Előadást letölteni
1
AZ INAK ÉS SZALAGOK BIOMECHANIKÁJA

4
Az inak és szalagok összetétele és szerkezete
Sejtes anyag 20 %, Sejtközötti állomány 80% A sejtközötti állomány 70 % vizet, 30 % szilárd anyagot tartalmaz

5
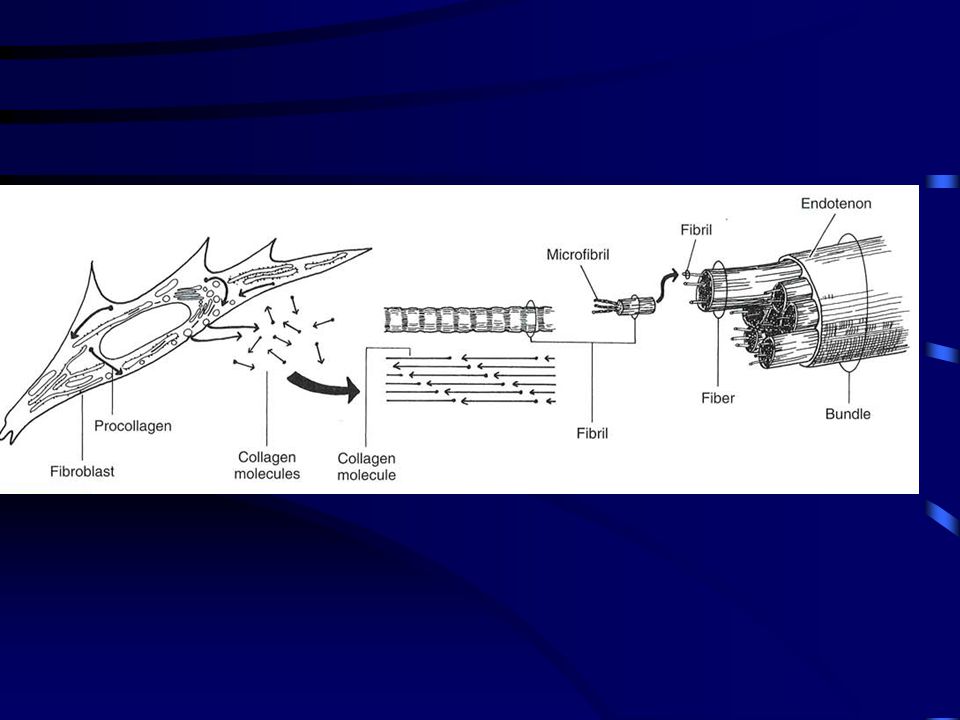
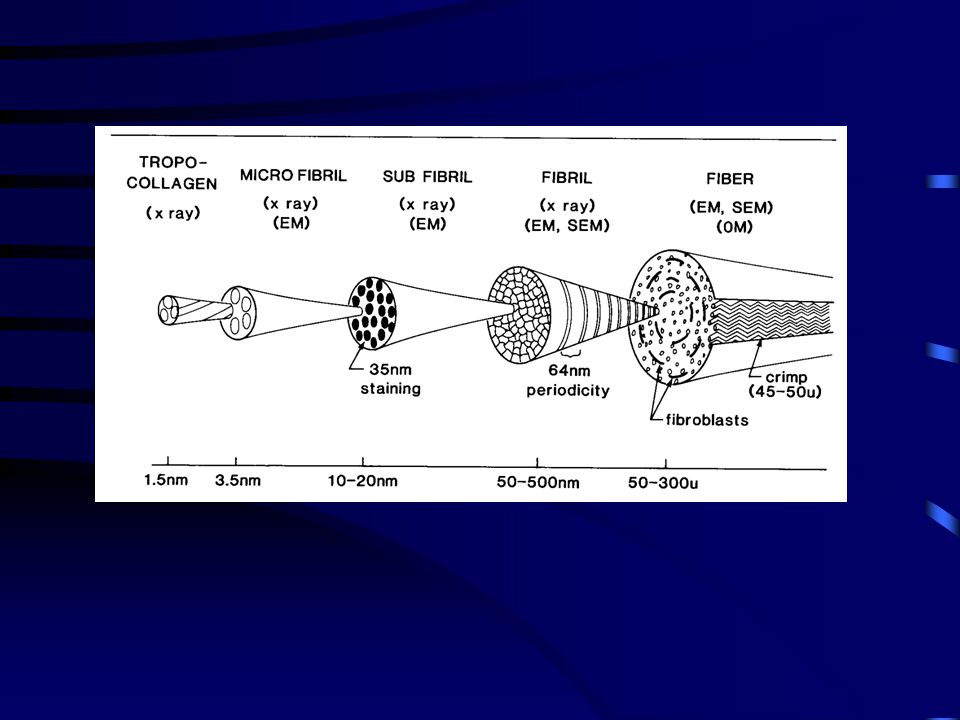
A kollagének mikrostruktúrája
Keresztösszeköttetés is található a kollagén molekulák között, amelyek lényeges szereppel bírnak a molekulák fibrulomokká alakításában. A keresztösszekötetés növeli a kollagén fibrillumok erőkifejtését a nyújtó erővel szemben. A kollagén molekulák lépcsőzetesen eltolt kötegekké szerveződnek. Az inak és szalagok I típusú kollagénekből állnak. Ez a molekula három polipeptide láncból ( lánc) formálódik, mindegyik helixé tekeredve.
 formálódik, mindegyik helixé tekeredve.")
6
INAK SZALAGOK

