusztítják el a gazdát közvetlenül. Parazitoid:Lárváik egy másik rovarban fejlődnek, mely legtöbbször annak pusztulásához vezet – hymenopterák (Trichomonidae, Mymariade) Nem tekinthetők ragadozóknak: Az áldozat testét az utódoknak juttatja. A parazitált gazdát újból meg lehet támadni.
 Nem tekinthetők ragadozóknak: Az áldozat testét az utódoknak juttatja. A parazitált gazdát újból meg lehet támadni.")
13
Nicholson-Bailey modell
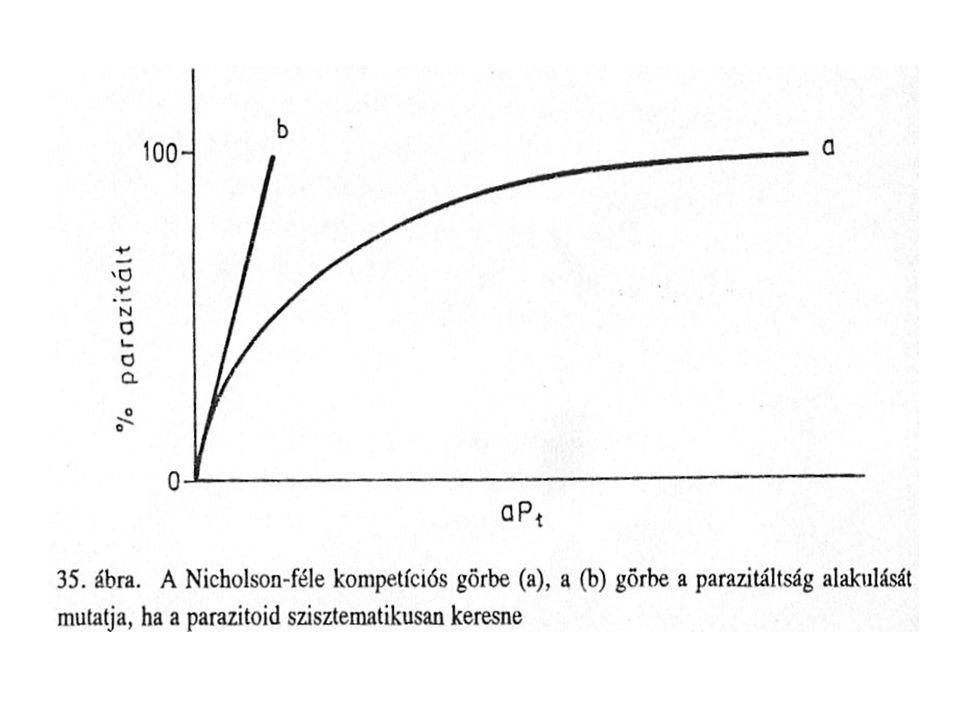
Feltételek: Gazda-parazitoid találkozások száma időegység alatt (Ne) a két populáció egyedsűrűségének (NtPt) függvénye. Ne= aNtPt Ahol „a” a parazitoid hatékonysága: -„area of discovery” -egy fajta nem telítődő funkcionális válasz (meredekségét a bejárt terület nagysága határozza meg) gazdával való találkozás véletlenszerű
 a két populáció egyedsűrűségének (NtPt) függvénye. Ne= aNtPt. Ahol „a a parazitoid hatékonysága: -„area of discovery -egy fajta nem telítődő funkcionális válasz (meredekségét a bejárt terület nagysága határozza meg) gazdával való találkozás véletlenszerű.")
14
A gazda – parazitoid találkozás valószínűségét a Poisson eloszlás adja meg:
Pk=λkeλ/k! Ahol (λ) a találkozások számának és a gazdaszámnak a hányadosa: λ =Ne/Nt A már parazitált gazdák kezdetben életben maradnak és a modell szerint megkülönböztethetetlenek.
 a találkozások számának és a gazdaszámnak a hányadosa: λ =Ne/Nt. A már parazitált gazdák kezdetben életben maradnak és a modell szerint megkülönböztethetetlenek.")
15
0-szor való találkozás valószínűsége:
P0= exp( -Ne/Nt) 1 parazitoiddal való találkozás valószínűsége: Pa= 1- exp( -Ne/Nt) Legalább egyszer parazitált állat: Na= paNt = Nt[1- exp( -Ne/Nt)] Mivel Ne/Nt = aPt Na= Nt[1- exp( - aPt)]
 1 parazitoiddal való találkozás valószínűsége: Pa= 1- exp( -Ne/Nt) Legalább egyszer parazitált állat: Na= paNt = Nt[1- exp( -Ne/Nt)] Mivel Ne/Nt = aPt. Na= Nt[1- exp( - aPt)]")
17
A véletlenszerű kereső parazitoid populáció nagyságával arányosan nő annak valószínűsége, hogy egy már parazitált gazdával találkoznak. Egy gazda-egy parazita a modellben: Pt+1= Na Tehát, a parazitált gazdák száma a következő generációbeli parazitoidok számát adja meg. A tovább szaporodni képes gazdákra érvényes: Nap0= Nt[exp(-aPt)]
]")
18
Gazda- és parazitoid populáció növekedését leíró differencia-egyenlet rendszer, ahol γ a gazdapopuláció növekedési rátája: Nt+1= γNtexp(-aPt) Pt+1= Nt[1- exp(-aPt)] Nt+1= Nt ; Pt+1= Pt N* = γlnγ/[(γ-1)]; P* = lnγ/a, instabil egyensúlyt képvisel, kimozdítva bármelyik populációt a rendszer divergens oszcillációba kezd.
 Pt+1= Nt[1- exp(-aPt)] Nt+1= Nt ; Pt+1= Pt. N* = γlnγ/[(γ-1)]; P* = lnγ/a, instabil egyensúlyt képvisel, kimozdítva bármelyik populációt a rendszer divergens oszcillációba kezd.")
19
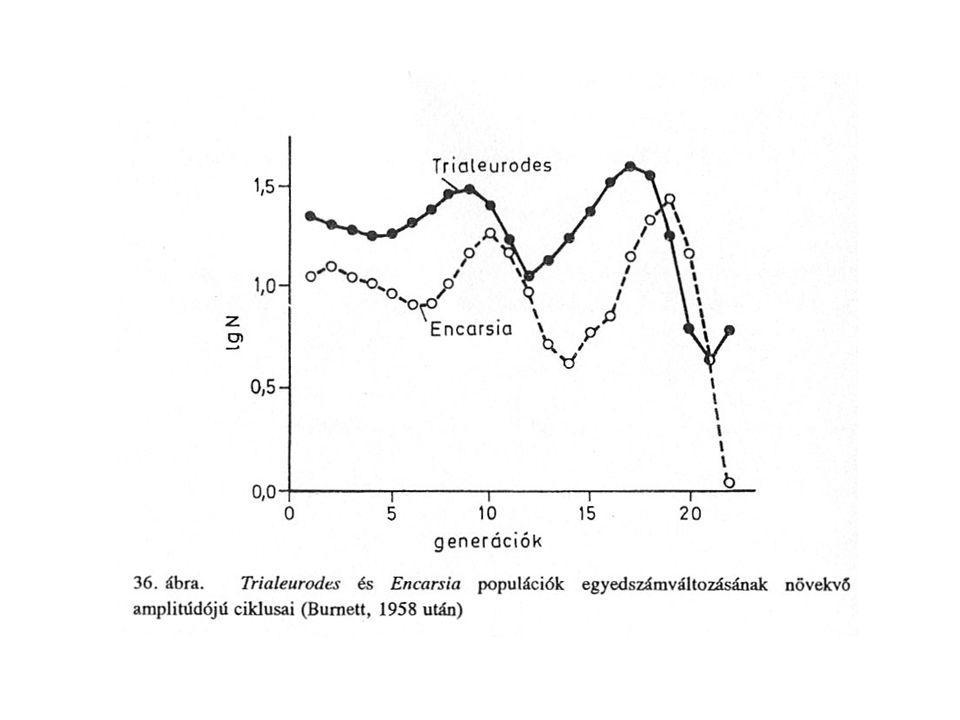
(Burnett, 1958) Trialeurodes vaporiorum, Encarsia formosa
A generációkat úgy hozta létre, hogy a parazitoidok számát az előző generációban parazitált gazdák számából Gazdákét a nem parazitált gazdák számának kétszeresére emelte Eredmény instabil oszcilláció azonban stabilitása nagyobb volt, mint amit a modellből számítottak.

