Előadást letölteni
1
Virulens/intemperált bakteriofágok
Fág vizsgálati módszerek A T4 fág Más virulens fágok T sorozat ssDNS vírusok RNS fágok Bacillus subtilis fágok

2
Fizikai módszerek Biológiai módszerek Titrálás (plakk)
Elektron mikroszkópia Hibridizálás heteroduplex analízis Radioaktív jelölés Biológiai módszerek Titrálás (plakk) Egylépéses növekedés (one step growth) Single burst Korai lízis (premature lysis)
 Egylépéses növekedés (one step growth) Single burst. Korai lízis (premature lysis)")
3
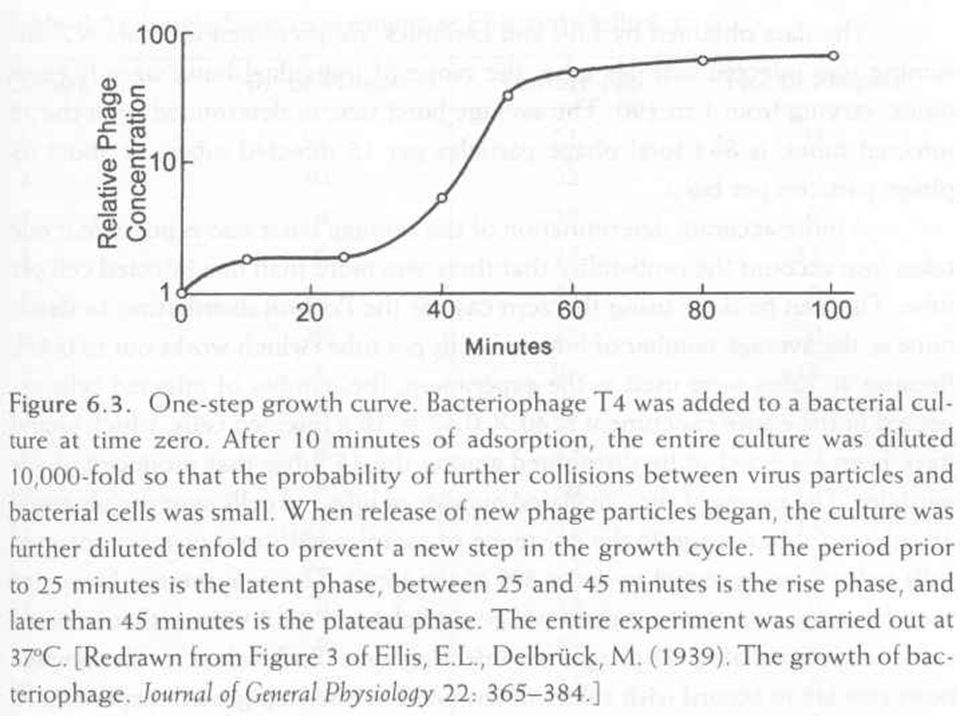
Egylépéses növekedési görbe
Egyidejű fáginfekció Fág adszorpció gátlás Növekedést gátló anyag (KCN) Különböző időkben vizsgáljuk a infekciós centrumok számát (titrálás, plakk) Három fázisú görbe Látens fázis Az infekciós centrum a fertőzött sejt (amely lizál) Emelkedési szakasz A sejtek kezdenek lizálni, az infekciós centrumok vagy a kiszabadult virionok, vagy a fertőzött sejtek Plató Az infekciós centrum a szabad virion
 Különböző időkben vizsgáljuk a infekciós centrumok számát (titrálás, plakk) Három fázisú görbe. Látens fázis. Az infekciós centrum a fertőzött sejt (amely lizál) Emelkedési szakasz. A sejtek kezdenek lizálni, az infekciós centrumok vagy a kiszabadult virionok, vagy a fertőzött sejtek. Plató. Az infekciós centrum a szabad virion.")