7
ELASZTIKUS ROSTOK ÉS ELASZTIN
Az egészséges emberi inak 10 %-ban elasztikus rostokból épülnek fel. 2%-az inak szárazanyag tartalmának nem kollagén fehérje, hanem elastin. A rostos porc és az ásványi anyag tartalmú rostos porc csont-ín összeköttetésnél elasztikus rostokból állnak.

8
LIGAMENTUM FLAVUM ELASZTIN : KOLLAGÉN = 2 : 1

9
ÍN-IZOM, ÍN-CSONT ÁTMENET
Ín-izom átmenet midsubstance IZOM ÍN CSONT

10
Ujjszerű befűződések:
1-8 m ín Az átkapcsolódási régió %-al nagyobb a II típusú izomrostok esetében izom

11
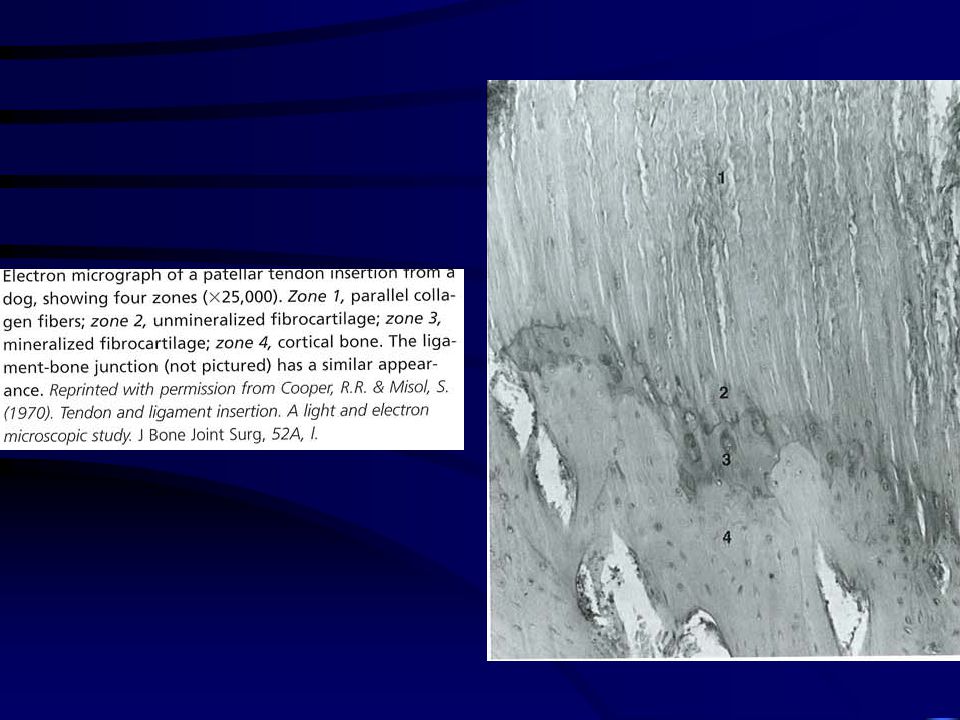
Rostos porc (gyerekek 1-2 mm, felnőttek 150-400 m)
Ín-csont átmenet Rostos porc (gyerekek 1-2 mm, felnőttek m) Rostos porc
 Rostos porc.")
13
Vérellátás Az Achilles inban 2-6 cm-es zónában nem találhatók kapillárisok az ín-izom átmenetnél.

