Előadást letölteni
1
A Biotechnológiai Tanszék oktatási anyaga
1 előadás A Biotechnológiai Tanszék oktatási anyaga az alábbi internet címen érhető el:

3
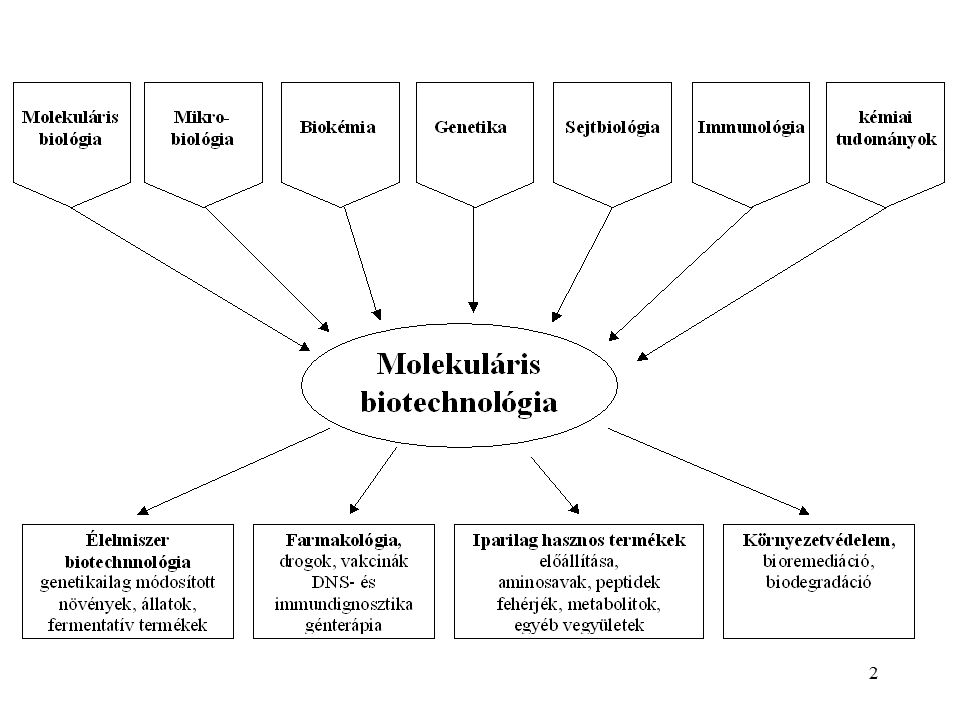
A FEHÉRJÉK TÚLTERMELTETÉNEK ELŐNYEI
Fő ok: Ha egy fehérjére nagy mennyiségben van szükség, és a természetben csak kis mennyiségben fordul elő, vagy az eredeti sejtvonal nehezen kezelhető, belõle fehérje csak körülményesen nyerhető. KÖZVETLEN FELHASZNÁLÁS IPARI, proteázok, szénhidrátbontó enzimek, lipázok, polimerázok,bioaktív peptidek, fehérjék, hormonok, farmakológiai, gyógyászati felhasználásra KUTATÁSI CÉLOKRA biokémiai és biofizikai vizsgálati módszerek, 3 dimenziós röntgenszerkezet, mutációs analízis, FELTÉTEL: A felhasználás előtt igazolni kell, hogy az eredeti fehérje és a rekombináns megfelelője biológiailag ekvivalens

5
DNA RNA 1 U H 3 OH 2

7
Egyéb tisztítási módszerek
DNS etanollal, izopropilalkohollal kicsapható Kromoszómális DNS nagy, makroszkopikus molekula kiemelhető Minél nagyobb egy DNS, annál könnyebben törik. Kb. 20 kb – osnál nagyobb DNS erős fizikai behatásnak (vortexes kevertetés) nem tehető ki! Egyéb tisztítási módszerek kromatográfia, - ioncserés DEAE (dietil-aminoetil) kötés alacsony, elúció magas sókoncentráció esetén A DNS savas ionos kölcsönhatás hordozó - szilikagél alapú elválasztás magas sókoncentráció mellett felkötődik a DNS, alacsony sókoncentrációnál eluálódik
 nem tehető ki! Egyéb tisztítási módszerek kromatográfia, - ioncserés DEAE (dietil-aminoetil) kötés alacsony, elúció magas sókoncentráció esetén A DNS savas ionos kölcsönhatás. hordozó. - szilikagél alapú elválasztás. magas sókoncentráció mellett felkötődik a DNS, alacsony sókoncentrációnál eluálódik.")
8
RNS tisztítása - CsCl sűrűséggradiens centrifugálás
adott koncetrációjú CsCl oldat centrifugálás során grádienst képez, a DNS a saját sűrűségének megfelelő helyre vándorol és ott megáll. különböző konformációjú DNS-k szétválaszthatóak, nagy tisztaság RNS tisztítása RNS nagyon könnyen degradálódik, RNázok stabilak a tisztítás során RNáz mentesíteni kell mindent DEPC: dietil pirokarbonát

9
RNS tisztítása sok eltérő protokol,
ma univerzálisan használható rendszer a guanidium isotiocianát-fenol-kloroform oldattal történő extrakció egy lépésben sejtfeltárás és szerves-vizes folyadék fázisú extrakció. A vizes fázis csak totál RNS-t tartalmaz ( és némi kromoszómális DNS szennyezést, DNázI kezeléssel eltávolítható guanidin és izotiociánsav sója

10
eukarióták esetén stabil poliA farok
mRNS tisztítás eukarióták esetén stabil poliA farok AAAAAAAAA mRNS hordozó tRNS rRNS TTTTTTTTTT- magas só AAAAAAAAA mRNS TTTTTTTTTT- hordozó alacsony só AAAAAAAAA mRNS

