Előadást letölteni
1
Sejtalkotók és citoplazma

2
Élet, élő anyag, sejt Minden ma élő szervezet sejtes felépítésű
Sejt csak sejtből származik: ma nem keletkeznek új szervezetek Minden ma élő szervezet közös ősből származik információ hordozó molekulák szekvencia hasonlóság rokonsági kapcsolatok kiszámíthatók működésbeli egyezések Szervezettség alapján: prokarióták és eukarióták egyaránt fejlett szervezetek közös ős, eltérő irányú adaptáció

3
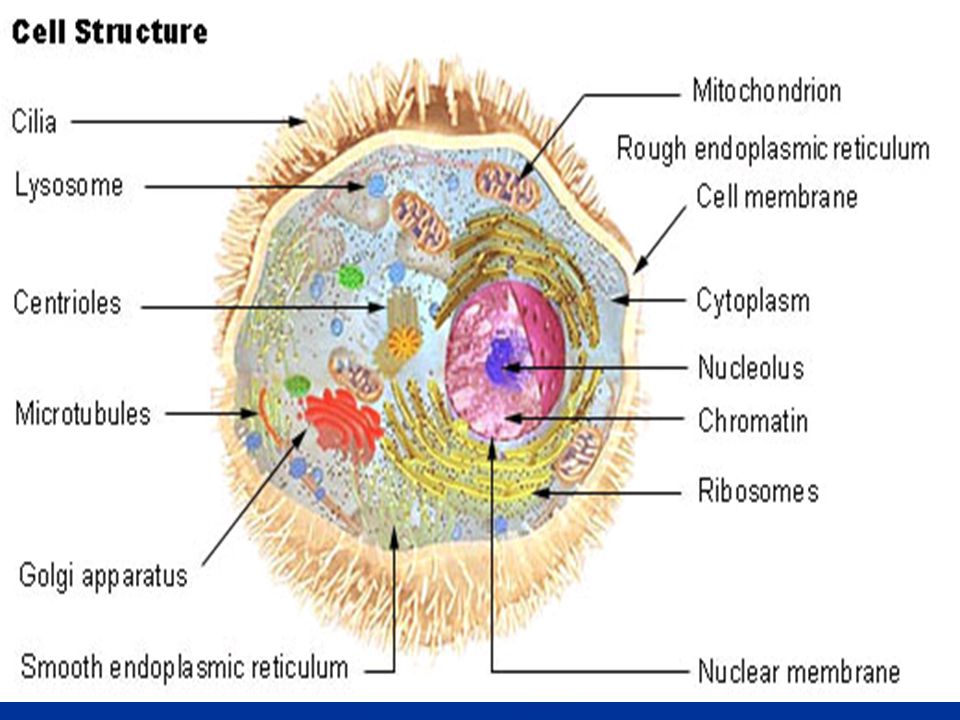
A sejt felépítése 1. Bevezetés
A sejteket fénymikroszkóppal vizsgálva csak kismértékű strukturáltságot figyelhetünk meg. Látható a sejt alakja, sejthártya, sejtmag és a citoplazma. Speciális festési eljárásokkal egy-egy nagyobb – pl. mitokondrium –még láthatóvá tehető. A legegyszerűbb felépítésű sejtek a prokariótákra jellemzőek. A sejteket határoló hártya (plazmamembrán) választja el a külvilágtól. A sejten belül viszont nincsenek membránnal határolt terek. Az eukarióta sejtek sokkal nagyobbak, működésükben pedig sokkal bonyolultabbak. Belsejében sokféle speciális összetételű, különböző működésű membrán tagolja a sejt plazmáját.
 választja el a külvilágtól. A sejten belül viszont nincsenek membránnal határolt terek. Az eukarióta sejtek sokkal nagyobbak, működésükben pedig sokkal bonyolultabbak. Belsejében sokféle speciális összetételű, különböző működésű membrán tagolja a sejt plazmáját.")
4
A prokarióta sejt A sejthártyával határolt citoplazmában találhatók az információhordozó molekulák és a riboszómák. Nincsenek sejtszervecskék, sejtváz és citoplazmás membránok. Valamennyi kémiai reakció egyetlen térben zajlik A tömeghez képest óriási felszín – jó körülmények között - igen gyors anyagcserét és szaporodást tesz lehetővé. A prokarióták nagyon alkalmazkodóképesek gyors nemzedékek, nagy változékonyság, örökítőanyag cseréje A sérülékeny sejthártyát (többrétegű) sejtfal, tok, kapszula, vastag nyálkaréteg védheti fizikai, kémiai és biológiai behatásoktól
 sejtfal, tok, kapszula, vastag nyálkaréteg védheti fizikai, kémiai és biológiai behatásoktól.")
5
Prokarióta sejt felépítése