14
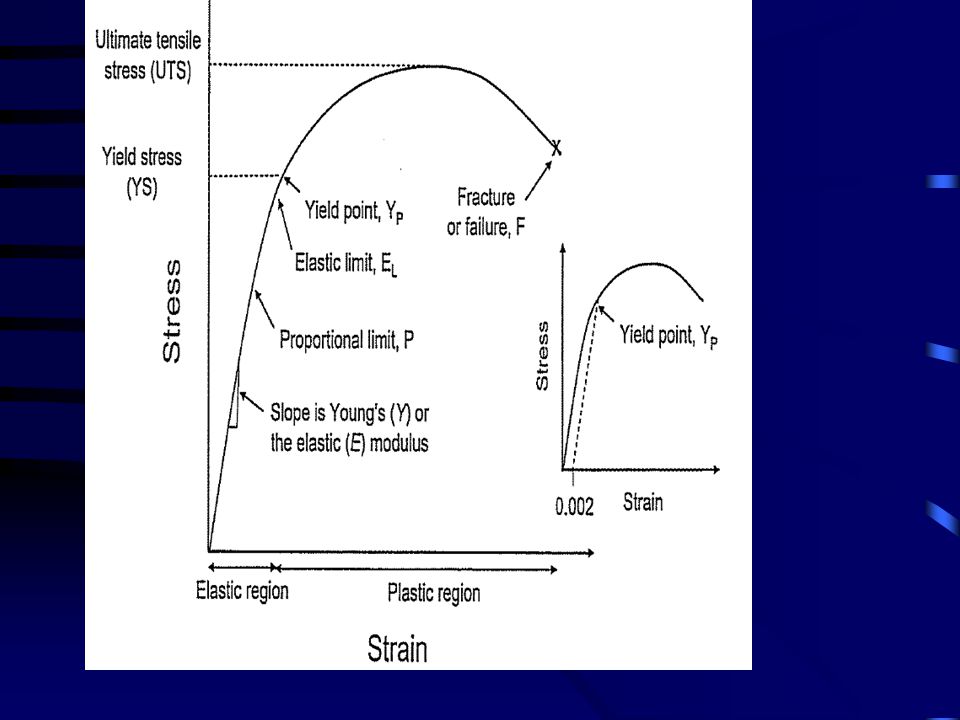
BIOMECHANIKAI JELLEMZŐK

15
ELASTIKUS/ YOUNG MODULUS
NYÚJTÁSI ERŐ NYÚJTÁSI ERŐ MEGNYÚLÁS MEGNYÚLÁS STRESS - STRAIN STIFFNESS - COMPLIENCE ELASTIKUS/ YOUNG MODULUS NYÚJTÁSI ENERGIA HISZTERÉZIS

16
Erő-elmozdulás összefüggés
ACL Length: area:

17
ERŐ – MEGNYÚLÁS GÖRBE dL = 15 mm F = 1500 N Noyes and Grood, 1976
Noyes et al. 1984 dL = 15 mm F = 1500 N

19
Relaxált Megnyújtott

20
A patella ín hossz-feszülés jellemzőinek mérése kadaver modellen
PT ACL Noyes et al. 1984 Ahmed et al. 1987

21
Kadaver inak megnyúlása
Noyes et al. 1984 dL = 10 mm F = 3000 N Ahmed et al. 1987 Huberti et al. 1984 F = N

22
ELASTIKUS/ YOUNG MODULUS
NYÚJTÁSI ERŐ MEGNYÚLÁS STIFFNESS - COMPLIENCE STRESS - STRAIN ELASTIKUS/ YOUNG MODULUS NYÚJTÁSI ENERGIA HISZTERÉZIS