11
DNS ELVÁLASZTÁSA ELEKTROFORÉZIS
denaturáló nem-denaturáló lúgos, (DNS, agaróz) formaldehid, glioxál, DMSO (RNS, agaróz) urea (DNS, akrilamid) Nincs roncsoló ágens méret > 50 kb 100bp-50 kb < 1000 bp Mátrix agaróz agaróz % poliakrilamid technika pulzáló gélelektroforézis hagyományos gélelektroforézis (5-20%) hagyományos LÁTHATÓVÁ TÉTEL: etídium bromid, interkaláló festék Þ nem-denturáló körülmények, elsőősorban duplaszálú DNS-t fest, de egyszálú nukleinsavakat is. A DNS 254 nm-en elnyel Þ energia a festékre Þ 590 nm-en emisszió, a festék maga 302 és 366 nm-en nyel el Lehet gélbe rakni, utófesteni, illetve a mintához adni. Érzékenység kb 10 ng Egyéb festékek, fluoreszkáló anyagok: pl. fluoreszcein, minden körülményközött radioaktivitás: minden körülmény között, 35S, 33P, 32P beta sugárzók .
 formaldehid, glioxál, DMSO. (RNS, agaróz) urea (DNS, akrilamid) Nincs roncsoló ágens. méret. > 50 kb 100bp-50 kb < 1000 bp. Mátrix. agaróz. agaróz % poliakrilamid. technika. pulzáló gélelektroforézis. hagyományos gélelektroforézis. (5-20%) hagyományos. LÁTHATÓVÁ TÉTEL: etídium bromid, interkaláló festék Þ nem-denturáló körülmények, elsőősorban duplaszálú DNS-t fest, de egyszálú nukleinsavakat is. A DNS 254 nm-en elnyel Þ energia a festékre Þ 590 nm-en emisszió, a festék maga 302 és 366 nm-en nyel el. Lehet gélbe rakni, utófesteni, illetve a mintához adni. Érzékenység kb 10 ng. Egyéb festékek, fluoreszkáló anyagok: pl. fluoreszcein, minden körülményközött. radioaktivitás: minden körülmény között, 35S, 33P, 32P beta sugárzók. .")
12
DNS IZOLÁLÁSA GÉLBŐL - közös pont: először elektroforetikus úton elválasztjuk a DNS-t - a megfelelő sávot kivágjuk steril pengével AGARÓZ dialízis, dializáló hártyában, elektrodialízis a kapott DNS további tisztítása fenolos extrakcióval, alkoholos kicsapással, univerzális, széles mérettartomány - fagyasztásos módszer : a kivágott – DNS-t tartalmazó - agarózdarabot –80oC-on megfagyasztjuk az agarós szerkezete roncsolódik, ebből a DNS egy szűrőn keresztül centrifugálással kinyerhető további tisztítás szükséges, univerzális - kromatográfiás módszerek 6M NaI mellett a DNS 55oC-on (az agaróz megolvad) szilikagél felületére kötődik, a mátrix mosása után innen o55 C-on alacsony só(víz, TE) eluálható, majdnem univerzális, elég széles mérettartomány DEAE membránba futtatjuk a DNS-t, innen magassókoncentrációval (1.5M) magas hőmérsékleten eluálható nagy tisztaság, szűk, alacsony mérettartomány < 1.5 – 2 kb esetén jó a kitermelés POLIAKRILAMID (PAGE) a kivágott darabot passzív módon vagy elektromos térben eluáljuk majd töményítjük, ioncserésen tisztítjuk
 szilikagél felületére kötődik, a mátrix mosása után innen o55 C-on alacsony só(víz, TE) eluálható, majdnem univerzális, elég széles mérettartomány. DEAE membránba futtatjuk a DNS-t, innen magassókoncentrációval (1.5M) magas hőmérsékleten eluálható. nagy tisztaság, szűk, alacsony mérettartomány < 1.5 – 2 kb esetén. jó a kitermelés. POLIAKRILAMID (PAGE) a kivágott darabot passzív módon vagy elektromos térben eluáljuk. majd töményítjük, ioncserésen tisztítjuk.")
13
DNS/ RNS MÉRÉSE Ez leginkább oligonukleotidok esetén használatos.
A különböző nukleotidoknak 260 nm körül erős abszorbanciájuk van. Egyszálú DNS esetén: e(µmol * cm/cm3) bázis dA 15.4 dG 11.7 dC 7.5 dT 8.8 átlagosan e = 10 (µmol * cm)/cm3 Ez leginkább oligonukleotidok esetén használatos. Nagyobb egyszálú, kétszálú DNS-k, RNS-k esetén a bázisok kölcsönhatásamiatt a számítás módosul 1 cm-es küvettában 260 nm-en 1.0 az abszorbanciája 50 µg/ml kettősszálú DNS-t 33 µg/ml egyesszálú DNS-t 40 µg/ml egyesszálú RNS-t tartalmazó oldatnak A fehérjék 280 nm-en nyelnek el. Az A260nm/A280nm ará’ny a nukleinsav tisztaságára jellemző A260nm/A280nm 2 tiszta a preparátum A260nm/A280nm 1 - 1,5 sok a fehérje szennyezés Egyéb módszerek: fluoreszcein, kemilumineszcens festékekkel. Érzékenyebb, de fluorimétert igényel.
 bázis. dA dG dC dT átlagosan e = 10 (µmol * cm)/cm3. Ez leginkább oligonukleotidok esetén használatos. Nagyobb egyszálú, kétszálú DNS-k, RNS-k esetén a bázisok kölcsönhatásamiatt a számítás módosul. 1 cm-es küvettában 260 nm-en 1.0 az abszorbanciája. 50 µg/ml kettősszálú DNS-t. 33 µg/ml egyesszálú DNS-t. 40 µg/ml egyesszálú RNS-t. tartalmazó oldatnak. A fehérjék 280 nm-en nyelnek el. Az A260nm/A280nm ará’ny a nukleinsav tisztaságára jellemző. A260nm/A280nm 2 tiszta a preparátum. A260nm/A280nm 1 - 1,5 sok a fehérje szennyezés. Egyéb módszerek: fluoreszcein, kemilumineszcens festékekkel. Érzékenyebb, de fluorimétert igényel.")
14
A NUKLEINSAVAK MANIPULÁCIÓJA SORÁN HASZNÁLATOS ENZIMEK
RESTRIKCIÓS ENDONUKLEÁZOK II típus Speciális célszekvenciát ismer fel és hasítja a DNS-t mindkét szálon a felismerési szekvenciában vagy annak környezetében pl. EcoRI Mg2+ kell Ez a típus az elterjedt

15
A NUKLEINSAVAK MANIPULÁCIÓJA SORÁN
HASZNÁLATOS ENZIMEK RESTRIKCIÓS ENDONUKLEÁZOK Enzim Felismerőhely hossza Felismerő-hasítóhely Forrás organizmus AluI 4 AG/CT Arthrobacter luteus HphI 5 GGTGAN8/ Haemophilus parahaemolyticus EcoRI 6 G/AATTC Escherichia coli BamHI 6 G/GATCC Bacillus amyloliquefaciens Ragadós végeket adó hasítások (sticky end) 5' túlnyúló Sal I 6 5' G TCGAC 3' 3' CAGCT G 3' Streptomyces albis 3' túlnyúló Kpn I 6 5' GGTAC C 3' 3' C CATGG 5' Klebsiela pneumonia Tompa végű hasítások (blunt end) Hae III 4 5' GG CC 3' 3' CC GG 5' Haemophilus aegyptus
 5 túlnyúló. Sal. I G TCGAC 3 3 CAGCT G 3 Streptomyces albis. 3 túlnyúló. Kpn. I GGTAC C 3 3 C CATGG 5 Klebsiela pneumonia. Tompa végű hasítások (blunt end) Hae. III GG CC 3 3 CC GG 5 Haemophilus aegyptus.")
16
FEHÉRJE TÚLTERMELTETÉS SZEMPONTJÁBÓL
Isoschisomers AtcI ' GGTAC/C 3' KpnI ' GGTAC/C 3' XmaI ' C/CCGGG 3' SmaI ' CCC/GGG 3' Kompatibilis véget adó enzimek SalI ' G/TCGAC 3' XhoI ' C/TCGAG 3' FEHÉRJE TÚLTERMELTETÉS SZEMPONTJÁBÓL KITÜNTETT ENZIMEK AflIII ' A/CPuPyGT 3' BspHI ' T/CATGA 3' NcoI ' C/CATGG 3' NdeI ' CA/TATG 3'

17
DNS METILÁZOK - dam metiláz (dezoxiadenin metiláz)
5’ GATC 3’ felismerő hely N6 pozícióban metilez adenin sok dam metiláz érzékeny enzim van pl. MboI, XhoI, egyenként ellenőrizni kell, érzékenység esetén - dam- törzs használata - nem érzékeny izoskizomer használata (ha van) (MboI - Sau3AI)
 (MboI - Sau3AI)")
18
dcm metiláz (dezoxicitozin metiláz)
5’ CCAGG 3’ vagy 5’ CCTGG 3’, C5 pozícióban metilez - hasonlóképpen léteznek dcm metiláz érzékeny enzimek - megoldás ugyanaz, mint a dam esetén citozin Sok metiláz van még, hasonló felismerő kanonikus szekvenciával, mint a restrikciós enzimek pl. M. EcoRI metiláz

19
Polimerázok DNS függő DNS polimerázok RNS függő DNS polimerázok
Templátfüggetlen DNS polimeráz DNS függő RNS polimeráz 5’ 3’ polimeráz aktivitás DNS polimerázoknak primer (indítószekvencia) kell RNS polimerázoknak promóter
 kell. RNS polimerázoknak promóter.")
20
dNTP: dATP, dCTP, dGTP, dTTP
DNS függő DNS polimerázok E. coli DNS polimeráz I aktivitásai: 5’ 3’ polimeráz 5’ 3’ 5’…pCpApGpTOH 3’ 3’…pGpTpCpApApCpGpGpTpTp… 5’…pCpApGpTpTPGpCpCpApAp… 3’ E. coli DNS polimeráz I dNTP: dATP, dCTP, dGTP, dTTP

21
E. coli DNS polimeráz I aktivitásai:
5’ 3’ exonukleáz E. coli DNS polimeráz I 5’ 3’ 5’ 3’ 5’CpApGpTpTPGpCpCpApAp… 3’ 3’GpTpCpApApCpGpGpTpTp… 5’ 5’ CpCpApAp… 3’ 3’…pGpTpCpApApCpGpGpTpTp… 5’ E. coli DNS polimeráz I 3’ 5’ exonukleáz E. coli DNS polimeráz I 3’ 5’ 3’ 5’ 3’ 5’ 5’…CpApGpTpTPGpCpCpApAOH 3’ 3’…GpTpCpApApCpGpGpTpTp 5’ 5’…CpApGpTpT 3’ 3’…GpTpCpApApCpGpGpTpTp… E. coli DNS polimeráz I

22
E. coli DNS polimeráz I Klenow fragment és a T4 DNS polimeráz aktivitásai:
5’ 3’ 5’…pCpApGpTOH 3’ 3’…pGpTpCpApApCpGpGpTpTp… 5’…pCpApGpTpTPGpCpCpApAp… 3’ Klenow, vagy T4 polimeráz dNTP: dATP, dCTP, dGTP, dTTP 3’ 5’ exonukleáz Klenow, vagy T4 polimeráz 3’ 5’ 3’ 5’ 3’ 5’ 5’…CpApGpTpTPGpCpCpApAOH 3’ 3’…GpTpCpApApCpGpGpTpTp 5’ 5’…CpApGpTpT 3’ 3’…GpTpCpApApCpGpGpTpTp… NINCS 5’ 3’ exonukleáz