6
A prokarióta sejt: formái

7
Az eukarióta (állati) sejt
Többsejtű (valódi szövetes) állatok sejtjei, Különböző méretű, típusú és feladatú sejtek Több ezerszer nagyobbak a prokarióta sejteknél A nagy belső tér nem egységes, eltérő kémiai/fizikai körülmények Citoplazmás membránok tagolják : kompartmentalizáció Az elválasztott terek között szervezett kommunikáció és anyagcsere Sejtváz: architektúra és szállítási útvonalak, mozgás és alakváltoztatás A genetikai információ tárolása, működése külön térben: sejtmag Szállítás (megmunkálás) és kézbesítés: vezikuláris transzport, szignál szekvenciák és G fehérjék Endoszimbionta organellumok
 állatok sejtjei, Különböző méretű, típusú és feladatú sejtek. Több ezerszer nagyobbak a prokarióta sejteknél. A nagy belső tér nem egységes, eltérő kémiai/fizikai körülmények. Citoplazmás membránok tagolják : kompartmentalizáció. Az elválasztott terek között szervezett kommunikáció és anyagcsere. Sejtváz: architektúra és szállítási útvonalak, mozgás és alakváltoztatás. A genetikai információ tárolása, működése külön térben: sejtmag. Szállítás (megmunkálás) és kézbesítés: vezikuláris transzport, szignál szekvenciák és G fehérjék. Endoszimbionta organellumok.")
8
A növényi sejt

9
Az „állati” sejt

10
Prokarióták Eukarióták
Prokarióták Eukarióták Jellemző élőlények baktériumok, archeák protisták, gombák, növények, állatok Általános méret ~ 1-10 µm ~ µm (a spermiumok, a farokrésztől eltekintve, kisebbek) A sejtmag nukleoid régió; nincs igazi sejtmag valódi sejtmag kettős membránnal körülvéve DNS körkörös (általában) lineáris molekulák (kromoszómák hiszton fehérjékkel) RNS- és fehérjeszintézis a citoplazmában zajlik az RNS-szintézis a magban fehérjeszintézis a citoplazmában Riboszómák 50S+30S = 70S 60S+40S = 80S Citoplazmatikus szerkezet kevésbé szervezett magasan szervezett, endomembránokkal és citoszkeletonnal Sejtmozgás flagellinből felépülő flagellumok flagellumok és tubulinból felépülő ostorok Mitokondrium nincs 1-től néhány tucatig (van, ahol hiányzik) Színtestek algákban és növényekben Organizáció általában egysejtűek egysejtűek, kolóniák, magasabb rendű többsejtű szervezetek specializált sejtekkel Sejosztódás hasadás mitózis meiózis
 A sejtmag. nukleoid régió; nincs igazi sejtmag. valódi sejtmag kettős membránnal körülvéve. DNS. körkörös (általában) lineáris molekulák (kromoszómák hiszton fehérjékkel) RNS- és fehérjeszintézis. a citoplazmában zajlik. az RNS-szintézis a magban fehérjeszintézis a citoplazmában. Riboszómák. 50S+30S = 70S. 60S+40S = 80S. Citoplazmatikus szerkezet. kevésbé szervezett. magasan szervezett, endomembránokkal és citoszkeletonnal. Sejtmozgás. flagellinből felépülő flagellumok. flagellumok és tubulinból felépülő ostorok. Mitokondrium. nincs. 1-től néhány tucatig (van, ahol hiányzik) Színtestek. algákban és növényekben. Organizáció. általában egysejtűek. egysejtűek, kolóniák, magasabb rendű többsejtű szervezetek specializált sejtekkel. Sejosztódás. hasadás. mitózis meiózis.")
11
Finom szálakból áll, az egész sejtet behálózó vázfehérje-szövedék.
2. Citoplazma - A sejt alapállománya. - Két részből áll: citoszol: Folyékony, vízben gazdag. A fehérjefonalak által határolt teret tölti ki. citogél: Finom szálakból áll, az egész sejtet behálózó vázfehérje-szövedék. Befolyásolja a sejtek alakját, elősegíti a sejtalkotók mozgását, alkalmazkodik a sejt életműködéséhez.

12
A citoplazma összetétele
Víz 80,5-95,5% Fehérje 10-15% Lipid 2-4% Poliszacharid 0,1-1,5% DNS 0,4% RNS 0,82% Kis szerves molekulák 0,4% Szervetlen molekulák és ionok 1,5%

13
3. Sejtváz (citoszkeleton)
Az eukarióta sejtekben vékony fonálszerű struktúrából álló belső támaszték van, amelyet sejtváznak vagy citoszkeletonnak nevezünk. Ez a váz biztosítja: A sejt mechanikai ellenálló-képességét Stabilizálja a sejt alakját Módot ad arra, hogy a sejt változtassa alakját, mozogjon, összehúzódj A sejten belül aktív mozgások jöjjenek létre

14
A sejtváz (cytoskeleton)
A sejtváz (mikrotubulusok és filamentumok) adják meg a sejt alakját, segítik mozgását, szerepet játszanak a szállítási folyamatokban és a jelátadási folyamatokban. A sejtosztódás során a húzófonalak osztályozzák szét a két leánysejt kromoszómáit. tubulin actin actin
 adják meg a sejt alakját, segítik mozgását, szerepet játszanak a szállítási folyamatokban és a jelátadási folyamatokban. A sejtosztódás során a húzófonalak osztályozzák szét a két leánysejt kromoszómáit. tubulin. actin. actin.")
15
Fehérjetermészetű fonalak Mikrotubulusok Mikrofilamentumok Intermedier filamentumok
Csőszerű struktúra 25 nm vastag Mikrofilamentumok Vékony fonalak 5-7 nm vastagságú Intermedier filamentumok Fonálszerű struktúrák Vastagsága 10 nm

