Előadást letölteni
1
A minimális sejt és az anyagcsere autokatalitikus magja
Biokémia II – Anyagcsere Dr. Kun Ádám, Ph.D. okleveles biológus, okleveles vegyész Itt is felhívom a kedves hallgatók figyelmét arra, hogy képleteket, teljes reakcióutakat NEM kérdezek vissza. Ha például felteszem a kérdést, hogy mire jó a glikolízis? Akkor, arra azt kell válaszolni, hogy anaerob energiatermelesre (ATP termelés). S nem kell felírni a lépéseit.
. S nem kell felírni a lépéseit.")
2
Az RNS Világ

3
A DNS / fehérje világ előtt…
Ma élő élőlényekben Az információDNS-ben tárolódik Fehérjék az enzimek Kódolja Fehérje DNS Replikál Monomert állít elő

4
RNS enzimként is működhet és információt is tárolhat!
… egy RNS világ volt RNS enzimként is működhet és információt is tárolhat! RNS Replikálódik, hogy enzimet kapjunk Reakciókat katalizál, monomert állít elő Cech és munkatársai (Kruger, K. et al. Self-splicing RNA: autoexcision and autocyclization of the ribosomal RNA intervening sequence of Tetrahymena. Cell 31, (1982)) és nem sokkal később Altman és munkatársai is (Guerrier-Takada, C., Gardiner, K., Marsh, T., Pace, N. & Altman, S. The RNA moiety of ribonuclease P is the catalytic subunit of the enzyme. Cell 35, (1983)) olyan RNS molekulát írtak le, amely kémiai reakciót katalizál. Az első esetben a Tetrahymena nukleuszában a riboszómális RNS érésénél egy intron fehérje enzim nélkül vágódott ki az RNS láncból. A másik esetben RNáz-P-ről derült ki, hogy a katalízist az RNS rész végzi. Ezek a felfedezések lettek az alapjai az RNS világ hipotézisnek (Gilbert, W. The RNA world. Nature 319, 618 (1986)) DNS stabilabb és a fehérjék jobb enzimek
) és nem sokkal később Altman és munkatársai is (Guerrier-Takada, C., Gardiner, K., Marsh, T., Pace, N. & Altman, S. The RNA moiety of ribonuclease P is the catalytic subunit of the enzyme. Cell 35, (1983)) olyan RNS molekulát írtak le, amely kémiai reakciót katalizál. Az első esetben a Tetrahymena nukleuszában a riboszómális RNS érésénél egy intron fehérje enzim nélkül vágódott ki az RNS láncból. A másik esetben RNáz-P-ről derült ki, hogy a katalízist az RNS rész végzi. Ezek a felfedezések lettek az alapjai az RNS világ hipotézisnek (Gilbert, W. The RNA world. Nature 319, 618 (1986)) DNS stabilabb és a fehérjék jobb enzimek.")
5
RNS világ bizonyítékai a mai szervezetek biokémiájában I
Természetes RNS enzimek (ribozim) Mindegyik RNS hasítást katalizál I. Csoportbeli intronok II. Csoportbeli intronok RNáz P Kalapácsfej Hajtűhurok Hepatitis Delta Vírus Neurospora Varkund Satelite RNA A természetes ribozimeknek napjainkig 7 csoportját izolálták. A természetes ribozimek (összefoglaló Doudna, J. A. & Cech, T. R. The chemical repertoire of natural ribozymes. Nature 418, (2002)) mindegyike RNS hasítást végez, bár különböző mechanizmussal (Westhof, E. Chemical diversity in RNA cleavage. Science 286, (1999); Doherty, E. A. & Doudna, J. A. Ribozyme structure and mechanism. Annual Review of Biochemistry 69, (2000); Doudna, J. A. & Lorsch, J. R. Ribozyme catalysis: not different just worse. Natue structural & molecular biology 12, (2005)). Joyce, G. (2002) Nature 418: alapján