23
ERŐ – MEGNYÚLÁS KAPCSOLAT
Stiffness = dF • dl-1 769.2 N m-1 335 N m-1 dF dl Noyes et al. 1984

24
Az ín hosszúság és keresztmetszet hatása a stiffness-re
STIFFNESS = dF / dL COMPLIENCE = dL / dF

25
STIFFNESS 129 N/mm 182 N/mm 194 N/mm 140.8 N/mm 161 N/mm 756 N/mm
ACL Ember (50 yr) 129 N/mm Noyes et al.1976 Ember (22 yr) 182 N/mm Rheusus majom 194 N/mm In vivo Első interosseus izom ina a mutatóujjon 140.8 N/mm Cook and McDnogh, 1996 Tibialis anterior ín 161 N/mm Maganaris and Paul, 1999 Patella ín 756 N/mm Tihanyi et al., 2000
 129 N/mm. Noyes et al Ember (22 yr) 182 N/mm. Rheusus majom. 194 N/mm. In vivo. Első interosseus izom ina a mutatóujjon N/mm. Cook and McDnogh, Tibialis anterior ín. 161 N/mm. Maganaris and Paul, Patella ín. 756 N/mm. Tihanyi et al.,")
26
ELASTIKUS/ YOUNG MODULUS
NYÚJTÁSI ERŐ MEGNYÚLÁS STIFFNESS - COMPLIENCE STRESS - STRAIN ELASTIKUS/ YOUNG MODULUS NYÚJTÁSI ENERGIA HISZTERÉZIS

27
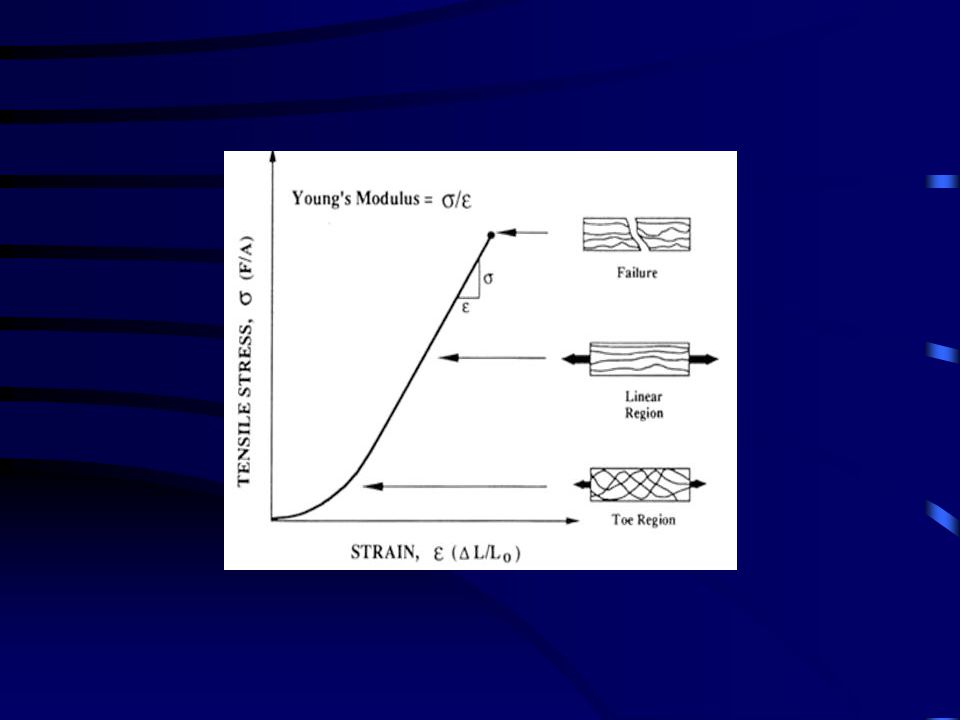
N / m2, Pa STRESS - STRAIN Hogyan számítjuk ?
Erő / keresztmetszeti terület N / m2, Pa

28
digital extensor és flexor ín
ÁLLAT MODELL Shadwick (1990) Újszülött Felnőtt Sertés digital extensor és flexor ín Megnyúlás: Stress : 17%, 7-9 % 16 MPa 40-90 MPa
 Újszülött. Felnőtt. Sertés. digital extensor és flexor ín. Megnyúlás: Stress : 17%, 7-9 % 16 MPa MPa.")
29
HUMAN MODELL ACL Idős Fiatal Rheosus majom Strain (%): Stress (MPa): 21.9 25.5 38.0 13.3 37.8 66.1 In vivo Tibialis anterior Patella ín Strain (%): Stress (MPa): 2.5 17.9 25 39.3
: Stress (MPa):")
30
ELASTIKUS/ YOUNG MODULUS
NYÚJTÁSI ERŐ MEGNYÚLÁS STIFFNESS - COMPLIENCE STRESS - STRAIN ELASTIKUS/ YOUNG MODULUS NYÚJTÁSI ENERGIA HISZTERÉZIS