23
DNS függő termofil DNS polimerázok

24
Klenow (fragment) polimeráz
DNS függő DNS polimerázok 5’ 3’ polimeráz 3’ 5’ polimeráz 5’ 3’ exonukleáz DNS polimeráz I Van Klenow (fragment) polimeráz Nincs T4 DNS polimeráz Van, erős Taq polimeráz Van/nincs
 polimeráz. Nincs. T4 DNS polimeráz. Van, erős. Taq polimeráz. Van/nincs.")
25
Templát független DNS polimeráz

26
RNS függő DNS polimeráz

27
DNS jelölés, nick transzláció

28
DNS jelölés, random priming
N: A,C,G,T (NNNNNN) denaturálás hibridizáció DNS polimeráz dNTP polimerizáció denaturálás Jelölt szálak
 denaturálás. hibridizáció. DNS polimeráz dNTP. polimerizáció. denaturálás. Jelölt szálak.")
30
DNS függő RNS polimerázok

38
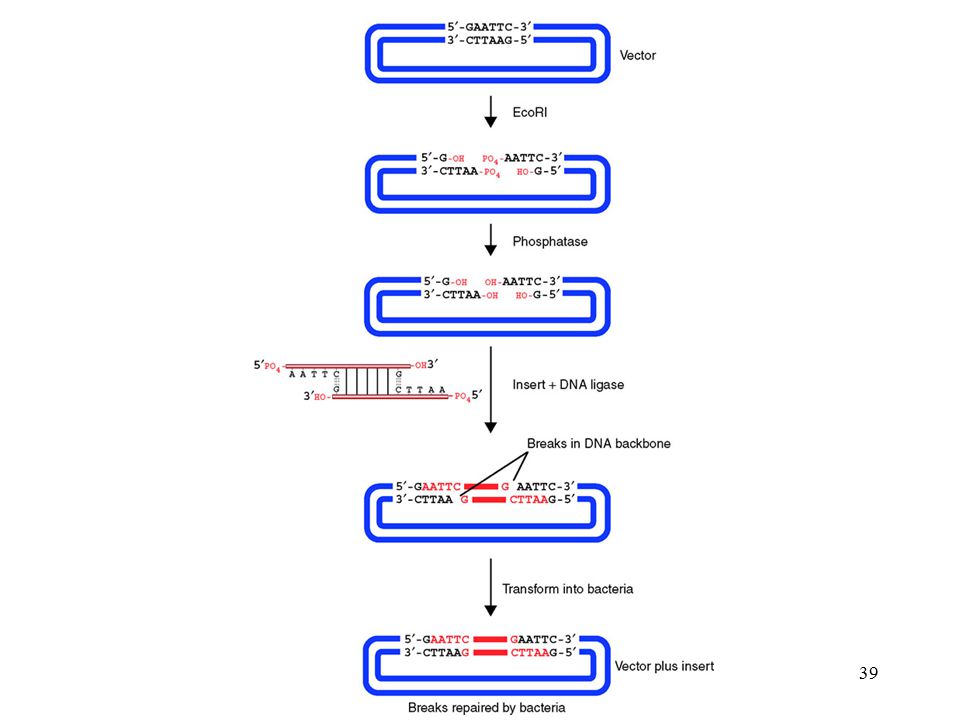
Klónozás fogalma ligálás vektor
Egy általunk kiválasztott DNS darabot vektor segítségével gazdasejtbe juttatunk és ott felszaporítunk Szubklónozás: további kisebb darabok hasonló felszaporítása vektor Hasítás, A,B enzimekkel Hasítás, A,B enzimekkel A inszert A B ligálás B Transzformálás, felszaporítás, tisztítás Vektor: olyan nukleinsav hordozó, amellyel nukleinsavakat sejtbe lehet juttatni, Felhasználás: klónozás, fehérje termeltetés, genetikai manipulációk stb.

40
KÓNOZÁSBAN ÁLTALÁNOSAN HASZNÁLT
VEKTORTÍPUSOK inszert méret példa plazmidok pUC18,19 < kb fonalas fágok mp18, 19 < kb fagemidek pBluescriptKS, SK± < kb l fágok EMBL3,4 néhányszor 10 kb kozmidok pHC79 néhányszor 10 kb néhány 100 kb BAC, YAC pBAC108L, pYAC3 BAC, YAC: bacterial, yeast artificial chromosome PLAZMIDOK replikációs origo ORI rezisztencia marker Cirkuláris kettősszálú extrakromoszómális elemek

42
Egy ősi plazmid: pBR322

43
Magas kópiaszámú változat: pUC19

44
Inszertet tartalmazó klónok kiválasztása
antibiotikum rezisztencia, ld. pBR3222, két antibiotikum, az egyik elromlik, ha inszert épül be, fáradtságos szurkálások, két antibiotikum rezisztencia gén szükséges kolónia hibridizáció, univerzális mindig használható plazmid tisztítás, térképezés restrikciós emésztéssel hosszú fáradtságos polimeráz láncreakció sejteken, kombinatorikus gyors ha nincs más szelekció kék fehér színszelekció, pozitív szelekciós vektorok, kondicionálisan letális gén a vektoron, az inszert beépül, elrontja a gént megszűnik a letalitás auxotrofiát komplementáló génbe történő klónozás ugyanaz a probléma, mint az antibiotikumok esetén

45
Kolónia hibridizáció

46
LacZ a komplementáció F' plazmidon: defektív b- galaktozidáz gén, hiányzik aminosav közötti régió bevitt vektor: tartalmazza a lacZ szabályozó régiót és az aminosavat a kettő együtt: aktív b – galaktozidáz X-gal szubsztráttal kék telep a bevitt N-terminális fragmentben : polilinker régió (leolvasási keret marad) ebbe lehet klónozni fragmentumot, ha kis fragmentum és leolvasási keret nem romlik el X-gal szubsztráttal kék telep, ha elromlik vagy nagy fragment X-gal szubsztráttal fehér telep
 ebbe lehet klónozni fragmentumot, ha kis fragmentum és leolvasási keret nem romlik el X-gal szubsztráttal kék telep, ha elromlik vagy nagy fragment X-gal szubsztráttal fehér telep.")
47
PLAZMIDOK SEJTBE JUTTATÁSÁNAK MÓDJAI
1. Kémiai transzformálás Kompetens sejt: a DNS felvételére alkalmassá tett sejt A sejteket felnövesztés után centrifugáljuk speciális kétértékű kationokat (Ca2+, Mn2+) tartalmazó oldattal kezeljük, sejtfal permeabilitást növelő ágenst (DMSO) adunk hozzá transzformációs hatékonyság: transzformáns /µg DNS elvi szám a transzformációt£ 1 ng mennyiségű DNS-sel hajtjuk végre : normál érték: 106 – 108 nagyon jó: 109 a transzformáció hatékonyságát meghatározó tényezők: oldatok edények tisztasága, - sejtek növekedési sebessége, a növesztés fázisa, hőmérséklete hősokk hőmérséklete hossza permeabilizáló faktor a lineáris DNS transzformációs gyakorisága kb 2 nagyságrenddel alacsonyabb, mint a cirkulárisé egyéb fogások: spheroplast készítés ozmotikum jelenlétében és ezt transzformáljuk - a DNS-t liposzómába csomagoljuk transzfomálás előtt
 tartalmazó oldattal kezeljük, sejtfal permeabilitást növelő ágenst (DMSO) adunk hozzá. transzformációs hatékonyság: transzformáns /µg DNS. elvi szám a transzformációt£ 1 ng mennyiségű DNS-sel hajtjuk végre. : normál érték: 106 – 108. nagyon jó: 109. a transzformáció hatékonyságát meghatározó tényezők: oldatok edények tisztasága, - sejtek növekedési sebessége, a növesztés fázisa, hőmérséklete. hősokk hőmérséklete hossza. permeabilizáló faktor. a lineáris DNS transzformációs gyakorisága kb 2 nagyságrenddel alacsonyabb, mint a cirkulárisé. egyéb fogások: spheroplast készítés ozmotikum jelenlétében és ezt transzformáljuk. - a DNS-t liposzómába csomagoljuk transzfomálás előtt.")
48
Transzformálás hatékonyságát meghatározó tényezők II.
hősokk hőmérséklete, hossza plazmid mérete tárolhatóság permealizáló ágens