 Mindegyik RNS hasítást katalizál. I. Csoportbeli intronok. II. Csoportbeli intronok. RNáz P. Kalapácsfej. Hajtűhurok. Hepatitis Delta Vírus. Neurospora Varkund Satelite RNA. A természetes ribozimeknek napjainkig 7 csoportját izolálták. A természetes ribozimek (összefoglaló Doudna, J. A. & Cech, T. R. The chemical repertoire of natural ribozymes. Nature 418, (2002)) mindegyike RNS hasítást végez, bár különböző mechanizmussal (Westhof, E. Chemical diversity in RNA cleavage. Science 286, (1999); Doherty, E. A. & Doudna, J. A. Ribozyme structure and mechanism. Annual Review of Biochemistry 69, (2000); Doudna, J. A. & Lorsch, J. R. Ribozyme catalysis: not different just worse. Natue structural & molecular biology 12, (2005)). Joyce, G. (2002) Nature 418: alapján.")
6
RNS világ bizonyítékai a mai szervezetek biokémiájában II: Koenzimek
Koenzimek: valamilyen specifikus kémiai csoport átadásában résztvevő metabolitok (rengeteg reakcióban) Acetyl koenzim A (koenzim A): acetyl csoport NADH, FADH2 (NAD+, FAD): hidrogén és elektron ATP (ADP): nagyenergiájú foszfát NADP FAD NAD CoA ATP
 Acetyl koenzim A (koenzim A): acetyl csoport. NADH, FADH2 (NAD+, FAD): hidrogén és elektron. ATP (ADP): nagyenergiájú foszfát. NADP. FAD. NAD. CoA. ATP.")
7
RNS világ bizonyítékai a mai szervezetek biokémiájában III
Dezoxi-ribonukleotidok ribonukleotidokból keletkeznek (de ezt valószínűleg ribozimek nem tudják katalizálni). Riboswitches: Génreguláció vélhetően legősibb formája. Az mRNS térszerkezete határozza meg, hogy lefordítódik fehérjévé. Nukleozid-difoszfátot a ribonukleotid-reduktáz enzim redukálja dezoxi-nukleozid-difoszfáttá.
. Riboswitches: Génreguláció vélhetően legősibb formája. Az mRNS térszerkezete határozza meg, hogy lefordítódik fehérjévé. Nukleozid-difoszfátot a ribonukleotid-reduktáz enzim redukálja dezoxi-nukleozid-difoszfáttá.")
8
RNS világ bizonyítékai a mai szervezetek biokémiájában IV: Transzláció
mRNS tRNS riboszóma Mesterségesen előállított ribozimekkel a fehérjeszintézis megoldható. A transzkripciót az RNS-polimeráz enzim végzi. A DNS fehérje „átmentet” (transzláció) RNS közvetítésével történik Riboszómában a peptidil transzfert egy ribozim végzi!
 RNS közvetítésével történik. Riboszómában a peptidil transzfert egy ribozim végzi!")
9
RNS világ lehetőségei:
RNS szintézis: Nukleotid képzés pirimidinből és aktivált ribózból; Polinukleotidok 5’ foszforilációja; 5’ foszfát aktiválása 5’,5’ pirofoszfát kötött nukleotid kapcsolással; Ligáz aktivitás; Legjobb szintetáz kb. 200 bázis hosszú és 14 nuklotidot tud egy templát alapján hozzákapcsolni egy oligonukleotid lánchoz 97.5%-os másolási hűséggel. Protein szintézis: Minden lépés megoldható. Aminosavak aktiválása (sokféle aminoaciláció); Peptid kötés kialakítás (peptidil transzfer) Membrán transzfer Redoxi reakciók (NAD függő alkohol dehidrogenáz) Egyéb reakciók (amid kötés bontás, alkiláció, porfirin metiláció, kén alkiláció, Diels-Alder cikloaddíció, amid kötés kialakítás, hidas bifenil izomeráció) Ezeket az RNS enzimeket mind in vitro evolúcióval állították elő.
; Peptid kötés kialakítás (peptidil transzfer) Membrán transzfer. Redoxi reakciók (NAD függő alkohol dehidrogenáz) Egyéb reakciók (amid kötés bontás, alkiláció, porfirin metiláció, kén alkiláció, Diels-Alder cikloaddíció, amid kötés kialakítás, hidas bifenil izomeráció) Ezeket az RNS enzimeket mind in vitro evolúcióval állították elő.")
10
Információ replikáció Eigen Paradoxona