21
A Nicholson-Bailey modell eredetileg nem tartalmaz negatív visszacsatolást, ezáltal szabályozó hatás nem érvényesül, A modell alkalmazhatóságát korlátolja, hogy ebben a formában természetes populációk jellemzésére nem alkalmas. Free & Lawton ( 1975): szabályozás megjelenik a modellben Nt+1= Ntγexp[(1-Nt/K) -aPt] Pt+1= Nt[1-exp(-aPt)] Ahol a negatív visszacsatolást a gazda limitáltsága okozza
: szabályozás megjelenik a modellben. Nt+1= Ntγexp[(1-Nt/K) -aPt] Pt+1= Nt[1-exp(-aPt)] Ahol a negatív visszacsatolást a gazda limitáltsága okozza.")
22
Előreláthatalan rések, fluktuáló környezet
Rések keletkezhetnek valós és “ökológiai térben” egyaránt (tüzek, földcsuszamlás, nyíladék az erdőben stb.) “Fugitive species” Fluktuáló környezet (Hutchinson, 1961) Alga paradoxon kompetitív egyensúly eltolódás,
 Fugitive species Fluktuáló környezet (Hutchinson, 1961) Alga paradoxon. kompetitív egyensúly eltolódás,")
23
Megjósolhatatlan rések, hiányfoltok: a gyengébb kompetítor a jobb kolonizáló
Postelsia palmaeformis Mytilus californianus Hullámverés, kőgörgetés Átl. résarány ≥ 7% -P. palmaeformis megtelepszik