17
A sejtmag Az állati sejtek magja a citoplazmától elkülönült, maghártyával határolt organellum. Itt található az örökítő anyag (túlnyomó többsége). A sejtmag DNS-e az osztódási folyamat során kromoszómákba szerveződik. A kromoszómákban tárolt információ átadására (DNS-ből RNS-be) is a magban kerül sor A magon belül magvacskák figyelhetők meg, amelyek más organellumok (pl. riboszómák) szintéziséért és szervezéséért felelősek A magmembrán pórusain a makromolekulák szabályozott transzportja folyik. A magmembrán a citoplazmás membránokkal (endoplazmás retikulum) összefüggő rendszert alkot.
 is a magban kerül sor. A magon belül magvacskák figyelhetők meg, amelyek más organellumok (pl. riboszómák) szintéziséért és szervezéséért felelősek. A magmembrán pórusain a makromolekulák szabályozott transzportja folyik. A magmembrán a citoplazmás membránokkal (endoplazmás retikulum) összefüggő rendszert alkot.")
18
Sejtmag A sejtmag átmérője 10 és 20 mikrométer között váltakozhat. Dupla rétegű membrán veszi körül, melynek két rétege egymással néhol összeér, pórusokat hozva létre. Ezeken át zajlik a magi transzport, melynek fontos szerepe van a sejt működésében, hiszen az mRNS-ek a magban keletkeznek, de a citoplazmában szerelődnek össze, másrészt pedig a legtöbb kémiai reakció is a magon kívül zajlik. A külső membrán folyamatosan megy át a durva felszínű endoplazmatikus retikulumba (DER), és riboszómák kapcsolódnak hozzá. A két membrán közötti teret perinukleáris térnek nevezzük, mely pedig a DER terével folytonos. A magmembrán belülre néző oldalán számos filamentum látható, ezek hozzák létre a nukleáris laminát. A sejtmagban elszórva találunk egy vagy néhány nukleoluszt (magvacskát), melyeket a nukleoplazma nevű mátrix vesz körül. A nukleoplazma folyékony, gélszerű állaggal rendelkezik (ilyen szempontból hasonlít a citoplazmára, melyben számos vegyület van oldott állapotban. Ezen anyagok közé soroljuk a nukleotid trifoszfátokat (pl. ATP), enzimeket, fehérjéket, transzkripciós faktorokat. Ezeken kívül találunk még egy rosthálózatot is, melyet nukleáris mátrixnak nevezünk.
, és riboszómák kapcsolódnak hozzá. A két membrán közötti teret perinukleáris térnek nevezzük, mely pedig a DER terével folytonos. A magmembrán belülre néző oldalán számos filamentum látható, ezek hozzák létre a nukleáris laminát. A sejtmagban elszórva találunk egy vagy néhány nukleoluszt (magvacskát), melyeket a nukleoplazma nevű mátrix vesz körül. A nukleoplazma folyékony, gélszerű állaggal rendelkezik (ilyen szempontból hasonlít a citoplazmára, melyben számos vegyület van oldott állapotban. Ezen anyagok közé soroljuk a nukleotid trifoszfátokat (pl. ATP), enzimeket, fehérjéket, transzkripciós faktorokat. Ezeken kívül találunk még egy rosthálózatot is, melyet nukleáris mátrixnak nevezünk.")
19
A sejtmag Az aktív sejtekben a citoplazma sokkal nagyobb térfogatú, mint a nyugvó sejtekben. A naív limfocita (nyíl) szinte teljes térfogatát kitölti a mag, ellentétben pl. egy endo- tél sejttel.
 szinte teljes térfogatát kitölti a mag, ellentétben. pl. egy endo- tél sejttel.")
20
Sejtmagok és osztodás A sejtmag (latinul: nucleus; görögül: karyon) pontos meghatározását nehéz megadni, ha tekintetbe vesszük a pro- és az eukarióták egymástól igen eltérő homológ sejtmagstruktúráját, valamint azt, hogy örökítő anyag (dezoxiribonukleinsav, rövidítve: DNS) nemcsak a sejtmagban, hanem a mitokondriumokban és a plasztiszokban, sőt a baktériumok plazmidjaiban is előfordul. Az eukarióták maganyagát kétrétegű sejtmaghártya határolja, a prokarióta sejt kromoszómái viszont a citoplazmában szabadon, egy riboszómamentes plazmarétegbe ágyazódva helyezkednek el. A prokariota magot ezért, az eukariota nukleusztól megkülönböztetve, nukleoidnak[241] nevezzük. A növényi sejtet mikroszkóp alatt vizsgálva a sejtfal mellett a sejtmag tűnik elsősorban a szemünkbe. Alapanyag (nucleoplasma), a kromoszómák bázikus festékekkel festődő anyaga (chromatin), az ehhez kapcsolódó egy vagy több sejtmagvacska és a sejtmaghártya alkotja. Mérete különböző lehet. Alakja is meglehetősen változó. Leggyakoribb a gömb forma. Minél aktívabb egy sejtmag, annál nagyobb a felülete és a térfogata közötti arány. A kevés és kicsi vakuólumú sejtekben rendszerint középen helyezkedik el. A nagy vakuólumok viszont kiszorítják a perifériára a többi sejtalkotóval együtt. Helye a sejtben gyakran változhat. A legtöbb sejt egyetlen sejtmagot tartalmaz. Ismerünk azonban számos olyan növényi és gombasejtet, amelyekben a magvak száma kettő vagy több. A sejtmag határolóhártyája az ER ciszternájából keletkezett, pórusokkal áttört burok. Közös eredetük ellenére az ER és a maghártya kémiai összetétele különbözik. A