11
Információ replikáció
A replikáció nem hibátlan (főleg nem replikáz és javító mechanizmusok nélkül) A másolás pontossága korlátozza a fenntartható információ hosszát. Mekkora információ (milyen hosszú RNS szál) tartható meg adott másolási pontosság mellett?
 A másolás pontossága korlátozza a fenntartható információ hosszát. Mekkora információ (milyen hosszú RNS szál) tartható meg adott másolási pontosság mellett")
12
„Replikáció” egy példája
RNA RGA RNX RNH DNM RQA RNJ RPA WORLD WORLF WORLL IDRYD KORLD WERLD WORUD WORHD WORWD WRRLD HYPOTHESIS EYPKTHYSII HYPEXHESIS HYPETHESKS HYYOTHESIS HYPOSHESIS HYPOTMESIS HTPOTHESIS CYPOTGESIS HYPOTHEGIA HYPOXHLSIS HYPXTHESIS HYPUTHESIS

13
Eigen Pradoxona és a hibaküszöb
Nincs enzim nagy genom nélkül, és nincs nagy genom enzim nélkül Az Eigen féle összefüggés azt mondja ki, hogy a fenntartható információ hossza (N) fordítottan arányos a hibarátával (1-q). A modell alapfeltevése, hogy az információt hordozó vad típusú molekula replikációs rátája a legmagasabb (tehát az szaporodik a leggyorsabban, ez az ő előnye). Az ln(s) mutatja, hogy mennyivel jobb a vad típus a mutánsainál. Általában ls(s)=1-nek veszik, így 99% másolási hűség mellett (1% hiba) 100 hosszú enzim tartható fent. Az 1% hibarátánál valószínűleg sokkal magasabb hibarátával kell számolnunk azon körülmények között, tehát optimista becslés mellett is alacsony a fenntartható genomméret. Ebből ered az Eigen paradoxon. A genom méret növelhető a másolási pontosság növelésével. Ehhez enzim kell, ami viszont nem fér bele a 100-as hosszba, ami megengedhető. Tehát a nagyobb genomhoz jó enzim kell, amihez viszont hosszú genom, hogy kódolja. Ez egy tipikus 22-es csapdája. A jobb oldali ábra a hibakatasztrófát ábrázolja. Az Eigen modell (az egyenletek ideillesztésétől megkímélnélek titeket) eredménye, hogy egy adott hiba felett a másolás nem megkülönböztethető a véletlenszerűtől, akkor sem, ha a vad típus másolási sebessége gyorsabb. Ez a hibaküszöb (error threshold). N hossz lns a mesterkópia szelekciós fölénye q másolási pontosság Swetina és Schuster 1998 alapján
 fordítottan arányos a hibarátával (1-q). A modell alapfeltevése, hogy az információt hordozó vad típusú molekula replikációs rátája a legmagasabb (tehát az szaporodik a leggyorsabban, ez az ő előnye). Az ln(s) mutatja, hogy mennyivel jobb a vad típus a mutánsainál. Általában ls(s)=1-nek veszik, így 99% másolási hűség mellett (1% hiba) 100 hosszú enzim tartható fent. Az 1% hibarátánál valószínűleg sokkal magasabb hibarátával kell számolnunk azon körülmények között, tehát optimista becslés mellett is alacsony a fenntartható genomméret. Ebből ered az Eigen paradoxon. A genom méret növelhető a másolási pontosság növelésével. Ehhez enzim kell, ami viszont nem fér bele a 100-as hosszba, ami megengedhető. Tehát a nagyobb genomhoz jó enzim kell, amihez viszont hosszú genom, hogy kódolja. Ez egy tipikus 22-es csapdája. A jobb oldali ábra a hibakatasztrófát ábrázolja. Az Eigen modell (az egyenletek ideillesztésétől megkímélnélek titeket) eredménye, hogy egy adott hiba felett a másolás nem megkülönböztethető a véletlenszerűtől, akkor sem, ha a vad típus másolási sebessége gyorsabb. Ez a hibaküszöb (error threshold). N hossz. lns a mesterkópia szelekciós fölénye q másolási pontosság. Swetina és Schuster 1998 alapján.")
14
Szekvencia vs. Szerkezet
Átíródik A szekvenciát kell megtartani Mutáns: rátermettség 0 DNS fehérje GCATGACTCATATGC ATCGTCTGTCGGCGAT Structure has to be maintained Mutáns: Azonos rátermettség RNS AUCGUCUGUCGGCGAU GCAUGACUCAUUAUGC

15
1D-2D-3D szerkezet AAACAGAGAAGUCAACCAGAGAAACACACGUUGUGGUAUAUUACCUGGUA

16
RNS szerkezet Az enzimaktivitás a szerkezettől függ
A ribozim fenotípusa a szerkezete Kevesebb szerkezet van, mint szekvencia Egy kevés mutáció általában nem változtatja meg a szerkezetet Szerkezet könnyebben fenntartható, mint a szekvencia. (fenotipikus hibaköszöb)
")
17
Az élet kódja: Transzláció A genetikai kód eredete