31
ELASTIKUS/ YOUNG MODULUS
E = (F/A) / dl/L
 / dl/L.")
32
ELASTIKUS (YOUNG) MODULUS
Δσ Δε

33
HUMAN MODELL 65.3 111 186 1200 260 Számítások: 1200 - 2900 MPa
ACL Idős Fiatal Rheosus majom E (MPa): 65.3 111 186 In vivo Tibialis anterior Patella ín E (MPa): 1200 260 Számítások: MPa
: In vivo. Tibialis anterior. Patella ín. E (MPa): Számítások: MPa.")
34
RUGÓ TÍPUSÚ AZ ÍN ? AQF APT > 30

35
Nyújtási energia W = dF · dl

36
2.0 – 5.0 AZ ÍNAK BIZTONSÁGI FAKTORA Maximális feszülés (erő)
A fizikai aktivatás alatt meghatározott maximális erő 2.0 – 5.0

37
Mélybeugrás Leugrási magasság: 40 cm Erőplató: Kistler FP 9287A

38
Forgatónyomaték a térdnél
Patella ínra ható erő Biztonsági faktor 3.0 L = m

39
Biztonsági faktor 1.4

40
M = 580 Nm F = N ? Biztonsági faktor

41
HISZTERÉZIS

42
HISZTERÉZIS Hiszterézis = A/ A+B · 100 5.1 %

43
A keresztösszeköttetések száma 20 éves korig nővekszik, majd csökken.
MATURÁCIÓ ÉS ÉLETKOR Az inak mechanikai tulajdonságai a keresztösszeköttetések számától függ. A keresztösszeköttetések száma 20 éves korig nővekszik, majd csökken.

44
digital extensor és flexor ín
Nyújtási energia (J/kg) digital extensor és flexor ín Újszülött Felnött E: 900 Human patella ín E: 5744
 digital extensor és flexor ín. Újszülött. Felnött. E: Human patella ín. E:")
45
A FIZIKAI TERHELÉS HATÁSA
Növekszik a maximális nyújtóerő elasztikus energiatárolás a sérülésekkel szembeni ellenállóképesség

46
A bemelegítés hatása Növekszik A stiffness nem változik
a nyújtási erő, A megnyúlás mértéke elasztikus energia tároló képesség A stiffness nem változik

47
IMMOBILIZÁCIÓ - REHABILITACIÓ

48
energy % Force 100 100 91 92 79 78 69 61 intact intact IMMOB (8 hét)
95 energy 90 Force 85 80 75 100 100 91 92 70 65 79 78 60 69 61 55 50 intact intact IMMOB (8 hét) IMMOB (8 hét) REHAB (5 hónap) REHAB (12 hónap) REHAB (5 hónap) REHAB (12 hónap)
 IMMOB (8 hét) REHAB (5 hónap) REHAB (12 hónap) REHAB (5 hónap) REHAB (12 hónap)")
49
A GYÓGYSZEREK HATÁSA

50
NEM SZTEROIDOK (aspirin, indometacin stb.)
Mechanikai hatás: Megnövekedett ellenállás a nyújtással szemben Szöveti hatás: a kollagén tartalom megnövekszik a keresztösszeköttetés száma megnövekszik

51
Corticosteroid (katabolikus hatás)
gyengíti a kötőszövetet atrofia Anabolikus steroid csökkenő ellenállás a megnyújtó erővel szemben izom-ín erő arány növekszik

53
A sérülés bekövetkezése (%) az ínhossz függvényében
 az ínhossz függvényében")
54
Molecular Basis for Elastic Energy Storage in Mineralized Tendon
Frederick H. Silver,*,† Joseph W. Freeman,† Istvan Horvath,† and William J. Landis‡ Molecular Basis for Elastic Energy Storage in Mineralized Tendon Biomacromolecules 2001, 2, pdf It is proposed that mineralization increases the stored elastic energy by preventing flexible regions within the positively stained bands from stretching. These observations suggest that mineralization begins in the hole region due to the large number of charged amino acid residues found in the d and e bands.