maghártya külső és belső oldalát alkotó membránok a pórusoknál átfolynak egymásba, s ezért a kettős maghártyát tulajdonképpen csak egyetlen membrán alkotja. Ennek ellenére a külső és a belső réteg felépítése is jelentősen eltér egymástól. Mindkét membránréteg mintegy 7 nm vastagságú. A közöttük húzódó mag körüli (perinukleáris) tér szélessége nm. Azokon a helyeken, ahol a külső és a belső membrán átmegy egymásba, nm átmérőjű pórusok helyezkednek el. Minél aktívabb a sejtmag, annál több pórusa van. A pórusok nem csupán egyszerű lyukak, hanem sokkal inkább komplikált felépítésű struktúrák. Szélükön három darab, 8 szemcséből álló fehérjegyűrű van. Közepükön gyakran még egy központi szemcse is megfigyelhető. A sejtmag több száz féle fehérjéje a citoplazmában szintetizálódik. A citoplazmatikus fehérjeszintézisben közreműködő ribonukleinsavak (rövidítve: RNS-ek) viszont a sejtmag DNS-mintáján készülnek. Ennélfogva a sejtmag és a citoplazma között jelentős anyagtranszportnak kell lebonyolódnia. Mindkét irányú szállítás a pórusokon keresztül zajlik. A sejtmaghártya belső oldalát egy nm vastagságú, a pórusoknál megszakadó réteg (ún. lamina) borítja. A lamina a mitózis kezdetekor a citoplazmába kerülve feloldódik, és csak a kései telofázisban alakul ki ismét. Szabaddá válása eredményezi a sejtmaghártya feloldódását. Fontos szerepet tölt be a belső membrán stabilizálásában és a sejtmagban lejátszódó folyamatok szabályozásában. A kromoszómák anyaga hozzákapcsolódik a laminához úgy, hogy rajta a homológ kromoszómák egymás mellett helyezkednek el. A nukleoplazmában, a citoszolhoz hasonlóan, egy fehérjeszálakból álló váz mutatható ki. Ez szoros kapcsolatban áll a maghártyához belülről simuló laminával. Valószínűleg a fehérjeváz kereszteződési pontjaihoz kötődnek a DNS feltekeredésekor kialakuló rozetták.
 pontos meghatározását nehéz megadni, ha tekintetbe vesszük a pro- és az eukarióták egymástól igen eltérő homológ sejtmagstruktúráját, valamint azt, hogy örökítő anyag (dezoxiribonukleinsav, rövidítve: DNS) nemcsak a sejtmagban, hanem a mitokondriumokban és a plasztiszokban, sőt a baktériumok plazmidjaiban is előfordul. Az eukarióták maganyagát kétrétegű sejtmaghártya határolja, a prokarióta sejt kromoszómái viszont a citoplazmában szabadon, egy riboszómamentes plazmarétegbe ágyazódva helyezkednek el. A prokariota magot ezért, az eukariota nukleusztól megkülönböztetve, nukleoidnak[241] nevezzük. A növényi sejtet mikroszkóp alatt vizsgálva a sejtfal mellett a sejtmag tűnik elsősorban a szemünkbe. Alapanyag (nucleoplasma), a kromoszómák bázikus festékekkel festődő anyaga (chromatin), az ehhez kapcsolódó egy vagy több sejtmagvacska és a sejtmaghártya alkotja. Mérete különböző lehet. Alakja is meglehetősen változó. Leggyakoribb a gömb forma. Minél aktívabb egy sejtmag, annál nagyobb a felülete és a térfogata közötti arány. A kevés és kicsi vakuólumú sejtekben rendszerint középen helyezkedik el. A nagy vakuólumok viszont kiszorítják a perifériára a többi sejtalkotóval együtt. Helye a sejtben gyakran változhat. A legtöbb sejt egyetlen sejtmagot tartalmaz. Ismerünk azonban számos olyan növényi és gombasejtet, amelyekben a magvak száma kettő vagy több. A sejtmag határolóhártyája az ER ciszternájából keletkezett, pórusokkal áttört burok. Közös eredetük ellenére az ER és a maghártya kémiai összetétele különbözik. A maghártya külső és belső oldalát alkotó membránok a pórusoknál átfolynak egymásba, s ezért a kettős maghártyát tulajdonképpen csak egyetlen membrán alkotja. Ennek ellenére a külső és a belső réteg felépítése is jelentősen eltér egymástól. Mindkét membránréteg mintegy 7 nm vastagságú. A közöttük húzódó mag körüli (perinukleáris) tér szélessége nm. Azokon a helyeken, ahol a külső és a belső membrán átmegy egymásba, nm átmérőjű pórusok helyezkednek el. Minél aktívabb a sejtmag, annál több pórusa van. A pórusok nem csupán egyszerű lyukak, hanem sokkal inkább komplikált felépítésű struktúrák. Szélükön három darab, 8 szemcséből álló fehérjegyűrű van. Közepükön gyakran még egy központi szemcse is megfigyelhető. A sejtmag több száz féle fehérjéje a citoplazmában szintetizálódik. A citoplazmatikus fehérjeszintézisben közreműködő ribonukleinsavak (rövidítve: RNS-ek) viszont a sejtmag DNS-mintáján készülnek. Ennélfogva a sejtmag és a citoplazma között jelentős anyagtranszportnak kell lebonyolódnia. Mindkét irányú szállítás a pórusokon keresztül zajlik. A sejtmaghártya belső oldalát egy nm vastagságú, a pórusoknál megszakadó réteg (ún. lamina) borítja. A lamina a mitózis kezdetekor a citoplazmába kerülve feloldódik, és csak a kései telofázisban alakul ki ismét. Szabaddá válása eredményezi a sejtmaghártya feloldódását. Fontos szerepet tölt be a belső membrán stabilizálásában és a sejtmagban lejátszódó folyamatok szabályozásában. A kromoszómák anyaga hozzákapcsolódik a laminához úgy, hogy rajta a homológ kromoszómák egymás mellett helyezkednek el. A nukleoplazmában, a citoszolhoz hasonlóan, egy fehérjeszálakból álló váz mutatható ki. Ez szoros kapcsolatban áll a maghártyához belülről simuló laminával. Valószínűleg a fehérjeváz kereszteződési pontjaihoz kötődnek a DNS feltekeredésekor kialakuló rozetták.")