18
Transzláció eredete Fehérjék jobb katalizátorok (4 kémiailag hasonló bázis vs. 20 kémiailag sokféle aminosav) Mivel az RNS központi szerepet játszik a transzlációban, így valószínűleg az RNS világban „találták fel”

19
Kódtábla genetikai kód - tulajdonképpen egy jelkulcs. Három bázisból álló csoport (kodon), mely egy adott aminosavat kódol kodon - 3 bázis alkotta csoport a nukleinsavakban, melyek 1-1 aminosavat kódolnak, vagy start ill. stop jelként funkcionálnak a gének "leolvasásakor".Gyakran tripletként is emlegetik őket. Az univerzális kód, például mitokondriumban sem a fentinek megfelelő. Itt például a UGA STOP helyett Triptofán jelent, az AUA Izoleucin helyett metionin. Stb. Több más verziója is lehet a genetikai kódnak, azonban ezek mind csak dialektusok, azaz csak kevés helyen térnek el a legelterjedtebb variánstól. Érdekesség: Az egyes aminosavak kodonhoz rendelése nem véletlenszerű, hanem evolúciósan a minél kisebb mutációs teherre optimalizált. Egy bázis megváltozása – legtöbbször - vagy egyáltalán nem okoz aminosav változást (általában az utolsó kodoné), vagy nem okoz hidrofobicitás változást. Bár szokás abból levezetni a tripletet, hogy a 20 aminosav kódolásához 3 bázis kell (helyenként ugye 4 lehetséges bázissal), ez visszafele gondolkodás. Ha több bázissal kapcsolódna a tRNS az mRNS-hez (mondjuk kvartettel), akkor a két molekula kötése túl erőse lenne. Ha kevesebbel (mondjuk 1 v. 2 bázissal), akkor a kötés túl gyenge lenne. Persze elképzelhető 1 v. 2 bázist alkalmazó kód, de továbbra is triplettel kapcsolódna a tRNS az mRNS-hez.
, vagy nem okoz hidrofobicitás változást. Bár szokás abból levezetni a tripletet, hogy a 20 aminosav kódolásához 3 bázis kell (helyenként ugye 4 lehetséges bázissal), ez visszafele gondolkodás. Ha több bázissal kapcsolódna a tRNS az mRNS-hez (mondjuk kvartettel), akkor a két molekula kötése túl erőse lenne. Ha kevesebbel (mondjuk 1 v. 2 bázissal), akkor a kötés túl gyenge lenne. Persze elképzelhető 1 v. 2 bázist alkalmazó kód, de továbbra is triplettel kapcsolódna a tRNS az mRNS-hez.")
20
Néhány tény a genetikai kódról
Közel univerzális Redundáns Miért triplet? A triplet optimális a reverzibilis kapcsolódáshoz Miért 20 aminosav? Az enzim sokféleség növekszik a több aminosavval, de a mutációs robusztusság csökken. A kód optimálizált mutációs robosztusságra

21
A megfejtetlen rejtély
Oligo-nukleotidok ásványi felszínen nukleotidok RNS ribozimok

22
A megfejtetlen rejtély
Kódolja fehérje RNS DNS Membrán monomert állít elő replikálja monomert állít elő

23
Anyagcsere

24
Minimális sejt

25
Minimum sejt Top – down: Meglevő szervezetek genomjából indulunk ki
Bottom – up: Minimális funkciók, élő sejt szintézise