49
Elektroporáció A sejteket felnövesztés után kis vezetőképességű, glicerines (nagy ellenállású 600 ) pufferben szuszpendáljuk nagy feszültségű impulzust adunk rá kb 5 ms-ig transzformációs hatékonység x jobb (1010/µg DNS) sejttípusonként optimalizálni kell maghatározó faktorok: - az oldat ellenállása - az impulzus nagysága, hossza - permeabilizáló, redox potenciált befolyásoló faktorok adagolása
 sejttípusonként optimalizálni kell. maghatározó faktorok: - az oldat ellenállása. - az impulzus nagysága, hossza. - permeabilizáló, redox potenciált befolyásoló faktorok adagolása.")
50
KONJUGÁCIÓ Sejtből sejtbe történő DNS átadás lépései:
párosodás: speciális kontaktus a donor és a recipiens között egy speciális sejtfelszíni ponton keresztül (pl. pilus) DNS átjuttatását közvetítő folyamatok, replikcáció (rolling circle, az egyik szál átjutása) konjugációs elemek donorból donort csinál az első ilyen a a szex faktor, F episzóma másik: IncP csoportba tartozó: RP4, RK2 plazmidok (szilárd fázishoz mobilizálható plazmidok a recipiensből nem lesz donor a plazmid tartalmazza a DNS processzáló apparátust, oriT, mob régió tra gének, pilus: N-acetilált TraA
 DNS átjuttatását közvetítő folyamatok, replikcáció (rolling circle, az egyik szál átjutása) konjugációs elemek. donorból donort csinál. az első ilyen a a szex faktor, F episzóma. másik: IncP csoportba tartozó: RP4, RK2 plazmidok (szilárd fázishoz. mobilizálható plazmidok. a recipiensből nem lesz donor. a plazmid tartalmazza a DNS processzáló apparátust, oriT, mob régió. tra gének, pilus: N-acetilált TraA.")
51
A DNS transzfer mechanizmusa a konjugáció során
donor recipiens donor recipiens TraI 5’ 5’ TraI Töréspont, nick donor recipiens Egyszálú DNS kötő fehérje TraC primáz 5’ RNS primer Replikatív DNS polimeráz TraI

52
Mobilizálható vektorok
Nem tartalmazzák a tra géneket, csak a transzferhez szükséges oriT-t tra gének: integrálva a kromoszómába, három komponensű konjugáció: sem a donor sejt sem a recipiens nem tartalmazza a transzferhez szükséges géneket, hanem egy harmadik sejt

53
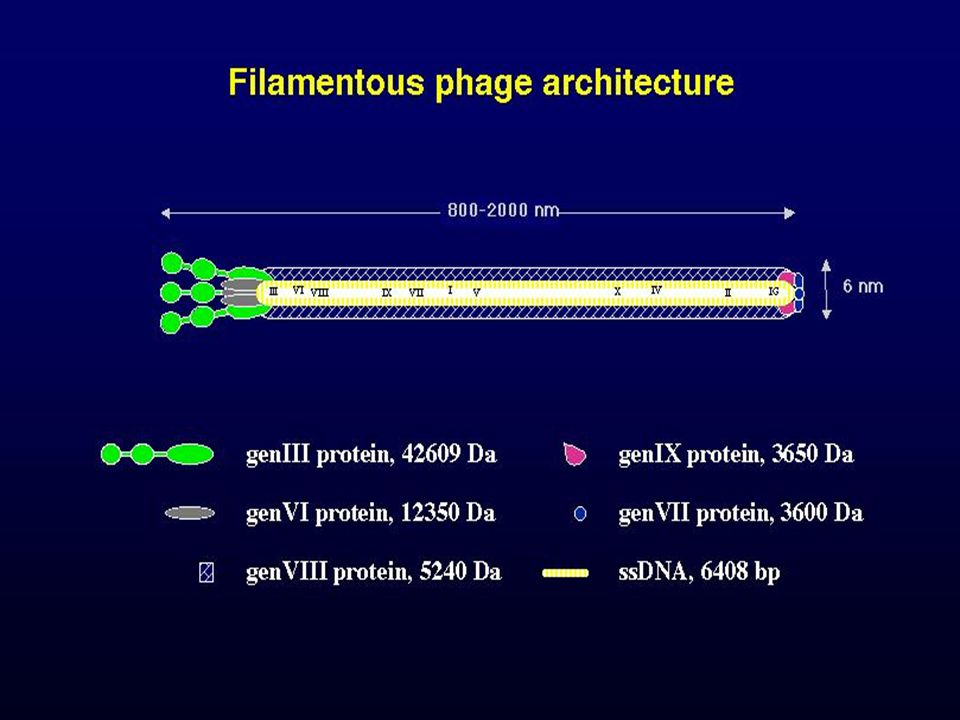
Fonalas fágok I. M13, f1 és fd fágok, genomjuk 98%-ban azonos rekombinálnak egymással Az érett fágok genomja egyszálú cirkuláris DNS, a sejten belül: RF, replikatív forma ez alkalmas genetikai manipulációkra A fág genomjának 90%-a 10 fehérjét kódol, nagyobb intergenikus régió a VIII és a III gének illetve II és IV gének között, regulátor funkció A fág a szexpiluson keresztül fertőz, a DNS (+) szál jut be a sejtbe ebből lesz kétszálú replikatív forma perc, kópia Nem öli meg a gazdasejtet, csak lassítja a növekedést VIII fehérje a fő strukúr protein, 2700 kópia, a III. számú fehérje a filament végén néhány kópiában Az RF forma képződése után transzkripció, transzláció, Replikáció: II fehérje töréspontot idéz elő, DNS polimeráz I polimerizál WC papír modell. a II. fehérje vagdossa egységnyi darabokra V. fehérje: egyszálú DNS kötő fehérje, represszálja a II gén expresszióját kópiaszám kontroll
 szál jut be a sejtbe ebből lesz kétszálú replikatív forma perc, kópia. Nem öli meg a gazdasejtet, csak lassítja a növekedést. VIII fehérje a fő strukúr protein, 2700 kópia, a III. számú fehérje a filament végén néhány kópiában. Az RF forma képződése után transzkripció, transzláció, Replikáció: II fehérje töréspontot idéz elő, DNS polimeráz I polimerizál WC papír modell. a II. fehérje vagdossa egységnyi darabokra. V. fehérje: egyszálú DNS kötő fehérje, represszálja a II gén expresszióját. kópiaszám kontroll.")
55
A fonalas fágok életciklusa

56
Egy fonalas fág térképe

58
Epitóp könyvtár készítése, fág display vektorok
gVIII (vagy gIII) gén (NNN)x X : 6 vagy hosszabb
 gén. (NNN)x. X : 6 vagy hosszabb.")
59
Egy példa: anthrax toxin inhibitor tervezése phage display technikával
Az anthrax toxin érése, patogenezise, potenciális célpontok A (PA63) heptamert felkötjük egy oszlopra
 heptamert felkötjük egy oszlopra.")
60
Egy példa: anthrax toxin inhibitor tervezése phage display technikával II.
TYWWLD ACCTATTGGTGGCTGGAT DNS izolálás, szekvenálás (NNN)x A (PA63)7 oszlopon kromatografáljuk elúció specifikus felkötődik átfolyó
x. A (PA63)7 oszlopon kromatografáljuk. elúció. specifikus felkötődik. átfolyó.")
61
Fagemidek: fonalas fágok és plazmidok hibridjei