24
Megjósolhatatlan rések, hiányfoltok: “elővásárolt tér”
“first come, best served” California, USA Bromus madritensis, B. rigidus Kevert tenyészet- egyenlőarány B. rigidus több fitomassza, kizárása a B. madritensis-nek kedvez Inherens kompetitív tulajdonságok?....Nem! Fluktuáló környezet

25
Ideiglenes foltok, megjósolhatalan élettartam
“A jó és a gyors együttélése” Kiszáradó mocsárétek, Indiana, USA Physa gyrina, korai reprodukció Lymnea elodes, erősebb kompetítor Közös tenyészet L. elodes sikeres, ha egész nyáron át fennáll az interakció

26
Aggregált eloszlás A tömeges erős kompetítor denzitásfüggése révén réseket nagyhat a gyengébb kompetítornak Inta > Inter = aggregált eloszlás (- binomiális) Döglegyek, Finnország 5m2 foltok Telepítés: 9faj/folt Átlagos fajszám: 2.7/folt Territórium fenntartása több foltból “Gyepautomata”- Agrostis stolonifera, Cynosurus cristatus, Holcus lanatus, Lolium perenne, Poa trivialis Rendezett parcella- Lolium aránya 60% Random elrendezés – “a szomszédok döntenek”
 Döglegyek, Finnország. 5m2 foltok. Telepítés: 9faj/folt. Átlagos fajszám: 2.7/folt. Territórium fenntartása több foltból. Gyepautomata - Agrostis stolonifera, Cynosurus cristatus, Holcus lanatus, Lolium perenne, Poa trivialis. Rendezett parcella- Lolium aránya 60% Random elrendezés – a szomszédok döntenek")
27
Mi van? Általánosságban a heterogenitás (a környezetben) stabilizál.
 stabilizál.")
28
Látszólagos versengés: ellenségmentes környezet
Santa Catalina Island, California Prédák: Tegula aureotincta, T. eiseni Astrea undosa-csigák; Chama arcana-kagyló Ragadozók: Panulirus interruptus, Octopus bimaculatus, Kelletia kelleti Magas tagoltság- kagylók, sok ragadozó Alacsony tagoltság- csigák, kevés ragadozó Artemisia bozótos, Arizona diverzitás vs. menedék

29
Az interspecifikus kompetíció evolúciós hatásai
Kompetítiv szétterjedés Új Guinea- Gyümölcsgalambok Chalcophaps indica - parti élőhelyek C. stefani - belső (másodlagos) erdők Gallicolumba rufigula - belső esőerdők Más szigetek (ahol nincs G. rufigula) szomszédos élőhelyen élő faj kolonizálja a másik faj élőhelyét is
 erdők. Gallicolumba rufigula - belső esőerdők. Más szigetek (ahol nincs G. rufigula) szomszédos élőhelyen élő faj kolonizálja a másik faj élőhelyét is.")
30
Karakter eltolódás A kompetíció bizonyitéka melyet a niche expanzió eredményez a kompetítor hiányából adódóan (kompetitiv terjedés) a fajok realizált niche-ben jelentkező különbségek szim-, illetve allopatrikus populációk esetén. Ha ezek a változások morfológiai változásokkal járnak - karakter eltolódás

31
Veromessor pergandei Davidson, 1978 A mandibula variácó ott nagyobb, ahol nagyobb az interspecifikus kompetíció mértéke Sitta tephronota, S. neumayer Area átfedés Iránban- különböző csőrhossz

32
Dánia, Finnország Hydrobia ulvae – ár-apály zóna, H. ventrosa - lagunákban Allopatrikus populációk hasonló méretűek Szimpatrikus populációkban a H. ulvae a nagyobb Az H. ulvae nem árapály zóna élőhelyein nagyobbra nő

33
Korlátozott hasonlóság (“limiting similarity”)
Milyen nagyságú kell, hogy legyen a niche elkülönülés, hogy stabil együttélés alakuljon ki két populáció között ? Készlethasznosítási görbe (May, 1963) α =e-d2/4/w2 , ahol “d” a csúcsok közötti távolság, “w” a relatív niche szélesség (S.D.) d/w<1 - finom egyensúly 1<d/w <3 - stabil és instabil egyensúly d/w >3 – stabil koegzisztencia Optimális hasonlóság (“optimal similarity”)
 α =e-d2/4/w2. , ahol d a csúcsok közötti távolság, w a relatív niche szélesség (S.D.) d/w<1 - finom egyensúly. 1<d/w <3 - stabil és instabil egyensúly. d/w >3 – stabil koegzisztencia. Optimális hasonlóság ( optimal similarity )")
34
Irodalom Szentesi Árpád, Török János, Állatökológia. Kovásznai kiadó, Budapest. Begon, M., Harper, J.L., Townsend, C.R.,1996. Ecology. third edition, Blackwell Science Ltd. London, UK Kot, M Elements of Mathematical Ecology. Cambridge University Press, Cambridge, UK Kozár, F., Samu, F., Jermy, T., Az állatok populáció dinamikája. Akadémiai kiadó, Budapest.