26
Minimum sejt – felülről lefelé
A gének legkisebb lehetséges halmaza, amivel fenntartható egy működő sejt a legjobb körülmények között (minden forrás rendelkezésre áll, nincs környezeti stressz) Bioinformatika Knock-out kísérletek 206 gén A minimális sejt témában két tanulmányt emelnék ki: Gil, R., Silva, F. J., Peretó, J. & Moya, A Determination of the core of a minimal bacterial gene set. Microbiol Mol Biol Rev. 68, Gabaldón, T., Peretó, J., Montero, F., Gil, R., Latorre, A. & Moya, A Structural analyses of a hypothetical minimal metabolism. Philosophical Transaction of the Royal Society of London, accepted
 Bioinformatika. Knock-out kísérletek. 206 gén. A minimális sejt témában két tanulmányt emelnék ki: Gil, R., Silva, F. J., Peretó, J. & Moya, A Determination of the core of a minimal bacterial gene set. Microbiol Mol Biol Rev. 68, Gabaldón, T., Peretó, J., Montero, F., Gil, R., Latorre, A. & Moya, A Structural analyses of a hypothetical minimal metabolism. Philosophical Transaction of the Royal Society of London, accepted.")
27
Konzerválódott fehérjék
Legjobban a transzláció és az RNS polimerázok konzerválódtak Metabolizmusban viszont kevés konzerválódott (több genom összehasonlításában) enzim van A konzerváltság nem jelenti, hogy laborban elengedhetetlen (kilőhető). Például repair nélkül a sejt jól él, de fennmaradhat-e?
 enzim van. A konzerváltság nem jelenti, hogy laborban elengedhetetlen (kilőhető). Például repair nélkül a sejt jól él, de fennmaradhat-e")
28
Minimum sejt – funkciók I
DNS metabolizmus (replikáció, módosítás, javítás és hasítás) 16 gén RNS metabolizmus (transzkripció, transzláció, RNS degradáció) 106 gén Fehérje feldolgozás (módosítás, felgombolyodás, áthelyezés, lebontás) 15 gén Sejtszintű működés (osztódás, transzport) 5 gén
 16 gén. RNS metabolizmus (transzkripció, transzláció, RNS degradáció) 106 gén. Fehérje feldolgozás (módosítás, felgombolyodás, áthelyezés, lebontás) 15 gén. Sejtszintű működés (osztódás, transzport) 5 gén.")
29
Minimum sejt – funkciók II
Köztes metabolizmus és energetika Glikolízis (10 gén) ATP szintézis, H+ gradiens (9 gén) Pentóz-foszfát út (3 gén) Lipid metabolizmus (7 gén) Nukleotid bioszintézis (15 gén) Kofaktor bioszintézis (12 gén) Egyéb (8 gén)
 ATP szintézis, H+ gradiens (9 gén) Pentóz-foszfát út (3 gén) Lipid metabolizmus (7 gén) Nukleotid bioszintézis (15 gén) Kofaktor bioszintézis (12 gén) Egyéb (8 gén)")
31
Az Élet Szikrája az anyagcsere autokatalitikus magja
Mitől több – ha több - az élő sejt egy zsák enzimnél és némi DNS-nél? Mitől lesz élő egy sejt?

32
Gánti kemoton elmélete

33
Nincsen kis molekulasúlyú metabolit!
Egy zsák enzim és DNS Legyen egy sejtünk, amiben csak enzimek és DNS (RNS) van. A környezetben minden tápanyag jelen van, amit az élőlény fel tud venni (optimális mennyiségben) Enzimek Nincsen kis molekulasúlyú metabolit! DNA/RNA H2O, H+, CO2, Fe2+, CNO, NO2-, NO3-, HPO42-, SO42-, O2, cukrok, aminosavak, alkoholok
 van. A környezetben minden tápanyag jelen van, amit az élőlény fel tud venni (optimális mennyiségben) Enzimek. Nincsen kis molekulasúlyú metabolit! DNA/RNA. H2O, H+, CO2, Fe2+, CNO, NO2-, NO3-, HPO42-, SO42-, O2, cukrok, aminosavak, alkoholok.")
34
??? Egy zsák enzim és DNS Működőképes-e ez a sejt?
Kell-e valaminek eleve a sejtben lennie, hogy a metabolizmus beinduljon? Enzimek ??? DNA/RNA H2O, H+, CO2, Fe2+, CNO, NO2-, NO3-, HPO42-, SO42-, O2, cukrok, aminosavak, alkoholok

35
Elméleti kísérlet! Legyen egy reakcióhálózatunk
Adjuk meg a felvehető anyagok listáját Mi termelhető meg a reakciólista alapján? Ha minden megtermelhető, akkor az élő sejt nem több egy zsák enzimnél. Ha nem termelhető meg minden, akkor mi kell még?