55
Tendon consists primarily of cells, collagen fibers,
proteoglycans, and water. Type I collagen is the most abundant protein in tendon and is a fibril-forming protein that self-assembles into cross-striated fibrils having a 67 nm repeat. The molecule is a triple helix composed of approximately 1000 amino acids and extending about 300 nm in length.7 (7) Silver, F. H. Biological Materials: Structure, Mechanical Properties and Modeling of Soft Tissues; New York University Press: New York, 1987; pp 7-21, ,
 Silver, F. H. Biological Materials: Structure, Mechanical Properties. and Modeling of Soft Tissues; New York University Press: New. York, 1987; pp 7-21, ,")
56
Collagen has been reported to be a rodlike
molecule with little flexibility and high mechanical strength;8 however, some measurements indicate that the type I collagen molecule has numerous bends and is not completely rigid.9,10 (9) Hofmann, H.; Voss, T.; Kuhn, K.; Engle, J. Localization of flexible sites in threadlike molecules from electron micrographs: Comparison of interstitial, basement membrane and intimal collagens. J. Mol. Biol. 1984, 172, (10) Silver, F. H.; Birk, D. E. Molecular structure of collagen in solution: comparison of types I, II, III, and V. Int. J. Biol. Macromol. 1984, 6,
 Hofmann, H.; Voss, T.; Kuhn, K.; Engle, J. Localization of flexible. sites in threadlike molecules from electron micrographs: Comparison. of interstitial, basement membrane and intimal collagens. J. Mol. Biol. 1984, 172, (10) Silver, F. H.; Birk, D. E. Molecular structure of collagen in. solution: comparison of types I, II, III, and V. Int. J. Biol. Macromol. 1984, 6,")
57
Analysis of the amino acid sequence and physical properties in solution indicates that some regions of the molecule are more flexible than others.6,10 (6) Silver, F. H.; Freeman, J.; Seehra, G. P. Collagen self-assembly and development of matrix mechanical properties. Submitted for publication. (10) Silver, F. H.; Birk, D. E. Molecular structure of collagen in solution: comparison of types I, II, III, and V. Int. J. Biol. Macromol. 1984, 6,
 Silver, F. H.; Freeman, J.; Seehra, G. P. Collagen self-assembly and. development of matrix mechanical properties. Submitted for publication. (10) Silver, F. H.; Birk, D. E. Molecular structure of collagen in. solution: comparison of types I, II, III, and V. Int. J. Biol. Macromol. 1984, 6,")
58
the turkey gastrocnemius tendon change during mineralization.11
(11) Landis, W. J.; LiBrizzi, J. J.; Dunn, M. G.; Silver, F. H. A study of the relationship between mineral content and mechanical properties of turkey gastrocnemius tendon. J. Bone Mineral. Res. 1995, 10, While it has been proposed that mineralization increases the ability of turkey tendon to store elastic energy,5 (5) Silver, F. H.; Christiansen, D.; Snowhill, P. B.; Chen. Y.; Landis, W. J. The role of mineral in the storage of elastic energy in turkey tendons. Biomacromolecules 2000, 1,
 Landis, W. J.; LiBrizzi, J. J.; Dunn, M. G.; Silver, F. H. A study of. the relationship between mineral content and mechanical properties. of turkey gastrocnemius tendon. J. Bone Mineral. Res. 1995, 10, While it has been proposed that mineralization increases the ability of turkey tendon to store elastic energy,5. (5) Silver, F. H.; Christiansen, D.; Snowhill, P. B.; Chen. Y.; Landis, W. J. The role of mineral in the storage of elastic energy in turkey. tendons. Biomacromolecules 2000, 1,")
59
Figure 1. Stress-strain curve of a 14 week old turkey tendon (0
Figure 1. Stress-strain curve of a 14 week old turkey tendon (0.295 mineral content). Total, elastic and viscous tensile stress-strain curves for turkey tendon obtained at a strain rate of 10%/min. The total stress-strain curve was obtained from Landis et al.11 The elastic stress-strain curve was obtained from the total stress-strain curve after multiplication by the elastic fraction reported by Silver et al.5 The viscous stress-strain curve was obtained from the difference between the total and elastic stress-strain curves.