35
A predáció Egy élőlény (a préda) elfogyasztása egy másik élőlény (a ragadozó) által. A préda még él mikor a ragadozó először megtámadja. A ragadozók osztályozása: Taxonómiai: húsevők állatokat a növényevők növényeket, a mindenevők (majdnem) mindent fogyasztanak. Funkcionális: valódi ragadozók,”legelők”, parazitoidok és paraziták
 mindent fogyasztanak. Funkcionális: valódi ragadozók, legelők , parazitoidok és paraziták.")
36
Valódi ragadozók: „klasszikus ragadozás” a fogalom szerint
Valódi ragadozók: „klasszikus ragadozás” a fogalom szerint. A préda teljes v. részleges elfogyasztását jelenti, nemcsak húsevők! (magevő kisemlősök planktonfogyasztó bálnák is stb.) Legelők(!)(portyázók?): A préda többnyire részleges elfogyasztását jelenti, hatásuk, káros, olykor letális a prédára nézve rövid távon, de megjósolhatóan nem lethálisak (ebben az esetben valódi ragadozók lennének) pl.: növényevők, pióca, vér szívó rovarok. Paraziták: A préda többnyire részleges elfogyasztását jelenti nem pusztítják el a gazdát közvetlenül. Kevés egyedet támadnak meg, vagy egy konkrét csoportot (vírusok, galandférgek, bélférgek stb.) Parazitoid: Lárváik egy másik rovarban fejlődnek, mely legtöbbször annak pusztulásához vezet – hymenopterák (Trichomonidae, Mymariade)
 Legelők(!)(portyázók ): A préda többnyire részleges elfogyasztását jelenti, hatásuk, káros, olykor letális a prédára nézve rövid távon, de megjósolhatóan nem lethálisak (ebben az esetben valódi ragadozók lennének) pl.: növényevők, pióca, vér szívó rovarok. Paraziták: A préda többnyire részleges elfogyasztását jelenti nem pusztítják el a gazdát közvetlenül. Kevés egyedet támadnak meg, vagy egy konkrét csoportot (vírusok, galandférgek, bélférgek stb.) Parazitoid: Lárváik egy másik rovarban fejlődnek, mely legtöbbször annak pusztulásához vezet – hymenopterák (Trichomonidae, Mymariade)")
37
A predátor és “viselkedése”

38
A predáció legtöbbször a préda populáció demográfiailag érdektelen fenológiai stádiumában történik.
A predáció mérsékelheti az intraspecifikus kompetcíciót( Columba palumbus). Legtöbbször az intraspecifikus kompetícióhoz kellően magas populáció denzitás váltja ki a ragadozást, ami annak mérsékléséhez vezet.
. Legtöbbször az intraspecifikus kompetícióhoz kellően magas populáció denzitás váltja ki a ragadozást, ami annak mérsékléséhez vezet.")
39
A predátor legtöbbször a leggyengébb egyedet támadja meg, melyek általában a fiatalok vagy az öregek: A fiatal állatokat könnyű elkapni Kisebb állóképesség, lassabb futás Nem elég jók még a ragadozó elkerülésében- tapasztalatlanok Sokszor hibáznak a ragadozó felismerése során

40
Hasonló jelenség a hebrivória esetén is előfordul:
Ausztrália: Perga affinis csak azokat az Eucalyptus fákat támadja meg, melyek vagy gyökérkárt szenvedtek vagy nagyon rossz minőségű élőhelyen élnek (mindezt okozhatja a vízelvezetés is).
.")
41
A fogyasztás hatása a fogyasztókra
A táplálék térbeli, időbeli eloszlása A ragadozók funkcionális válaszától A funkcionális válasz limitált a generációs idő által. A táplálék felvétele nagyban függ annak minőségétől, mennyiségétől és megfelelő formájától (medence-effektus).
.")
42
A ragadozók viselkedése
Monofágok Oligofágok Polifágok – generalisták specialisták

43
Táplálék preferencia A táplálék preferencia egy fajta összehasonlító étrend kombinálva a táplálék hozzáférhetőségével. Rangsoron alapuló preferencia: amikor a az egyszerűen skálázható források vannak dominálnak. ( pl.: legtöbb húsevő) Kiegyensúlyozott preferencia: integrált részét képezi egy étrendnek A két típus azonban keveredhet is
 Kiegyensúlyozott preferencia: integrált részét képezi egy étrendnek. A két típus azonban keveredhet is.")
44
Prédaváltás (“switching”)
Szorosan köthető a preferenciához Guppi (Tubifex, Daphnia) Általában a prédadenzitás változása indukálja, felelős az aggregált, illetve az ideálisan szabad eloszlások kialakulásáért Stabilizáló hatású
 Általában a prédadenzitás változása indukálja, felelős az aggregált, illetve az ideálisan szabad eloszlások kialakulásáért. Stabilizáló hatású.")
45
Kapcsoltság A kapcsoltság magába foglalja egy szokványos forráshoz kapcsolódó preferenciát: Minimum létszám küszöb ami alatt nincs keresési viselkedés. Az orientáció megnövekedett valószínűsége egy préda iránt, pl.: a fogyasztó kifejleszt egy keresőképet a forrásról és erre „koncentrál”, és ebbe a képbe illeszti bele a megfelelő prédát. Megnő az üldözés valószínűsége Megnő a préda elkapásának valószínűsége Tehát megnő az adott préda elejtési valószínűsége