62
a középső, centrális régió eltávolítható
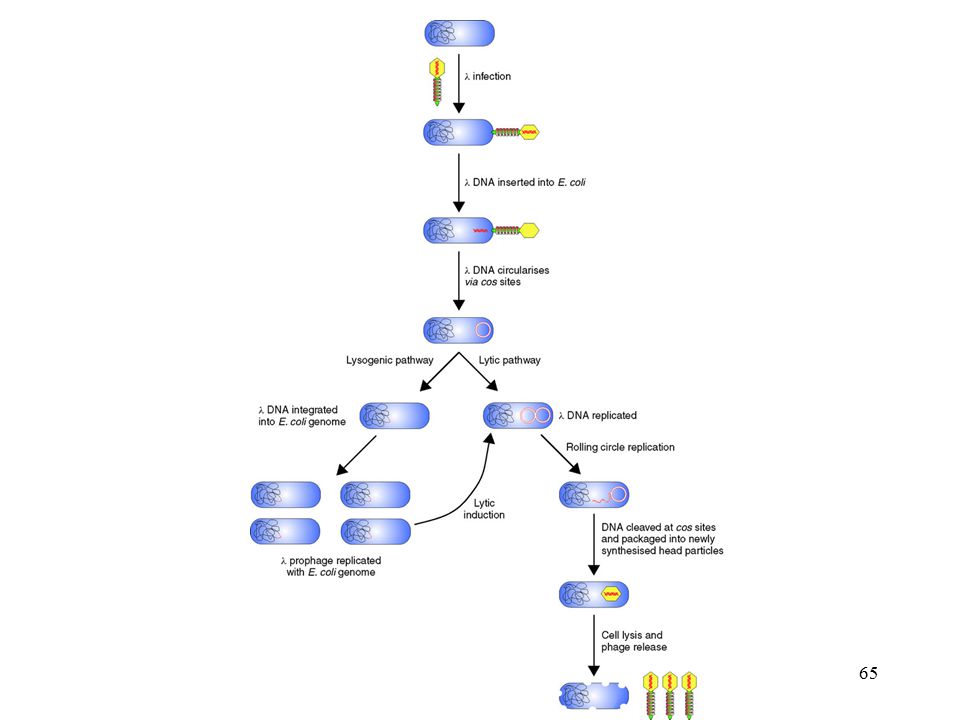
A fágok általános felépítése genetikai anyag: kb duplaszálú lineáris DNS fertőzéssel jut a sejtbe, nagy hatékonyság két életciklus: - lizogén: a fág genetikai anyaga beépül a kromoszómába, a sejt túlél - lítikus: érett fág képződése során a sejtek lizálnak elpusztulnak a végeken ragadós. ún. ’cos’ végek bal kar 20 kb centrális régió 14 kb jobb kar 9 kb cos a középső, centrális régió eltávolítható EcoRI BamHI SalI

64
A lambda fág életciklusa

66
Lambda fág példák Helyettesítő/ replacement vektorok
Inszerciós vektorok

67
A klónozás menete helyettesítő vektorok esetén
A pakolódás feltétele, hogy a rekombináns gemon mérete a vad típus méretének 79 – 105 %- a lehet.

68
Génátvitel transzdukcióval baktériumokban
bakteriális DNS-t is tartalmazó többnyire defektív fágok fág DNS intakt normál fág transzdukáló fág bakteriális fágfertőzés lizogén életciklus fág genom integrálódása a baktérium kromoszómájába a fág által hordozott bakteriális DNS beépülése rekombinációval

69
plazmidok és l fágok hibridjei
Kozmidok: plazmidok és l fágok hibridjei fertőzéssel bejuttatható plazmidok

70
GÉNEK ELŐÁLLÍTÁSÁNAK STRATÉGIÁI
Gének izolálása A megfelelő organizmus génkészletéből a kívánt régiót tartalmazó szakasz valamilyen módon való kinyerése elterjedtebb, általában gyorsabb, kevesebb munkát igényel (ha lehetséges) nem feltétlenül szükséges szekvencia információ, de nem árt, ha van a különböző organizmusok genom szekvenciái nagy segítséget jelenthetnek hátrány: kodon preferencia adott, nem tervezhető Gének szintézise Szintetikus oligonukleotidokból történő összeépítés kevésbé elterjedt a gén hosszától függően sok munkát igényelhet, a fehérje elsődleges szekvenciáját ismerni kell előny: tervezhető a biológia szabályainak figyelembevételével, az optimális kodon felhasználás a gazda sejthez igazítható, az optimális fehérje túltermeltetési tapasztalatok jobban kamatoztathatók
 nem feltétlenül szükséges szekvencia információ, de nem árt, ha van. a különböző organizmusok genom szekvenciái nagy segítséget jelenthetnek. hátrány: kodon preferencia adott, nem tervezhető. Gének szintézise. Szintetikus oligonukleotidokból történő összeépítés. kevésbé elterjedt. a gén hosszától függően sok munkát igényelhet, a fehérje elsődleges szekvenciáját ismerni kell. előny: tervezhető a biológia szabályainak figyelembevételével, az optimális kodon felhasználás a gazda sejthez igazítható, az optimális fehérje. túltermeltetési tapasztalatok jobban kamatoztathatók.")
71
KÖNYVTÁRAK TÍPUSAI - genomiális
a teljes genomból készül, elvileg minden információt tartalmaz (pl. nem transzlálódó régió, szabályozó régiók, intronok) mind prokariótákból, mind eukariótákból - cDNS az aktívan termelődő mRNS-ekből készül csak az érett mRNS szekvenciáját tartalmazza (nincs intron szabályozó régió stb) csak eukariótákból expressziós a cDNS-eket ún. expressziós kazettába klónozzuk, ezáltal a kódolt fehérje (ha a mRNS kódol ilyet) aktívan termelődhet Követelmények a könyvtárakkal szemben - fedje le a teljes genomot, illetve mRNS populációt - ne legyen redundáns
 mind prokariótákból, mind eukariótákból. - cDNS. az aktívan termelődő mRNS-ekből készül. csak az érett mRNS szekvenciáját tartalmazza. (nincs intron szabályozó régió stb) csak eukariótákból. expressziós. a cDNS-eket ún. expressziós kazettába klónozzuk, ezáltal a kódolt fehérje (ha a mRNS kódol ilyet) aktívan termelődhet. Követelmények a könyvtárakkal szemben. - fedje le a teljes genomot, illetve mRNS populációt. - ne legyen redundáns.")
72
in vitro pakolás: összekeverés l fágfehérje extraktummal
GENOMIÁLIS KÖNYVTÁR KÉSZÍTÉSÉNEK SÉMÁJA hasító helyek bal kar jobb kar centrális régió emésztés részleges vagy kb méretű teljes emésztés fragmentek bal kar jobb kar ligálás konkatamer in vitro pakolás: összekeverés l fágfehérje extraktummal A pakolódás feltétele, hogy a rekombináns gemon mérete a vad típus méretének 79 – 105 %- a lehet.

73
Bakteriális shot-gun könyvtár készítése
E. coli transzformálás elektroporálás kromoszómális DNS nebulizátor 2-3,5 kb fragmentek végek tompítása defoszforilálás összetört DNS Preparatív gél elektroforézis

74
A mRNS-ekkel szemben támasztott követelmények
cDNS könyvtár A mRNS-ekkel szemben támasztott követelmények Integritás A mRNS mérete, degradált-e Megoldás: denaturáló gélelektroforézis (várt méret kb, a többség kb) Lehet-e teljes hosszúságú cDNS-t szintetizáltatni Megoldás: elõzetes reverz transzkripció jelölt nukloetidokkal elektroforézis Lehet-e nagy molekulasúlyú fehérjéket in vitro szintetizálni Megoldás: in vitro transzláció jelölt aminosavakkal A kérdéseses fehérjét tudjuk-e szintetizáltatni Megoldás: in vitro transzláció + immunoprecipitáció
 Lehet-e teljes hosszúságú cDNS-t szintetizáltatni. Megoldás: elõzetes reverz transzkripció jelölt nukloetidokkal. elektroforézis. Lehet-e nagy molekulasúlyú fehérjéket in vitro szintetizálni. Megoldás: in vitro transzláció jelölt aminosavakkal. A kérdéseses fehérjét tudjuk-e szintetizáltatni. Megoldás: in vitro transzláció + immunoprecipitáció.")
75
A cDNS könyvtár mérete A kérdéses mRNS előfordulási gyakorisága
különböző mRNS emlős sejtekben nagy gyakoriságú %, pl. globin nem problematikus kis gyakoriságú 0.5 % nagy könyvtárat kell készíteni, és a detektálás is gond N = ln(1-P) / ln(1-1/n) N: a szükséges klónok száma a könyvtárban, P: annak a valószínûsége, hogy a keresett klón benne legyen a könyvtárban, 1/n: az a mRNS frakció, ami 1 db keresett mRNS-t tartalmaz immunoprecipitált in vitro transzlált termék/ összes transzlált termék pl. 14 molekula/sejt, 1/n = 37000, P= 0.99, N=
 / ln(1-1/n) N: a szükséges klónok száma a könyvtárban, P: annak a valószínûsége, hogy a keresett klón benne legyen a könyvtárban, 1/n: az a mRNS frakció, ami 1 db keresett mRNS-t tartalmaz. immunoprecipitált in vitro transzlált termék/ összes transzlált termék. pl. 14 molekula/sejt, 1/n = 37000, P= 0.99, N=")
76
cDNS-könyvtár készítése primer adapter módszerrel