21
A kromatinállomány Az eukarióta szervezetek DNS-éhez fehérjék kötődnek. Ezek közreműködésével tekeredik föl a DNS nagyobb egységekbe. A föltekeredett és ily módon összetömörült DNS-t már egy évszázaddal ezelőtt sikerült bázikus festékekkel kimutatni. A jól festődő anyagot kromatinállománynak nevezték el. A DNS-hez kötődő fehérjék legfontosabb csoportját a bázikus tulajdonságú hisztonok alkotják. Mellettük savas, ún. nem hisztonszerű fehérjék is kapcsolódnak a DNS-hez. A kromatinállomány további alkotói a DNS-mintán éppen átíródó RNS, valamint a folyamatot katalizáló enzimek A keleti jácint (Hyacinthus orientalis, Hyacinthaceae) gyökércsúcssejtjének sejtmagja sejtmagvacskával és kromatinállománnyal (Fridvalszky Lóránt felvétele) Az említett molekuláris alkotók három lépésben építik fel a kromatinállományt. Az első lépésben egy laza, gyöngyfüzérszerű állapot jön létre. Ennek gyöngyszemeit, az ún. nukleoszómákat, négyféle (H2A, H2B, H3 és H4 jelű) hisztonfehérje alkotja. A nukleoszómák létrehozásához a H3 és H4 jelű hisztonokra feltétlenül szükség van, míg a folyamat a H2A és a H2B jelű hisztonok nélkül végbemegy. A nukleoszómákra a 2 nm-es vastagságú DNS-szálak kétszer tekerednek fel. Egy-egy ilyen feltekeredett DNS-szakasz 166 bázispárt tartalmaz. A DNS-sel borított nukleoszómák 5,7 x 11 nm nagyságúak. A nukleoszómákat a laza gyöngyfüzéren belül bázispárt magukba foglaló DNS-szálak (ún. linkerek) kötik össze. A második lépésben a gyöngyfüzér a H1 jelű hiszton hatására összetömörül úgy, hogy a nukleoszómák a DNS-t nem tartalmazó oldalaikkal sűrűn egymás mellé rendeződnek. Ekkor 11 nm-es vastagságú fonal jön létre. Ez magnéziumionok jelenlétében spirális alakban feltekeredik, és egy nm-es vastagságú, fordulatonként 6-7 nukleoszómát tartalmazó fonalat képez. A H1-es hiszton elvonásakor a fonál ismét kinyúlt nukleoszómákra bomlik le. A harmadik lépésben a nm-es vastagságú fonalak hurkokból álló rozettaszerű képződményekké hajtogatódnak össze. Egy-egy ilyen rozetta bázispárt tartalmaz. Ekkorra már a DNS-molekula az eredeti hosszának mintegy ötszázad részére zsugorodik össze. A kromatinállomány jelentős része ilyen formában van jelen az interfázisban lévő sejtmagban. Információ leadásakor a rozetta sok kicsi hurokja egyetlen nagy hurokká bomlik le. Sejtosztódáskor viszont több rozetta egyesülésével kis csomók (chromomer) keletkeznek.
 gyökércsúcssejtjének sejtmagja sejtmagvacskával és kromatinállománnyal (Fridvalszky Lóránt felvétele) Az említett molekuláris alkotók három lépésben építik fel a kromatinállományt. Az első lépésben egy laza, gyöngyfüzérszerű állapot jön létre. Ennek gyöngyszemeit, az ún. nukleoszómákat, négyféle (H2A, H2B, H3 és H4 jelű) hisztonfehérje alkotja. A nukleoszómák létrehozásához a H3 és H4 jelű hisztonokra feltétlenül szükség van, míg a folyamat a H2A és a H2B jelű hisztonok nélkül végbemegy. A nukleoszómákra a 2 nm-es vastagságú DNS-szálak kétszer tekerednek fel. Egy-egy ilyen feltekeredett DNS-szakasz 166 bázispárt tartalmaz. A DNS-sel borított nukleoszómák 5,7 x 11 nm nagyságúak. A nukleoszómákat a laza gyöngyfüzéren belül bázispárt magukba foglaló DNS-szálak (ún. linkerek) kötik össze. A második lépésben a gyöngyfüzér a H1 jelű hiszton hatására összetömörül úgy, hogy a nukleoszómák a DNS-t nem tartalmazó oldalaikkal sűrűn egymás mellé rendeződnek. Ekkor 11 nm-es vastagságú fonal jön létre. Ez magnéziumionok jelenlétében spirális alakban feltekeredik, és egy nm-es vastagságú, fordulatonként 6-7 nukleoszómát tartalmazó fonalat képez. A H1-es hiszton elvonásakor a fonál ismét kinyúlt nukleoszómákra bomlik le. A harmadik lépésben a nm-es vastagságú fonalak hurkokból álló rozettaszerű képződményekké hajtogatódnak össze. Egy-egy ilyen rozetta bázispárt tartalmaz. Ekkorra már a DNS-molekula az eredeti hosszának mintegy ötszázad részére zsugorodik össze. A kromatinállomány jelentős része ilyen formában van jelen az interfázisban lévő sejtmagban. Információ leadásakor a rozetta sok kicsi hurokja egyetlen nagy hurokká bomlik le. Sejtosztódáskor viszont több rozetta egyesülésével kis csomók (chromomer) keletkeznek.")
22
A citoplazma organellumai és az endomembránrendszer