46
Étrend-szélességi modell (MacArthur and Pianka, 1966, Charnov, 1976)
Ei/hi≥E/(s+h) Ahol, Ei – energia tartalom hi – ráfordított idő Ei/hi – a felkínált préda átlagos profitabilitása-”tényleges nyereség” s(i) – az „i” prédára fordított keresési idő várt input energia szint
 Ahol, Ei – energia tartalom. hi – ráfordított idő. Ei/hi – a felkínált préda átlagos profitabilitása- tényleges nyereség s(i) – az „i prédára fordított keresési idő. várt input energia szint.")
47
A kereső generalista, akkor Ei/hi értéke nagy, illetve E/(s+h) értéke kicsi a rendelkezésre álló könnyen megszerezhető táplálék miatt. A kereső specialista, akkor Ei/hi értéke hasonló a E/(s+h)-hoz, az „s” értéke kicsi. Tehát csak a legnyereségesebb prédát fogyasztja el a ragadozó (oroszlánok – sánta és öreg állatok fogyasztása). Tehát a modellből kizárhatók a nem-profitábilis prédaállatok.
-hoz, az „s értéke kicsi. Tehát csak a legnyereségesebb prédát fogyasztja el a ragadozó (oroszlánok – sánta és öreg állatok fogyasztása). Tehát a modellből kizárhatók a nem-profitábilis prédaállatok.")
48
A fogyasztó denzitásának hatása
Mutuális interferencia – a ragadozó részt vesz a zsákmánypopuláció szabályozásában, de ő is szabályozott a saját denzitása által – csatolt osszcilláció. A táplálék térbeli heterogenitása – hosszabb táplálkozási idő. Szociális viselkedés a ragadozó egyedek között növeli a hatékonyságot (falka). A mutuális interferencia stabilizálja a ragadozó-préda populációt (modellt).
. A mutuális interferencia stabilizálja a ragadozó-préda populációt (modellt).")
49
Fogyasztók és táplálék foltok
A ragadozók a általában legprofitábilisabb foltban töltik a legtöbb időt. A legtöbb ragadozó itt ebben a foltban várható A préda azokban az alacsony denzitású foltokban a legértékesebb, amelyek viszonylag védettek és a legvalószínűbb, hogy túlélnek.

50
Phyllotreta cruciferae - a legtömegesebb a nagy denzitású Brassica Foltokban - direkt denzitás függés Pieris rapae - a legtömegesebb az alacsony denzitású Brassica foltokban - inverz denzitás függés Phyllotreta atriolata - a közepes denzitású Brassica foltokban a legtömegesebb - közepes denzitás függés („domed” relationship)
")
51
Az optimális gyűjtés-megközelítés folthasználat esetén
A határérték teória (Charnov, 1976; Parker and Stuart, 1976): az az optimális időtartam a foltban amely annak elhagyásáig kinyert energia mennyiséggel arányos („legrövidebb idő – legtöbb energia”) Függ a folt minőségétől Viszont a modell nem számol a foltok közötti vándorlási idővel.
: az az optimális időtartam a foltban amely annak elhagyásáig kinyert energia mennyiséggel arányos („legrövidebb idő – legtöbb energia ) Függ a folt minőségétől. Viszont a modell nem számol a foltok közötti vándorlási idővel.")
52
Ollason-féle mechanisztikus modell:
Emlék modell A friss emlékek képződnek, mikor az állat táplálkozik. De az emlékek folyamatosan törlődnek. A beviteli szint függ az állat táplálkozási hatékonyságától és a terület produktivitásától. Az felejtési szint függ az állat azon képességétől, hogy felidézze emlékeit.