77
cDNS-könyvtár készítése

78
Szubsztraktív hibridizáció
nem indukált minta, mRNS tisztítás indukált minta, mRNS tisztítás AAAAAAAAA 3' 5' TTTTTTTTT biotin reverz transzkripció B RnázH immobilizált streptavidin SA cDNS könyvtár csak indukált mRNS átfolyó cDNS, jelölés hibridizáció

79
RENDELKEZÉSRE ÁLLÓ VAGY SZÜKSÉGES INFORMÁCIÓK A KERESETT FEHÉRJÉRŐL, ILLETVE GÉNJÉRŐL
- nem tudunk semmit - ismeretek a fehérje funkciójáról csak fenotípus alapján jellemezhető mutagenezis (transzpozon) kromoszomális séta van tisztított fehérje fehérje funkció tesztelhető expressziós könyvtárak fehérje szekvenálás DNS próba tervezés ellenanyag termeltetés már van ilyen fehérje illetve gén más típusú sejtekből heterológ próba használata könyvtárak átvizsgálásához EGYÉB pl. ha a fehérje szabályozása ismert szubsztraktív hibridizáció - számítógép, adatbankok genom szekvenálások
 kromoszomális séta. van tisztított fehérje. fehérje funkció tesztelhető. expressziós könyvtárak. fehérje szekvenálás. DNS próba tervezés. ellenanyag termeltetés. már van ilyen fehérje illetve gén más típusú sejtekből. heterológ próba használata könyvtárak átvizsgálásához. EGYÉB. pl. ha a fehérje szabályozása ismert szubsztraktív hibridizáció. - számítógép, adatbankok. genom szekvenálások.")
80
POZITÍV KLÓNOK KÜLÖNBÖZŐ KÖNYVTÁRAKBÓL VALÓ KIVÁLASZTÁSÁRA SZOLGÁLÓ MÓDSZEREK
kolónia vagy plakkhibridizáció, jelölt DNS-sel DNS próba előállítása: - szintetikus (degenerált) oligonukleotidok - megfelelően választott primerekkel elôállított PCR fragment - izolált DNS fragment (lehet heterológ is) Jelölési módszerek - radioaktív vagy nemradioaktív - 5' foszforilálás, vagy 3' végen feltöltéses végjelölés vagy terminális transzferázzal, "nick-transzláció", "random priming", PCR honnan lehet próbánk: a gén már ismert, legalábbis egy darabon heterológ próba: más mikroorganizmusokból már ismert hasonló géne van tiszított fehérjénk fehérje szekvenálás DNS próba tervezés reverz transzlációval
 oligonukleotidok. - megfelelően választott primerekkel elôállított PCR fragment. - izolált DNS fragment (lehet heterológ is) Jelölési módszerek. - radioaktív vagy nemradioaktív. - 5 foszforilálás, vagy 3 végen feltöltéses végjelölés vagy terminális transzferázzal, nick-transzláció , random priming , PCR. honnan lehet próbánk: a gén már ismert, legalábbis egy darabon. heterológ próba: más mikroorganizmusokból már ismert hasonló géne. van tiszított fehérjénk fehérje szekvenálás DNS próba tervezés reverz transzlációval.")
82
Kodon felhasználási preferencia – táblázat
Egy példa AmAcid Codon Number / Fraction Gly GGG Gly GGA Gly GGT Gly GGC Glu GAG Glu GAA Asp GAT Asp GAC Val GTG Val GTA Val GTT Val GTC Ala GCG Ala GCA Ala GCT Ala GCC Arg AGG Arg AGA Ser AGT Ser AGC Lys AAG Lys AAA Asn AAT Asn AAC Met ATG Ile ATA Ile ATT Ile ATC Thr ACG Thr ACA Thr ACT Thr A CC Trp TGG End TGA Cys TGT Cys TGC End TAG 0.10 End TAA Tyr TAT Tyr TAC Leu TTG Leu TTA Phe TTT Phe TTC Ser TCG [gbbct]: 50 CDS's (13879 codons)
")
83
cDNS-ből készült expressziós könyvtárból
- Immunológiai detektálás poli- vagy monoklonális ellenanyaggal - Aktivitás mérés (csak bizonyos szerencsés esetekben) - Valamilyen módon jelölt liganddal (ha van a keresett fehérjének)
 - Valamilyen módon jelölt liganddal (ha van a keresett fehérjének)")
84
ÉLESZTŐ KÉT HIBRID RENDSZER
fehérje-fehérje kölcsönhatáson alapuló szelekció GAL4 transzkripciós aktivátor aktivátor régió (AR) DNS kötő domén (DKD) lacZ GAL4 kötő hely DKD Protein X Protein Y AR lacZ GAL4 kötő hely DKD Protein X AR Protein Y kölcsönhatás esetén: kék telepek
 DNS kötő domén (DKD) lacZ. GAL4. kötő hely. DKD. Protein X. Protein Y. AR. lacZ. GAL4. kötő hely. DKD. Protein X. AR. Protein Y. kölcsönhatás esetén: kék telepek.")
85
BAKTERIÁLIS KÉT HIBRID RENDSZER
lacZ GAL4 kötő hely DKD Protein X Protein Y RNS pol. lacZ DKD Protein X RNS pol Protein Y GAL4 kötő hely kölcsönhatás esetén: kék telepek pozitív szelekció, nagyobb könyvtárakra kölcsönhatás esetén: növekedés hisztidin mentes táptalajon GAL4 kötő hely his3 DKD Protein X RNS pol Protein Y

86
A POZITÍV KLÓNOK TOVÁBBI FELDOLGOZÁSA
A könyvtárakból kapott klónok általában túl nagyok közvetlen felhasználáshoz, ezért további munkálatok szükségesek: 1. AZ INSZERT RESTRIKCIÓS TÉRKÉPEZÉSE 2. A TÉRKÉP ALAPJÁN AZ INSZERT KISEBB DARABOKBAN TÖRTÉNŐ SZUBKLÓNOZÁSA 3. AZ SZUBKLÓNOK SZEKVENCIÁJÁNAK MEGÁLLAPÍTÁSA 4. AZ INSZERT SZEKVENCIÁJÁNAK MEGÁLLAPÍTÁSA 5. SZÁMÍTÓGÉPES ADATFELDOLGOZÁS A SZEKVENCIÁN BELÜLI GÉNEK, ELEMEK AZONOSÍTÁSA

87
RESTRIKCIÓS TÉRKÉPEZÉS

88
Klónozás, szubklónozás
A A B C vektor Hasítás, A,B enzimekkel Hasítás, A,B enzimekkel A inszert A B ligálás B Transzformálás, felszaporítás, tisztítás

89
A POZITÍV KLÓNOK TOVÁBBI FELDOLGOZÁSA
A könyvtárakból kapott klónok általában túl nagyok közvetlen felhasználáshoz, ezért további munkálatok szükségesek: 1. AZ INSZERT RESTRIKCIÓS TÉRKÉPEZÉSE 2. A TÉRKÉP ALAPJÁN AZ INSZERT KISEBB DARABOKBAN TÖRTÉNŐ SZUBKLÓNOZÁSA 3. AZ SZUBKLÓNOK SZEKVENCIÁJÁNAK MEGÁLLAPÍTÁSA 4. AZ INSZERT SZEKVENCIÁJÁNAK MEGÁLLAPÍTÁSA 5. SZÁMÍTÓGÉPES ADATFELDOLGOZÁS A SZEKVENCIÁN BELÜLI GÉNEK, ELEMEK AZONOSÍTÁSA

90
AZ SZUBKLÓNOK SZEKVENCIÁJÁNAK MEGÁLLAPÍTÁSA

91
A POZITÍV KLÓNOK TOVÁBBI FELDOLGOZÁSA
A könyvtárakból kapott klónok általában túl nagyok közvetlen felhasználáshoz, ezért további munkálatok szükségesek: 1. AZ INSZERT RESTRIKCIÓS TÉRKÉPEZÉSE 2. A TÉRKÉP ALAPJÁN AZ INSZERT KISEBB DARABOKBAN TÖRTÉNŐ SZUBKLÓNOZÁSA 3. AZ SZUBKLÓNOK SZEKVENCIÁJÁNAK MEGÁLLAPÍTÁSA 4. AZ INSZERT SZEKVENCIÁJÁNAK MEGÁLLAPÍTÁSA 5. SZÁMÍTÓGÉPES ADATFELDOLGOZÁS A SZEKVENCIÁN BELÜLI GÉNEK, ELEMEK AZONOSÍTÁSA