23
Az endoplazmás retikulum
A maghártyával összefüggő ER reakció-edények és csövek bonyolult hálózata. Itt termelődnek a fehérjék (az ER-hez kap-csolódó riboszómákon) és a lipidek. Az ER belső felszíne és lumene enzimek-ben gazdag (ld. fehérjék érési folyamatát). Az ER a sejt többi kompartmentjével inten-zív kapcsolatban van: „csövei”, vezikulumai fehérjéket és lipideket juttatnak célba más organellumokhoz (ld. vezikuláris transzport, és sorting). A riboszómákat tartalmazó ER-t „durva” (rough), a riboszóma-mentes ER-t síma (smooth) jelzővel különböztetjük meg (RER és SER).
 és a lipidek. Az ER belső felszíne és lumene enzimek-ben gazdag (ld. fehérjék érési folyamatát). Az ER a sejt többi kompartmentjével inten-zív kapcsolatban van: „csövei , vezikulumai fehérjéket és lipideket juttatnak célba más organellumokhoz (ld. vezikuláris transzport, és sorting). A riboszómákat tartalmazó ER-t „durva (rough), a riboszóma-mentes ER-t síma (smooth) jelzővel különböztetjük meg (RER és SER).")
24
A „durva” ER (RER) A RER-t a fenti képen sárgán fluoresz-káló festékkel tették láthatóvá, jól látszik a maggal való szoros kapcsoltsága. Az elektronmikroszkópos (EM) képen a riboszómák apró, sötét foltokként jelennek meg az ER membránjainak felszínéhez kapcsolódva. Az ER belseje – a magas fehérje tar-talom miatt – sötétebbnek (electron dense) látszik, mint a környező cito-plazma.
 képen a riboszómák apró, sötét foltokként jelennek meg az ER membránjainak felszínéhez kapcsolódva. Az ER belseje – a magas fehérje tar-talom miatt – sötétebbnek (electron dense) látszik, mint a környező cito-plazma.")
25
A riboszómák A riboszómák a sejt fehérjeszintézisének központjai. Nevük nagy RNS-tartalmukra utal. A citoplazmában, a kloroplasztiszokban és a mitokondriumokban, valamint az ER felszínén feltűnően nagy tömegben előforduló sejtalkotók. Kisebb számban a Golgi-apparátus, a sejtmag, a mitokondriumok és a kloroplasztiszok határoló hártyájának citoplazma felé eső felszínén is megtalálhatók. A durva felszínű ER nevét éppen a riboszómák jelenléte miatt kapta. Citoplazmatikus oldalán a riboszómák gyakran hurokszerű képleteket, ún. poliriboszómákat alkotnak. A citoplazma szabad riboszómái közül sok szintén rozetta vagy csavarvonal alakú poliriboszómákat képez. A riboszómák a képen látható kettéhasadó mitokondrium körül apró gömböcskékként ismerhetők fel (Fridvalszky Lóránt felvétele) Két növényi sejt elektronmikroszkópos részlete (Fridvalszky Lóránt felvétele) A baktérimokban egyféle, az állatokban, a gombákban és a nem fotoszintetizáló növényekben kétféle - citoplazmatikus és mitokondriális -, a zöld növényekben pedig háromféle - citoplazmatikus, mitokondriális és kloroplasztisz- - riboszóma fordul elő. A prokarióta, a mitokondriális és a kloroplasztisz-riboszóma nm, míg az eukarióták citoplazmatikus riboszómája 21 nm átmérőjű. Mindegyik egy kisebb és egy nagyobb alegységből áll, amelyek a sejtmagvacskában külön-külön képződnek, majd a sejtmaghártya pórusain kijutva a citoplazmába az mRNS felszínén egyesülnek riboszómává.
 Két növényi sejt elektronmikroszkópos részlete (Fridvalszky Lóránt felvétele) A baktérimokban egyféle, az állatokban, a gombákban és a nem fotoszintetizáló növényekben kétféle - citoplazmatikus és mitokondriális -, a zöld növényekben pedig háromféle - citoplazmatikus, mitokondriális és kloroplasztisz- - riboszóma fordul elő. A prokarióta, a mitokondriális és a kloroplasztisz-riboszóma nm, míg az eukarióták citoplazmatikus riboszómája 21 nm átmérőjű. Mindegyik egy kisebb és egy nagyobb alegységből áll, amelyek a sejtmagvacskában külön-külön képződnek, majd a sejtmaghártya pórusain kijutva a citoplazmába az mRNS felszínén egyesülnek riboszómává.")
26
A riboszómák A riboszómák fehérjék és RNS molekulák komplexei. A ribo-
szómák végzik a fehérjék szintézisét. Az oldható fehérjéket a citoplazma riboszómái készítik, a membrán fehérjéket és az „exportra” kerülő fehérjéket pedig a durva endoplazmás retiku-lum (RER) riboszómái.
 riboszómái.")
27
Membrán-kötött riboszómák és az ER
A mag felett riboszómákban gazdag RER helyezkedik el, két oldalon viszont jól láthatók a síma ER riboszóma-mentes csövei, hólyagjai, vezikulumai.