36
Reakcióhálózatok Eubacteria Archea Eukarióta Minimális metabolizmus
Escherichia coli Heliobacter pylori Staphylococcus aureus Lactococcus lactis Streptomyces coelicolor Geobacter sulfurreducens Synechocystis Archea Methanosarcina barkeri Eukarióta Saccharomyces cerevisiae Minimális metabolizmus

37
ATP Escherichia coli Reakció szám: 931 Metabolitok száma: 761
Megtermelhető metabolitok: 692 Külső molekulák: 53 Makromolekulák: 3 Külső molekulákból megtermelhető: 315 Hozzáadandó: ATP

38
Az anyagcsere autokatalitikus magja
Escherichia coli Heliobacter pylori Staphylococcus aureus Lactococcus lactis Streptomyces coelicolor ATP ATP ATP ATP ATP

39
Az anyagcsere autokatalitikus magja
Methanosarcina barkeri Geobacter sulfurreducens Synechocystis ATP + NAD ATP + NAD + THF + CoA ATP + NAD + THF + CoA + cukor

40
Autokatalitikus molekulák
ATP NAD (NADP) Koenzim A THF Kinon Cukor ATP NAD CoA
 Koenzim A. THF. Kinon. Cukor. ATP. NAD. CoA.")
41
ATP = energia eleve kell a szervezeteknek, hogy éljenek.
Az Élet Szikrája ATP = energia eleve kell a szervezeteknek, hogy éljenek.

42
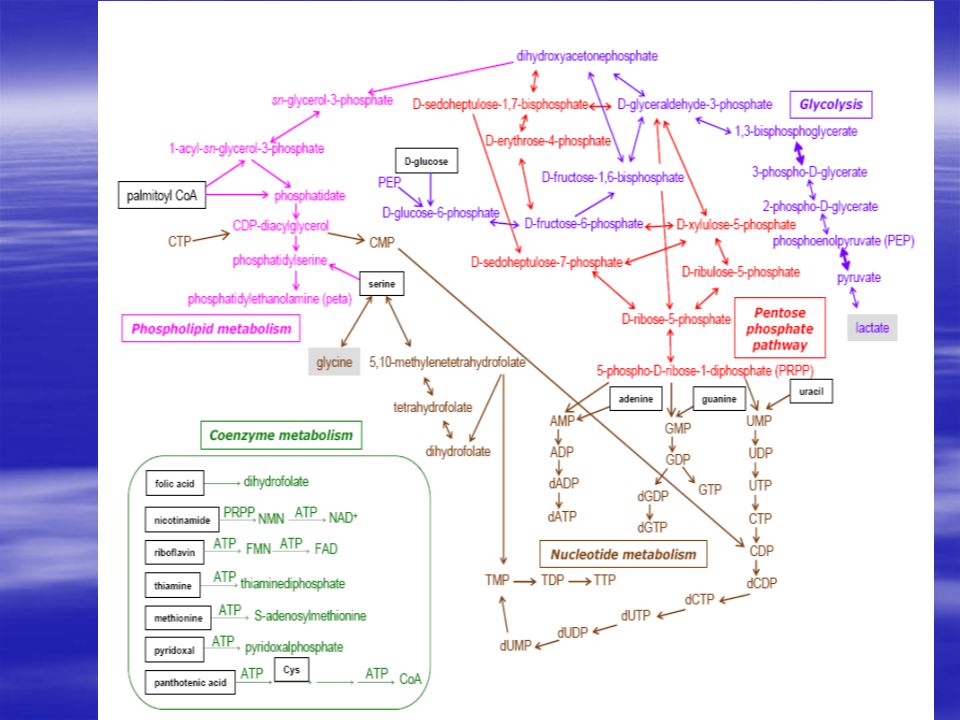
Anyagcsere főbb útvonalai

43
Anyagcsere főbb útvonalai
Szénhidrát metabolizmus Glikolízis Pentóz foszfát út Citromsav ciklus Energia metabolizmus Oxidatív foszforiláció Calvin ciklus Lipid metabolizmus Nukleotid bioszintézis Purin metabolizmus Pirimidin metabolizmus Aminosav metabolizmus Nucleotid Salvage Pathway Kofaktor bioszintézis