. Total, elastic and viscous tensile stress-strain curves for turkey tendon obtained at a strain rate of 10%/min. The total stress-strain curve was obtained from Landis et al.11 The elastic stress-strain curve was obtained from the total stress-strain curve after multiplication by the elastic fraction reported by Silver et al.5 The viscous stress-strain curve was obtained from the difference between the total and elastic stress-strain curves.")
61
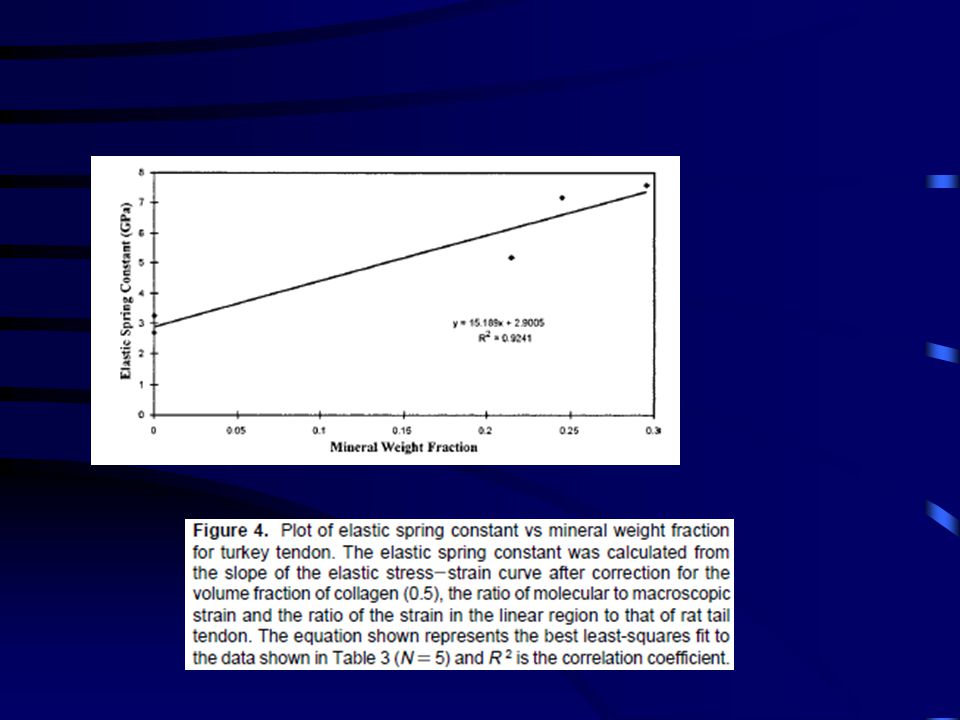
Figure 5. Plot of elastic energy stored vs mineral weight fraction for
turkey tendon. Elastic energy stored was calculated as the area under the elastic stress-strain curve (Figure 2) up to a strain of 20%. The equation shown represents the best least-squares fit to the data and R 2 is the correlation coefficient. Note there are two points at a mineral weight fraction of 0 making N ) 5.
 up to a strain of 20%. The. equation shown represents the best least-squares fit to the data and. R 2 is the correlation coefficient. Note there are two points at a mineral. weight fraction of 0 making N ) 5.")
63
Figure 6. (Top) Diagram illustrating the packing pattern of type I collagen molecules into a subfibrillar unit containing five molecules in cross section. The subunit contains collagen molecules that have a total length of about 4.4D and are staggered laterally by multiples of D, which is 67 nm. The D-period is the characteristic fingerprint of collagen that arises from the repeat of the hole and overlap regions. The hole region is characterized by the presence of only four molecules in the cross section while the overlap region contains cross sections of five molecules. The hole region is a result of the presence of a 0.6D space between neighboring molecules along the axis of the subunit. When collagen fibrils are viewed in the electron microscope after positive staining, 12 dark staining lines identified as bands c2, c1, b2, b1, a4, a3, a2, a1, e2, e1, and c3 are observed. These bands are diagrammed as flexible springs connected by rigid segments (solid areas) that represent the interband regions. The width of each region is proportional to the number of amino acids. (Bottom) Plot of band flexibility as a function of position along the positive staining pattern for a type I collagen fibril.12 Collagen fibril flexibility was estimated from the area under the conformational map for different pairs of amino acids in a subfibril containing five quarter-staggered collagen molecules. The plot of flexibility (number of conformations, a unitless variable) vs band number shown indicates that regions e2, e1, d, and c3 are continuously flexible. Other bands such as b2 and a1 are flexible but are separated by relatively rigid interband regions.
 that represent the interband. regions. The width of each region is proportional to the number of amino acids. (Bottom) Plot of band flexibility as a function of position along. the positive staining pattern for a type I collagen fibril.12 Collagen fibril flexibility was estimated from the area under the conformational map for. different pairs of amino acids in a subfibril containing five quarter-staggered collagen molecules. The plot of flexibility (number of conformations, a unitless variable) vs band number shown indicates that regions e2, e1, d, and c3 are continuously flexible. Other bands such as b2 and a1. are flexible but are separated by relatively rigid interband regions.")
64
Figure 7. Diagrammatic representation of calcium and phosphate ion binding to a type I collagen flexible region located in the d band of the fibril. The model depicts possible localizations of calcium and phosphate ions in a region devoid of proline and hydroxyproline. The addition of calcium and phosphate ions in this region of the molecule may increase the axial rise per residue and stretch the D-period of the fibril. This figure was constructed using a SYBYL molecular modeling program and the type I collagen amino acid sequence between residues 414 and 424 after addition of calcium and phosphate ions and energy minimization