53
Ideálisan szabad eloszlás és aggregáció és interferencia
Az ideálisan szabad eloszlás egyensúly a vonzó és a taszító erők között. Sutherland, (1983) szerint a ragadozók elterjedése (pi) az „i” területen függ a területen található prédák (hi) arányától: pi=k(hi1/m) Ahol, m = az interferenciális koefficines, k = normalizáló konstans m=0 – nincs interferencia; 0<m<1 – alig meglévő interferencia. m ≈ 1 a ragadozók aránya nő a prédák arányának megfelelően. m>1 a legnagyobb denzitású préda populációk legalacsonyabb rendű ragadozóval rendelkezik. m=3 a ragadozók direkt denzitás függés szerint viselkednek.
 szerint a ragadozók elterjedése (pi) az „i területen függ a területen található prédák (hi) arányától: pi=k(hi1/m) Ahol, m = az interferenciális koefficines, k = normalizáló konstans. m=0 – nincs interferencia; 0<m<1 – alig meglévő interferencia. m ≈ 1 a ragadozók aránya nő a prédák arányának megfelelően. m>1 a legnagyobb denzitású préda populációk legalacsonyabb rendű ragadozóval rendelkezik. m=3 a ragadozók direkt denzitás függés szerint viselkednek.")
54
A ragadozók funkcionális válasza
A ragadozók táplálkozási viselkedése is hatással van ragadozó-zsákmány-rendszerekre. A ragadozók táplálkozási viselkedésének és az elfogyasztott zsákmány mennyiségének a zsákmánypopuláció egyedsűrűségétől való függését funkcionális válasznak nevezzük. Grafikus megjelenítés: a ragadozó által egységnyi idő alatt elejtett zsákmány mennyisége. Holling (1959) három fő típust különített el. Részletek később.....
 három fő típust különített el. Részletek később.....")
55
A predátor hatása a préda populációra
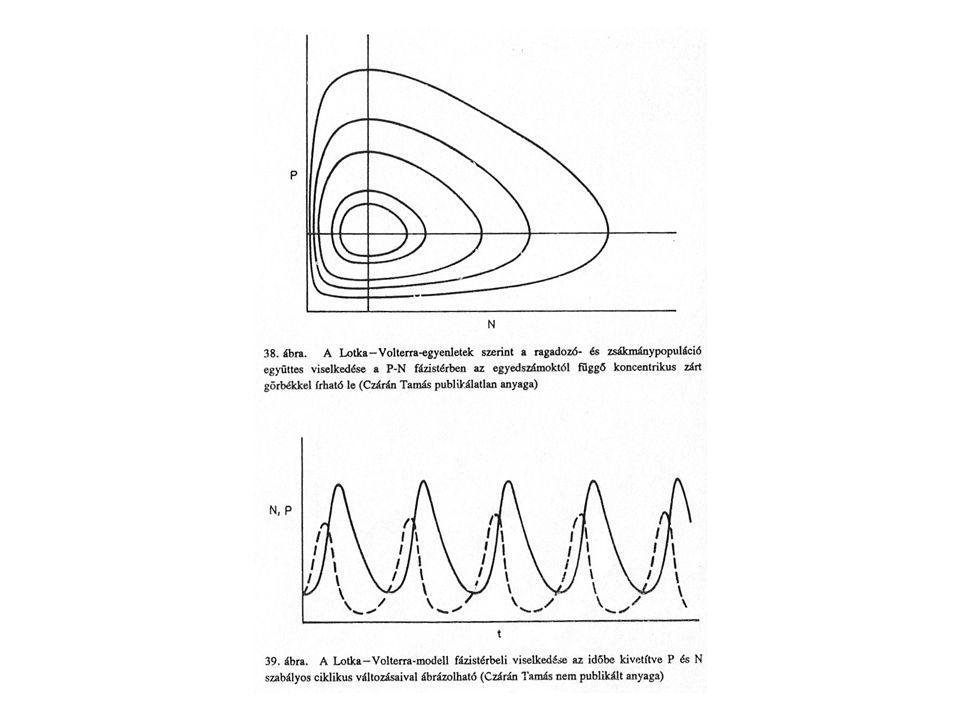
A ragadozó-préda rendszerek populációdinamikája – a Lotka-Volterra-modell