44
Glikolízis A glükóz lebontásának van egy olyan szakasza (a glikolízis), amely nem igényel oxigént, és akkor is működik, amikor a sejtek oxigénhiányos környezetbe kerülnek. Ilyenkor a lebontás csak piroszőlősavig történik, amelyből tejsav keletkezik, és eközben 1 molekula glükózból 2 molekula ATP keletkezik. Ez egy ősi útvonal, minden sejt képes rá, az olyan sejtek, amelyekben nincs mitokondrium - mint például a vörösvértestek -, kizárólag ily módon jutnak a glükózból energiához.
, amely nem igényel oxigént, és akkor is működik, amikor a sejtek oxigénhiányos környezetbe kerülnek. Ilyenkor a lebontás csak piroszőlősavig történik, amelyből tejsav keletkezik, és eközben 1 molekula glükózból 2 molekula ATP keletkezik. Ez egy ősi útvonal, minden sejt képes rá, az olyan sejtek, amelyekben nincs mitokondrium - mint például a vörösvértestek -, kizárólag ily módon jutnak a glükózból energiához.")
45
Pentóz-foszfát út A pentóz foszfát út során az egyes 4,5,6 szénatomos cukrok egymásba alakulhatnak, illetve a 6 szénatomos cukrokból 5 szénatomos cukrok keletkeznek. A szervezet pentóz raktárat (pool) tarthat fent így.
 tarthat fent így.")
46
Citromsav ciklus Feltételezések szerint ez a legősibb ciklus.

47
Lipid metabolizmus ACP – acil-hordozó-fehérje

48
Calvin ciklus A CO2 fixálás alapfolyamata az ún. Calvin ciklus, ami a növények jelentős részében a CO2 megkötés Melvin Calvin és mtsai. által feltárt kizárólagos mechanizmusa. A Calvin ciklus folyamán keletkezõ elsõdleges termékek 3 szénatomot tartalmaznak, ezért a széndioxidot ezen mechanizmus alapján kötõ növényeket C3-as növényeknek nevezzük. A Calvin ciklus kulcsenzime a Rubisco (ribulóz-1,5-biszfoszfát-karboxiláz-oxigenáz), ami a tilakoid membrán sztróma felöli oldalához tapad (azaz nincs a membránba ágyazódva). A ciklus elsõ lépésében a ribulóz 1,5-biszfoszfát (RuBP) CO2-t köt meg, majd egy 6 szénatomos átmeneti termék képzõdése után 2 molekula glicerinsav-3-foszfátra (3PG) esik szét. Ezek egy-egy ATP felhasználásával glicerinsav-1,3-biszfoszfáttá alakulnak. Ezt a folyamatot a 3-foszfoglicerát kináz enzim katalizálja. A folyamat következõ lépésében glicerinaldehid-3-foszfát (G3P) keletkezik, NADPH felhasználásával. Így a megkötött széndioxid szénhidrát szintig redukálódott. A folyamat során 6 CO2 molekula megkötésének eredményeként 12 molekula glicerinaldehid-3-foszfát keletkezik. Közülük kettõ továbbalakul glukózzá, aminek átmeneti terméke a fruktóz-1,6-biszfoszfát. A többi, ATP felhasználásával a Rubisco regenerálására fordítódik.
, ami a tilakoid membrán sztróma felöli oldalához tapad (azaz nincs a membránba ágyazódva). A ciklus elsõ lépésében a ribulóz 1,5-biszfoszfát (RuBP) CO2-t köt meg, majd egy 6 szénatomos átmeneti termék képzõdése után 2 molekula glicerinsav-3-foszfátra (3PG) esik szét. Ezek egy-egy ATP felhasználásával glicerinsav-1,3-biszfoszfáttá alakulnak. Ezt a folyamatot a 3-foszfoglicerát kináz enzim katalizálja. A folyamat következõ lépésében glicerinaldehid-3-foszfát (G3P) keletkezik, NADPH felhasználásával. Így a megkötött széndioxid szénhidrát szintig redukálódott. A folyamat során 6 CO2 molekula megkötésének eredményeként 12 molekula glicerinaldehid-3-foszfát keletkezik. Közülük kettõ továbbalakul glukózzá, aminek átmeneti terméke a fruktóz-1,6-biszfoszfát. A többi, ATP felhasználásával a Rubisco regenerálására fordítódik.")
49
Anyagcsere autokatalitikus magja

50
Ajánlott irodalom John Maynard-Smith és Szathmáry Eörs: Az evolúció nagy lépései. Scientia, Budapest, 1997 Bálint Miklós: Molekuláris biológia I-II. Műszaki könyvkiadó, Budapest, 2000 Ádám Veronika (szerkesztő): Orvosi biokémia. Semmelweis, Budapest, 1996 Láng Ferenc (szerk.): Növényélettan. Eötvös Kiadó
: Orvosi biokémia. Semmelweis, Budapest, Láng Ferenc (szerk.): Növényélettan. Eötvös Kiadó.")



















>")