65
(5) Silver, F. H. ; Christiansen, D. ; Snowhill, P. B. ; Chen. Y
(5) Silver, F. H.; Christiansen, D.; Snowhill, P. B.; Chen. Y.; Landis, W. J. The role of mineral in the storage of elastic energy in turkey tendons. Biomacromolecules 2000, 1, (6) Silver, F. H.; Freeman, J.; Seehra, G. P. Collagen self-assembly and development of matrix mechanical properties. Submitted for publication. (7) Silver, F. H. Biological Materials: Structure, Mechanical Properties and Modeling of Soft Tissues; New York University Press: New York, 1987; pp 7-21, , (8) Engle, J. Versatile collagens in invertebrates. Science 1997, 277, (9) Hofmann, H.; Voss, T.; Kuhn, K.; Engle, J. Localization of flexible sites in threadlike molecules from electron micrographs: Comparison of interstitial, basement membrane and intimal collagens. J. Mol. Biol. 1984, 172, (10) Silver, F. H.; Birk, D. E. Molecular structure of collagen in solution: comparison of types I, II, III, and V. Int. J. Biol. Macromol. 1984, 6, (11) Landis, W. J.; LiBrizzi, J. J.; Dunn, M. G.; Silver, F. H. A study of the relationship between mineral content and mechanical properties of turkey gastrocnemius tendon. J. Bone Mineral. Res. 1995, 10,
 Silver, F. H.; Christiansen, D.; Snowhill, P. B.; Chen. Y.; Landis, W. J. The role of mineral in the storage of elastic energy in turkey. tendons. Biomacromolecules 2000, 1, (6) Silver, F. H.; Freeman, J.; Seehra, G. P. Collagen self-assembly and. development of matrix mechanical properties. Submitted for publication. (7) Silver, F. H. Biological Materials: Structure, Mechanical Properties. and Modeling of Soft Tissues; New York University Press: New. York, 1987; pp 7-21, , (8) Engle, J. Versatile collagens in invertebrates. Science 1997, 277, (9) Hofmann, H.; Voss, T.; Kuhn, K.; Engle, J. Localization of flexible. sites in threadlike molecules from electron micrographs: Comparison. of interstitial, basement membrane and intimal collagens. J. Mol. Biol. 1984, 172, (10) Silver, F. H.; Birk, D. E. Molecular structure of collagen in. solution: comparison of types I, II, III, and V. Int. J. Biol. Macromol. 1984, 6, (11) Landis, W. J.; LiBrizzi, J. J.; Dunn, M. G.; Silver, F. H. A study of. the relationship between mineral content and mechanical properties. of turkey gastrocnemius tendon. J. Bone Mineral. Res. 1995, 10,")
69
Az ín szerkezete
















 n Áttekintés n Példák.>")