56
A Lotka-Volterra-modell
Feltételei: 1. A környezet állandó 2. A fajok egyedsűrűsége megfelelően reprezentálható egyetlen változóval, elhanyagolhatjuk a különböző fenotípusok és nemek közötti különbséget. 3. Az interakciók hatása azonnali: a táplálék elfogyasztása és anyagának a ragadozó testébe építése közti időt elhanyagolhatjuk. 4. A ragadozás a ragadozók és a zsákmány állatok számának szorzatával arányos. Ez a feltételrendszer csak akkor lehet igaz, ha a zsákmány random módon mozog, és a találkozások alkalmával (melyek gyakoriságát az”NP” szorzat jellemzi) állandó valószínűséggel következik be a zsákmányszerzés.
 állandó valószínűséggel következik be a zsákmányszerzés.")
57
Ragadozó hiányában a zsákmánypopuláció növekedését leíró egyenlet:
dN/dt= rmaxN A ragadozó jelenlétében a zsákmányolás arányos a ragadozó (P) és a zsákmánypopuláció nagyságával (N), amit a ragadozók keresési vagy támadási hatékonysága (a) befolyásol. Az exponenciális növekedést a zsákmányul ejtett egyedek száma csökkenti: dN/dt= rmaxN-aPN A ragadozó populációt vizsgálva egy kezdeti populációnagyságot feltételezve, préda hiányában exponenciális csökkenést várhatunk: dP/dt= -qP Ahol, (q) a mortalitási ráta. Ha zsákmány is van jelen, a ragadozó populáció növekedése a zsákmányejtések számától (aPN) és a zsákmány felhasználását megadó konstanstól (f ) függ: dP/dt=faPN-qP
 és a zsákmánypopuláció nagyságával (N), amit a ragadozók keresési vagy támadási hatékonysága (a) befolyásol. Az exponenciális növekedést a zsákmányul ejtett egyedek száma csökkenti: dN/dt= rmaxN-aPN. A ragadozó populációt vizsgálva egy kezdeti populációnagyságot feltételezve, préda hiányában exponenciális csökkenést várhatunk: dP/dt= -qP. Ahol, (q) a mortalitási ráta. Ha zsákmány is van jelen, a ragadozó populáció növekedése a zsákmányejtések számától (aPN) és a zsákmány felhasználását megadó konstanstól (f ) függ: dP/dt=faPN-qP.")
60
A zsákmány önkorlátozó logisztikus növekedést követ:
dN/dt= rmaxN-(r/K)N2-aPN A két populáció zéró növekedési görbéje: rmax-(rmax/K)N-aP=0 -q+faN=0 A zsákmány önkorlátozó viselkedése (egyedsűrűségfüggő növekedése) a modellben stabil egyensúlyra vezet.
N2-aPN. A két populáció zéró növekedési görbéje: rmax-(rmax/K)N-aP=0. -q+faN=0. A zsákmány önkorlátozó viselkedése (egyedsűrűségfüggő növekedése) a modellben stabil egyensúlyra vezet.")
61
A ragadozó önlimitáltsága, illetve zsákmánytól függő való egyedsűrűség-függés:
dN/dt=rmaxN-(1-N/K)-aPN, dP/dt= faP-bP2/N Stabil egyensúly gyorsabb elérése
-aPN, dP/dt= faP-bP2/N. Stabil egyensúly gyorsabb elérése.")
64
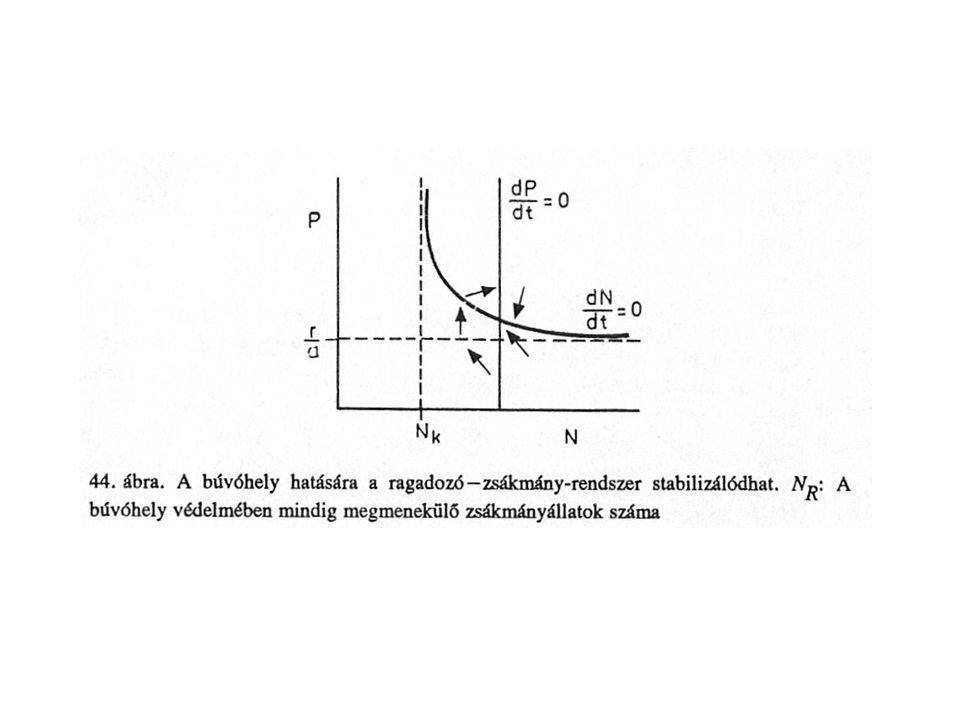
Az újratermelődő zsákmány, ha N>q/(fa) végtelen számú ragadozót eltarthat (A, B)