28
A Golgi készülék A Golgi apparátus az ER-hoz hasonló felépítésű, ahhoz kapcsolódó membrán-rendszer, amelynek feladata a fehérjék és lipidek módosítása, osztályozása és célba juttatása. A Golgi készülékről lefűződő vezikulu-mok más sejtszervecskékbe juttatják az oda „címzett” anyagokat, de a Golgi „fogadja” is az onnan érkező küldeményeket, további feldolgozásra. A Golgi fontos feladata a „selejtes” termékek eljuttatása a lebontó lizoszómákba.

29
A képen a Golgi zölden fluoreszkál

30
Golgi apparátus

31
Az ER és a Golgi készülék kapcsolata
Az ER-ból a Golgi készülékbe jutnak a frissen szintetizált anyagok mellett az ER fontos alkotóelemei (pl. enzimek, lipidek) is. Ezek a Golgiról lefűződő vezikulumokkal jutnak vissza az ER-ba
 is. Ezek a Golgiról lefűződő vezikulumokkal jutnak vissza az ER-ba.")
32
Proteaszómák és lizoszómák
A poteaszómák enzimkomplexek, amelyek feladata a fehérjék lebontása. A lizoszómák membránhatárolt vezikulumok, amelyek bontóenzimeket tartal-maznak. A lizoszómák savas pH-n működnek,fehérjék mellett nukleinsavak, lipidek és szénhidrátok lebontására is képesek. A fehérjéket több okból bonthatja le a sejt: azért, mert rossz szerkezettel rendelkeznek (selejtes vagy sérült) más fehérjék eleve rövid életidejűek enzimek, szabályzó fehérjék, receptorok szükségtelenné válnak a sejt aminosavakra éhezik Vannak specifikus „jelek”, amelyek a lebontandó fehérjéket megjelölik. lysosome
 más fehérjék eleve rövid életidejűek. enzimek, szabályzó fehérjék, receptorok szükségtelenné válnak. a sejt aminosavakra éhezik. Vannak specifikus „jelek , amelyek a lebontandó fehérjéket megjelölik. lysosome.")
33
Peroxiszómák A peroxiszómák enzimekkel teli, kicsi vezikulumok, amelyek feladata a veszélyes peroxidok és szabadgyökök lebontása, hatástalanítása. A peroxiszómák enzimei gyakran kikristályodnak. A peroxiszómák száma sejttípustól és anyagcsere-tevékenységtől függően igen változó lehet.

34
A sejtnedvüreg (vacuolum)
A Golgi-apparátus fontos feladata az állati sejtekben az emésztésben közreműködő elsődleges lizoszómák képzése és azok bontó, hidrolitikus enzimekkel való ellátása. A lizoszómák a sima felszínű ER és a Golgi-apparátus közötti átmeneti elemekből fűződnek le. Mivel azonban a növények savas hidrolázai mindenekelőtt a vakuólumokban fordulnak elő, a növényi sejtek ezen alkotóit tekintjük az állati lizoszómákkal egyenértékű sejtalkotóknak.