5
Az átlagos „burst” méret, az egy sejtből kiszabadult virionok száma
A fenti görbéből a plató és látens fázis közötti arány az átlagos „burst size”

6
„Single burst” kísérlet
A fágfertőzést követően a tápközeget higítjuk, hogy egy csőbe csak egy fertőzött sejt kerüljön Így a lízis után a virionok csak egy fertőzött sejtből származnak A fertőzési arány (MOI<1) kicsi Fertőzés után higítjuk a sejteket A kiszabaduló virionok nem találkoznak gazda sejttel Nagy különbségek 1-58-ig Komplex folyamat eredménye a „burst size”
 kicsi. Fertőzés után higítjuk a sejteket. A kiszabaduló virionok nem találkoznak gazda sejttel. Nagy különbségek 1-58-ig. Komplex folyamat eredménye a „burst size")
7
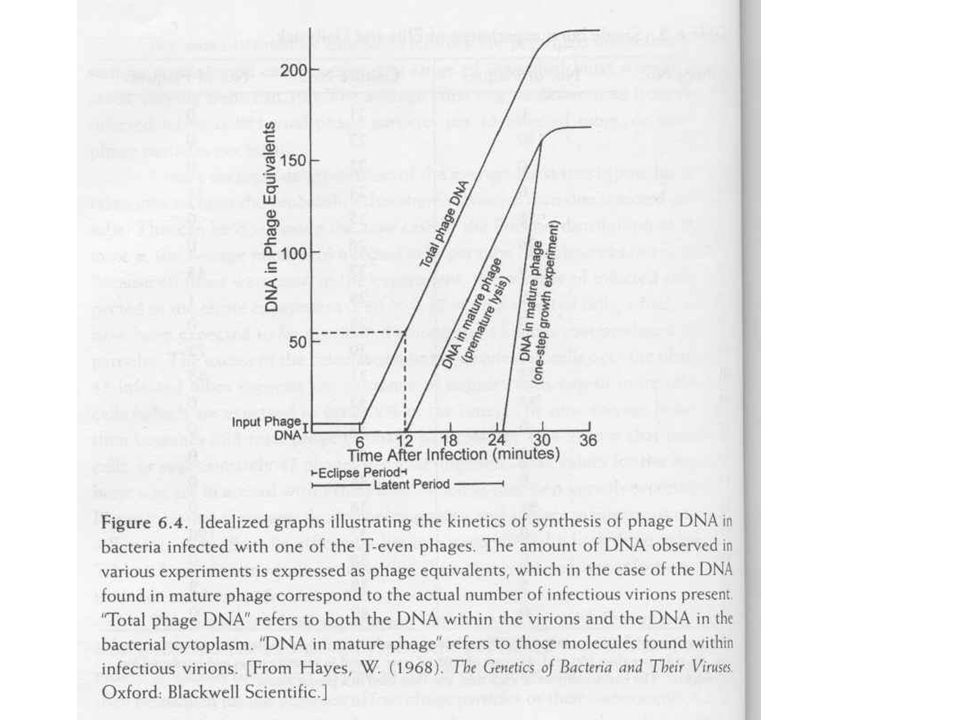
Korai lízis (megszakított fertőzés)
Módosított egylépéses növekedés Mintavétel után lizáljuk a sejteket Kloroformos rázatás T6 felülfertőzés (100 MOI), lízis kívülről Vizsgáljuk a kiszabadult fág részecskéket, vagy fehérje komponenseket Eklipsz fázis A növekedési szakasz azon része, amikor nincs fertőző