Az egyre erősebb önkorlátozás a stabilitást és a rezilienciát fokozza, növekszik azonban a zsákmányszint amelynél az egyensúly megvalósul (C, D). Stabilitást növelő tényező, ha a zsákmány egy része (NR) a ragadozók elől elmenekül (pl.: búvóhelyek révén). Így a ragadozók csak N-NR zsákmányhoz juthatnak. Az elérhetetlen zsákmány lehet: N állandó hányada: NR=cN, ez ekvivalens azzal ha az a és q konstansokat a(1-c) és q(1-c)-re változtatnák, mely nem befolyásolja a modellt NR az N-től független.
. Stabilitást növelő tényező, ha a zsákmány egy része (NR) a ragadozók elől elmenekül (pl.: búvóhelyek révén). Így a ragadozók csak N-NR zsákmányhoz juthatnak. Az elérhetetlen zsákmány lehet: N állandó hányada: NR=cN, ez ekvivalens azzal ha az a és q konstansokat a(1-c) és q(1-c)-re változtatnák, mely nem befolyásolja a modellt. NR az N-től független.")
66
A ragadozók funkcionális válasza
A ragadozók táplálkozási viselkedése is hatással van ragadozó-zsákmány-rendszerekre. A ragadozók táplálkozási viselkedésének és az elfogyasztott zsákmány mennyiségének a zsákmánypopuláció egyedsűrűségétől való függését funkcionális válasznak nevezzük. Grafikus megjelenítés: a ragadozó által egységnyi idő alatt elejtett zsákmány mennyisége. Holling (1959) három fő típust különített el:
 három fő típust különített el:")
67
1. Olyan állatok amelyek szűrögetés útján jutnak táplálékhoz, egyenes arányban növekedik egy telítési szintig. 2. A legtöbb ragadozó, a tápláléksűrűség növekedésével egyre csökkenő keresési idő és a kereséstől függetlenül a kezelésre fordítandó idő (TH) határozza meg: Ne= a’TsN Ahol, Ne a ragadozó által elfogyasztott zsákmány mennyisége; a’ támadási ráta; Ts keresési idő: Ts= T-THNe Átrendezve: Ne= a’NT/(1+a’THN) Holling –féle korong egyenlet
 határozza meg: Ne= a’TsN. Ahol, Ne a ragadozó által elfogyasztott zsákmány mennyisége; a’ támadási ráta; Ts keresési idő: Ts= T-THNe. Átrendezve: Ne= a’NT/(1+a’THN) Holling –féle korong egyenlet.")
69
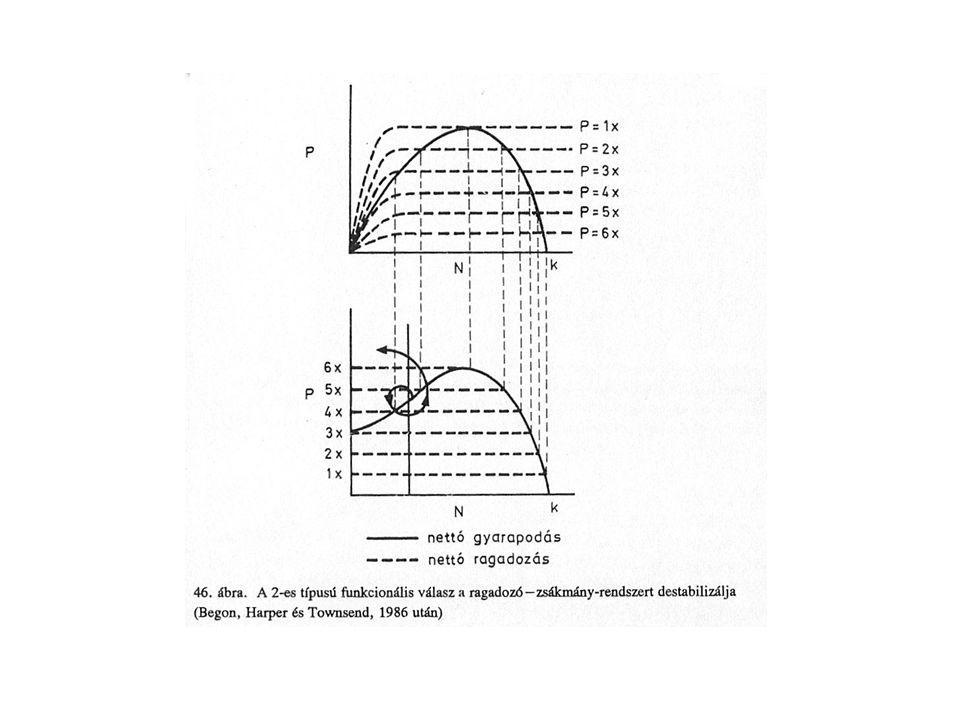
A 3. típusú funkcionális választ mutató ragadozók a zsákmánypopuláció egyedsűrűségének növekedésével egyre javuló hatásfokkal zsákmányolnak, míg alacsony zsákmánysűrűségnél az adott fajból relatíve keveset fogyasztanak. Valószínűleg keresési tevékenységének egy-vagy több komponense is egyedsűrűségfüggő. A prédaszám növekedésére bekövetkező prefenciaváltozást switching-nek nevezik. Zsákmány-búvóhelyen alacsony egyedszámban túlél, ragadozó alacsony egyedszám alternatív prédán-stabil egyensúly.

70
Természetes populációk azonban nem jellemezhetők egyetlen egyensúlyi helyzettel
több egyensúlyi pontot tartalmazó rendszer megjósolhatatlanul viselkedik kisebb változások hatására.

71
Irodalom Szentesi Árpád, Török János, Állatökológia. Kovásznai kiadó, Budapest. Begon, M., Harper, J.L., Townsend, C.R.,1996. Ecology. third edition, Blackwell Science Ltd. London, UK Kot, M Elements of Mathematical Ecology. Cambridge University Press, Cambridge, UK Kozár, F., Samu, F., Jermy, T., Az állatok populáció dinamikája. Akadémiai kiadó, Budapest.









>")






>")



”>")