92
De ha sikerül, és van szekvenciánk
Mi van rajta,van-e gén? Honnan tudjuk, hogy Valamit találtunk, találtunk-e gént? CTCGAGACGCTGTTTCTGGGGTCATTCATTCTTGGCGGGCTGCAACTGCTGGTGTGACCGACGCGACCTGGCAGGCCGCGGTGCGCAACTGGCCGGGCGGACTAATGGTGGAGCAAAAGA TCGGCATGTCCAGCGCACCTGAAGCTTGGGTGGTTGCTGCAATAGCAGCCTTCCTTATTGGCATGGCGAAGGGCGGTTTGGCCAATGTGGGGGTTATCGCCGTTCCCTTGATGTCCCTGG TCAAGCCGCCGCTTACCGCTGCCGGATTGCTGCTCCCGATCTATGTCGTTTCTGATGCATTCGGCGTCTGGCTTTATCGGCACCGGTATTCTGCCTCCAATCTGCGCATCCTGATTCCTT CGGGATTTTTTGGGGTCCTGATTGGCTGGTTATTGGCCGGGCAGATCTCCGACGCGATTGCCAGTGTCATTGTTGGTTTCACCGGCTGCGGCTTCGTGGCTGTGCTGCTGGCACGACGAG GGGTGCCATCGGTGCCGCGTCAAGCCAACGTGCCCAAAGGATGGTTTCTGGGGGTGGCCACCGGCTTTACCAGCTTTTTGACTCATTCCGGTGCGGCGACCTTCCAGATGTTCGTGCTGC CGCAACGGCTGGACAAGACCATGTTCGCGGGCACATCAACGCTTACCTTTGCTGCCATAAACCTATTCAAGATTCCGTCCTACTGGGCATTGGGACAGCTTTCGACTTCCTCGGTCATGT CCGCGCTAGTGTTGATTCCGGTGGCCGTGGCCGGGACGTTCGCAGGTGTTTTTGCGACGCGCAGGCTATCGACATCCTGGTTCTTCATTCTGGTCCAGGCGATGTTGCTGGTGGTCTCCA TTCAGCTTCTGTGGAGGGGAATGTCGGATATCCTGAACTAGCTGGAGATCGCAATGTCAGAACGCTCAATCAATCAGAATGTAATCTTGACATAGAATACCGTTCCGATTTATTGCTTCG AGTGAAGCTGCCCGTCCGCTGAGATGTCATGACATTTTCCCCGCTTGATTCCGCCCTGCTTGGACCGTTGTTCGCGACCGATGAAATGCGCACGGTCTTCTCCGAACGGCGTTTTTTGGC GGGAATGCTTCGTGTTGAAGTGGCCCTGGCGCGCGCGCAGGCGGCAGAGGGCCTTGTCAGTTCGGAATTGGCCGACGCGATCGAGGTTGTTGGTACTGCCGGGTTGGACCCCGAGGCGAT GGCGGCGACTACTCGCATGACAGGAGTGCCCGCAATATCGTTCGTCCGTGCGGTGCAATCGGCCCTGCCGCCCTCACTGGCGGGTGGATTTCATTTCGGCGCCACCAGTCAAGACATCGT GGATACGGCCCACGCGCTCCAGCTGGCCGAGGCACTCGATATTATAGAAGTCGATTTACACGCCACTGTCAGCGCAATGATGAATCTGGCCGCTGCTCACTGCAATACACCCTGTATCGG GCGCACGGCCTTGCAGCACGCAGCGCCAGTTACGTTCGGCTACAAGGCGTCCGGCTGGTGCGTTGCCCTGGCGGAGCATCTGGTGCAGCTTCCCGCGCTGCGAAAGCGGGTTCTGGTGGC GTCGCTAGGGGGGCCGGTTGGTACCCTTGCCGCGATGGAGGAGCGGGCCGACGCTGTACTGGAGGGTTTCGCTGCGGACCTGGGGTTGGCCATTCCCGCCCTGGCCTGGCACACGCAGCG GGCCCGGATCGTCGAGGTGGCCAGTTGGCTGGCCATATTGCTGGGAATTCTGGCAAAAATGGCCACCGATGTCGTTCACTTGTCCTCCACGGAAGTGCGCGAGCTTTCCGAACCTGTAGC GCCGGGCAGGGGGGGCTCCTCGGCGATGCCTCACAAGCGGAACCCGATTTCCTCGATTACCATCCTGTCCCAGCATGCTGCGGCAGGGGCCCAGCTCTCCATTCTCGTGAACGGCATGGC CAGTCTGCACGAACGTCCGGTGGGGGCGTGGCATTCGGAATGGTTGGCTCTGCCGACGCTGTTCGGCCTTGCCGGCGGTGCCGTGCGCGAGGGCAGGTTTCTGGCCGAGGGGCTGCTGGT CGATGCCGACCAGATGGGTCGCAATCTACAATTGACCAATGGCCTGATTTTCAGCGACGCGGTAGCCGGCCAGTTGGCAAAGCACTTGGGTCGGGCCGAGGCTTATGCCGCTGTCGAGGA TGCCGCCGCCGAGGTGTTGCGTTCAGGCGGCAGCTTTCAGGGTCAGCTGAACCAGCGCCTGCCCGATCACCGCGACGCTATCGCTATTGCTTTTGATACGACGCCGGCGATCCAGGCCGG GGCCGCCCGCTGCCGTAGTGCGCTGGATCATGTGGCTCGTATTCTTGGACCCGCCTCTACCATCGGATTTCAAGGAGGCTAATGACGTGACGACACTGTTTGAGGCGACGACCATCCCGA TTTGCGAGGGCCCGCGCGACCAGACCGCCGAGATCCTTTTCGAGATGCCGCCGGGTGCGTGGGATACCCATTTTCATGTTTTTGGCCCAGTTTCATCGTTTCCATACGCAGAACACAGGC TCTATTCCCCACCGGAGTCGCCACTTGAGGATTATCTGGTGTTGATGGAGGCTTTGGGGATCGAGCGCGGCGTTTGTGTCCATCCGAATGTTCATGGTGCCGACAATTCGGTGACGCTCG ACGCAGTTGCGCGGTCCGATGGTCGTCTGCTGGCGGTGATCAAGCCACATCACGAGATGACTTTTGTTCAGCTGCGGGACATGAAGGCGCAGGGGGTCTGCGGGGTACGTTTTGCCTTCA ATCCGCAGCATGGCTCGGGCGAGTTGGATACTCGTTTGTTCGAGCGTATGTTGGACTGGTGCCGCGACCTAGGCTGGTGCGTAAAATTGCATTTCGCGCCCGCTGCGCTGGACGGTCTGG CTGAACGTTTGGCGCGCGTCGATATTCCGATCATCATCGATCATTTCGGGCGGGTGGACACCGCGCAAGGTGTGGATCAGCCGCACTTCCTGCGTTTGCTCGATCTGGCCAAACTGGACC ATGTCTGGATCAAGCTTACGGGGGCAGATCGTATTAGCGGTTCCGGCGCGCCATATGACGATGTCGTGCCCTTCGCGCACGCTTTGGCAGATGTGGCGCCCGACCGCCTCCTCTGGGGTT CGGATTGGCCGCATTCAGGCTATTTCGATCCGAAGCACATACCCAATGACGGCGACTTGTTGAACCTTTTGGCGCGTTTTGCCCCCGATGCTGAACTGCGTCGTAAGATCCTTGTGGACA ACCCGCAGCGCCTGTTCGGGGCTGCTTGAGGAGCCGAGCCGATGCAACCTTTCGTCTACGAAACAGCCCCAGCGCGCGTCGTTTTCGGGCGCGGCACTTCGCAGAATCTGCGGCGGGAAC TTGAGGCCCTGAATTTTGGCAGGGCGCTGGTTCTTTCCACGCCCGACCAAAAAGAACAATCGCTGCGAATTGCCCAGGGCCTGGGTTCTCAGCTGGCGGGGTCGTTCCACGCCGCTGCCA TGCATACGCCTGTCGAGGTCACCTTGCAGGCGCTTGAGGTGCTGAAGGATGTGCAGGCCGATTGCATCGTGGCGATTGGCGGCGGCTCAACCATTGGGTTGGGCAAGGCACTGGCCCTGC GCACCGATCTGCCGCAGATCGTCGTCCCGACGACTTATGCCGGCTCGGAAATGACGCCGATCCTGGGAGAGACGGAAAACGGGCTGAAGACCACACAGCGTAATCCCAAAGTGCAGCCGA GGGTGGTTCTCTACGATGTGGACCTGACTGTGACGCTTCCGGTGCAGGCCTCGGTTACATCAGGCATGAATGCGATCGCCCATGCGGCCGAGGCATTATATGCGCGGGACGGCAATCCGG