35
Mitochondrium

36
Mitokondriumok A képen a mitokondriumok zölden fluoreszkálnak

37
A mikrotubulusok A mikrotubulusok leggyakrabban a citoplazma sejtfallal érintkező részén megjelenő csőszerű képződmények . A sejtfal képzésében működhetnek közre, ugyanis annak cellulóz-mikrofibrillumai mindig a plazmalemma mentén elrendeződő mikrotubulusok irányába rakódnak le. A sejt- osztódáskor kialakuló magorsót is mikrotubulusok alkotják. A fragmoplaszt, az alapi testek, az ostorok és a csillók létrehozásában is szerepük van. Átmérőjük 24 nm. Tubulin nevű fehérjealegységeikből a sejt meghatározott pontján keletkeznek. Az őszi kikerics kolchicin nevű és a kis télizöld vinblasztin, valamint vinkrisztin nevű alkaloidjai meggátolják a mikrotubulusok tubulin-alegységekből való összeállítódását. Az ilyen anyagokkal kezelt sejtek magjai a magorsó kialakulásának akadályoztatása miatt nem osztódnak, a cellulóz-mikrofibrillumok[267] rendezetlen lerakódása következtében pedig minden irányban azonos kiterjedésűek (izodiametrikusak) lesznek. A kromoszómák megkettőződését viszont az említett alkaloidok nem befolyásolják, és így alkalmazásukkor poliploid sejtek keletkeznek.
 lesznek. A kromoszómák megkettőződését viszont az említett alkaloidok nem befolyásolják, és így alkalmazásukkor poliploid sejtek keletkeznek.")
38
A mitokondrium A mitokondrium a sejt energia ellátását biztosítja. Egy sejten belül számos mitokondrium található. Általában, minél több energiát használ fel egy sejt, annál több mitokondriuma van. A mitokondriumot két membrán veszi körül, a belső prokariótákra jellemző tulajdonságokat mutat. A mitokondriumok öse a mai bíborbaktériumokhoz hasonló szervezet lehetett.

39
Centroszómák és centriolumok
A centroszómák a mag közelében, a citoplazmában találhatók.Az S fázis alatt megkettőződnek és a sejt távoli pontjaira vándorolnak. A mitózis alatt mikrotubulusok nőnek ki belőlük, amelyek húzófonalakat (spindle fibers) alkotnak és a kromoszómák szétválásá- ért felelősek. A centroszómában két iker centriólum van, ezek az állati sejtekre jellemző organellumok.
 alkotnak és a kromoszómák szétválásá- ért felelősek. A centroszómában két iker centriólum. van, ezek az állati sejtekre jellemző. organellumok.")
40
Csilló (cilium) és Ostor (flagellum)
Centriolum A sejt közepén, a sejtmag mellett elhelyezkedő organellum, a citocentrum (sejtközpont) központjában elhelyezkedő hengerded testecske, amely körben rendeződő mikrotubulustripletből áll. Két centriolum találhat a citocentrum közepén egymás mellett (diploszóma). Sejtbiológiai szerepe nem tisztázott. Csilló (cilium) és Ostor (flagellum) Egyes sejttípusok felszínéről kinyúló, hengeres nyúlványok. Jellegzetes csapkodó, kígyózó mozgás. Körben kilenc mikrotubuluspár, középen két mikrotubulus helyezkedik el. A mikrotubuluspárok közötti résben dineinkomplexek találhatóak.. A dineinkomplexek talpa a mikrotubulushoz tapad, mozgékony feji része a szomszédos mikrotubulust mozdítja el hosszanti irányba. A szomszédos mikrotubulusok elcsúsznak egymás mellett, ami a csilló meggörbüléséhez vezet. Ehhez ATP-hasításból származó energia szükséges.
 központjában elhelyezkedő hengerded testecske, amely körben rendeződő mikrotubulustripletből áll. Két centriolum találhat a citocentrum közepén egymás mellett (diploszóma). Sejtbiológiai szerepe nem tisztázott. Csilló (cilium) és Ostor (flagellum) Egyes sejttípusok felszínéről kinyúló, hengeres nyúlványok. Jellegzetes csapkodó, kígyózó mozgás. Körben kilenc mikrotubuluspár, középen két mikrotubulus helyezkedik el. A mikrotubuluspárok közötti résben dineinkomplexek találhatóak.. A dineinkomplexek talpa a mikrotubulushoz tapad, mozgékony feji része a szomszédos mikrotubulust mozdítja el hosszanti irányba. A szomszédos mikrotubulusok elcsúsznak egymás mellett, ami a csilló meggörbüléséhez vezet. Ehhez ATP-hasításból származó energia szükséges.")
41
Csilló (cilium) és Ostor (flagellum)
Kívülről sejtmembrán borítja, tövénél bazális test található. Mozgásuk hasznosul. Szabad sejt esetében az előremozgást szolgálja (pl. csillós és ostoros egysejtűek, spermiumok) Hámrétegbe épített csillós sejt a hámréteg felszínét borító folyadék áramlását biztosítja (légcső és a petevezeték hámja).
 Hámrétegbe épített csillós sejt a hámréteg felszínét borító folyadék áramlását biztosítja (légcső és a petevezeték hámja).")
42
Őszi kikerics (Colchicum autumnale, Colchicaceae)
")

















>")
>")