TGATCTCGCTGATGGCCGAAGAGGCGATCCGCGCGCTGGCCCATGCCCTGCCGCGTATCGTTGCCACTCCCGACGATATCGAAGCGCGCAGCGATGCCCTCTATGGCGCGTGGCTGTGCG GAACGTGCCTGGGTTCGGCCGGAATGGCGTTGCACCATAAGCTCTGCCACACCCTCGGCGGAAGTTTCGATTTGCCACATGCCCCGACCCACACGGTCATCCTCCCCTATGCGCTCGCCT ATAATAGTGATGCGGCCAGGCCCGCAATGGCAGCCATCGCGCGCGCGCTGGGCATGGCGGATGCAGCGATGGGCATGAGAGCGTTGTCCATGCGGTTGGGCGCCCCGACATCGCTGCGTG AGTTGGGCATGGCAGAAGCCGATCTTGACCGCGCCGCCGACCTGGCCACGCAAAATGCCTATTGGAACCCGCGACCCATCGAGCATGGGCCGATTCGTAACCTTCTGGGACGGGCCTGGG CTGGAACTCCGGTCTGAAGGACCTAGAGGACAGTCAATTCATTGATCTGAAGTCACCAACGAGGAGATATGGGATGAACGAGAACATTGCGATCCGCAAATTGGGCCGCCGACTCCGATT GGGCATTGCCGGTGGCGCGGGTCATTCGCTGATTGGTCCGGTTCACCGGGAGGCGGCTCGGCTTGACGATTTGTTCTCTCTCGATGCTGCGGTGCTGTCCAGTAACGCGGAACGCGGGGA TGCTGAGGCCGCGGCTCTCGGAATTCCGCGCTCCTATTCGTCCACCGCCGAGATGTTCGCAATGGAGAAGGCTAGGCCCGACGGTATTGAGGCCGTTGCCATAGCCACGCCGAATGACAG CCATTACCGGATTCTGTGCGAGGCGCTGGACGCCGGGTTGCATGTAATCTGCGACAAGCCTTTAACCTCCACGAAGGCCGAGGCCGACGACGTGCTGGTGCGGGCGAAGGCCGCGGGCAA GGTTGTGGTCCTGACCCACAATTATTCTGGCTACGCCATGGTACGCCAAGCCCGCGCCATGGTCGCCGCCGGTGAACTTGGGAAAATCCACCAGATTCACGGGGTCTACGCTCTGGGCCA GATGGGCCGTTTGTTCGAGGCCGACGAAGGGGGCGTGCCTCCGGGGATGCGTTGGCGGATTGATCCTGCGCGCGGTGGCGACAGTCACGCCCTGGTGGATATCGGCACCCATGTGCACCA TCTGGCTACCTTCATCACGCAGTTACAGGTCGTTGAGGTAATGGCCGATCTTGGGCCGGCGGTTCAAGGCCGCGCGGCCCATGACAGTGCCAACGTCATGTTCCGTATGGAAAACGGAGC TTTCGGATCGTTCTGGGCCACCAAGGCGGCATCGGGGGCCAGCAAGCTGGCGATCGAAGTCTACGGTGACAAGGGCGGCGTCCTGTGGGAGCAGGCCGACGCCAATAACTTGCTACATAT GCGGCAGGGCCAACCCCCAGCCCTGATTGGTCGACAAGTTGCCGGGCTGCATCCTGCGGCAATCCGCGCGATGCGGGGGCCGGGTTATCATTTCGTGGAAGGCTATCGCGAGGCCTTTGC GAATATGTACGTGGATTTCGCCGAACAGATCTTGGCCATGATGGGCAAGGGGGCCGCAGATCACCTGGCATTGGAAGCGCCGTCGGTCGTGGACGGCCTGCGCTCCATGGCGTTCATCGA AGCCTGTGTGGCGTCGTCGCAGGACCGCCAATGGCGGCAGGTGGAGCAAGTCAGTTGATCTCTCAGCGGCTTCGGCATTTTTCCCGGGCTGGCGGCTCCCCGCAGCTCCCTCCGGTGGAA AGAACGGGTAATCAAAATAATATTCTGATTTTAAAGGATGTTCCAGACAGCTGATTATTCCTGAAATTTAGGGCTCTTTCGGCTGTAGCAATTGACTAAAAGCCGAATTTAAGGGTAATTAAACAAACGCTGTTCGTATTATTTAAACAGGTGAGTGATGGCGATATTCCTGGAAGGCTGGCCGATGGTTTCATCTGAATACCCGGCCAGAAGCGTTGAGGCGCACCCGGCCTATCTGAC GCCAGACTATGTTTTCACGCGAAAGCGTGCGCCGACTCGACCGCTGCGGTTAATTCCTCAGTCTGCGACGGAGCTGTATGGCCCGGTTTATGGACAAGAGAGCGTCCGTCCGGGGGATAA CGACCTGACCCGTCAGCACGAAGCTGAGCCGGTGGGGGAGCGGATTCTGGTGACGGGGCGCGTGACCGACGAAGACGGGCGGGGTGTCCCTAATACGCTGCTAGAGATCTGGCAGGCCAA TGCCGCCGGTCGCTATATCCACAAGCTTGACCAGCATCTTGCCCCGCTTGATCCAAATTTCTCGGGGGCAGGGCGTACGGTTACGGGGGCTGATGGCTCTTATTCCTTCATCACGATCGT GCCGGGCGCCTATCCGGTCGTGGGGCTGCACAATGTCTGGCGCCCGCGCCACATCCATGTGTCGTTGTTCGGTCCGTCCTTCGTGACCCGCTTGGTTACCCAGATATATTTCGAGGGCGA TCCGCTGCTGAAATATGACACGATCTACAACACGGCGCCCGACATCTCGAAGCGCAGCATGGTGGCGCAGTTGGACATGGGCGCCACGCAATCCGAATGGGGCCTGACCTATCGCTTCGA CATCGTTCTGCGTGGGCGCAACGGCAGCTATTTCGAGGAACCCCATGACCACTAAGACCCCACTGACCATCACCCCCTCGCAGACTGTCGGGCCTTTCTATGCCTATTGCCTGACCCCGG AGGACTACGGGACGCTTCCACCGCTGTTCGGCGCGCAGCTTGCGACCGAGGACGCCGAAGGGGAACGGATTACGATCCAGGGAACGATCACGGACGGAGAGGGGGCCATGGTTCCCGATG CCTTGATCGAGATCTGGCAGCCGGACGGGCAGGGGCGTTTTGCTGGAGCCCATCCAGAGCTGCGGAATTCGGCCTTCAAGGGCTTCGGGCGCCGCCACTGTGACAAAAGCGGAAACTTCA GTTTCCAAACCGTGAAGCCTGGCCGGGTGCCCACTGCCGACGGCGTGATGCAGGCACCCCATATCGCTTTGTCGATCTTCGGCAAGGGATTGAACCGCCGGCTCTATACGCGGATCTACT TCGCAGACGAGGCATCGAATGCCGAGGACCCCGTTCTGTCGATGCTGTCCGAGGATGAGCGCGTGACCCTGATCGCCACCTCTGAATCGCCCGCCGCATATCGCCTCGACATCCGCCTGC AAGGCGACGGCGAAACGGTGTTTTTCGAGGCCTGAGTCGGCCGGCAAGTTTGCGGGGATCCGTCCGCCGCAATTGTGTTTCGCTATAGACGCCACGGCTGCCGCATGCCGCCGGGTGGAA GGGCCTTGCAAGGCCTGTCAACGGCGGAGTAAAATCCGGCCAGGCGGCGGAGTAAAACCAGGCCACTTGTGGCCCACGCATGAGACACCCGGGAGGGCGTAGCCCAAGCGGGGGTCTCAT GCGTGTGCGGCGGTTTTCTGGGGGTTCAGCCAGCCTTGCGGGCGCGGCTTTGAGCGAGACGATAGCTGTCGCCGTTCATCTCGAG

93
HONNAN LEHET TUDNI, HOGY GÉNT TALÁLTUNK?
1. DNS szekvencia homológia alapján Adatbankok, FASTA, BLAST 2. ORF KERESÉS, Általában ATG-vel kezdődő szakaszokat keresünk, amelyek ésszerűen hosszúak (100 aminosav) Clone Manager 3. A kapott ORF-k homológia kutatása
 Clone Manager. 3. A kapott ORF-k homológia kutatása.")




















 prokariótában>")












 Bihari Péter.>")