fágrészecske A fág DNS nem fertőző A fágrészecskék megjelenése után a részecskék számának növekedése lineáris Egyenként keletkeznek a részecskék, mint egy gyártósoron (nem osztódás)
, lízis kívülről. Vizsgáljuk a kiszabadult fág részecskéket, vagy fehérje komponenseket. Eklipsz fázis. A növekedési szakasz azon része, amikor nincs fertőző fágrészecske. A fág DNS nem fertőző. A fágrészecskék megjelenése után a részecskék számának növekedése lineáris. Egyenként keletkeznek a részecskék, mint egy gyártósoron (nem osztódás)")
9
T4 fág (genetikai analízis)
Mutáció, r+, RII, lízis gátlás<>rapid lízis Keresztezés Deléciós térképezés 2, 3 faktoros keresztezés Komplementáció Cisztron, egy komplementációs csoport egy fehérje Finomtérképezés Recon: a rekombináció legkisebb egysége, 1 bp Muton: a mutáció legkisebb egysége, 1bp A kód 3 bp-os

11
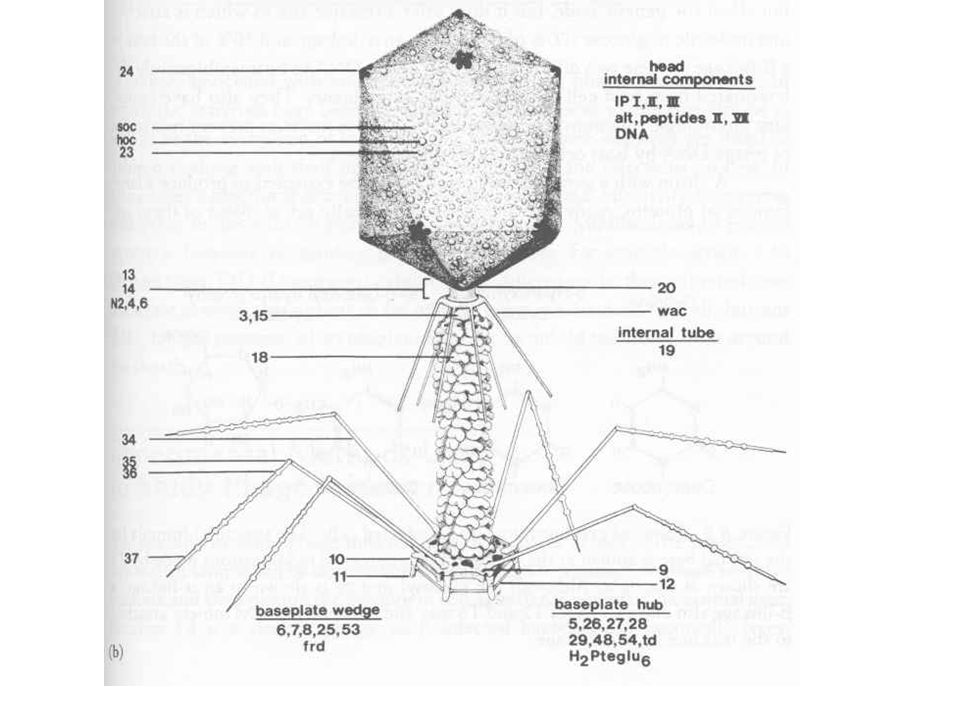
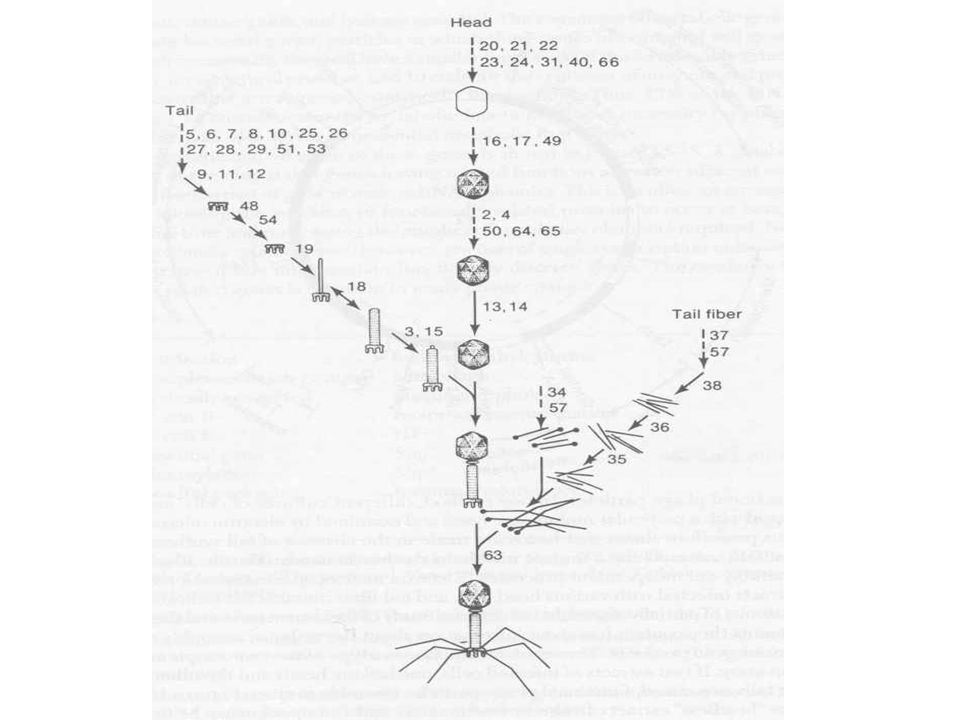
T4 morfológia és összetétel
Ikozaéder fej Kontraktilis/összehúzódó farok Farok szálak A fejben lineáris dupla szálú DNS

14
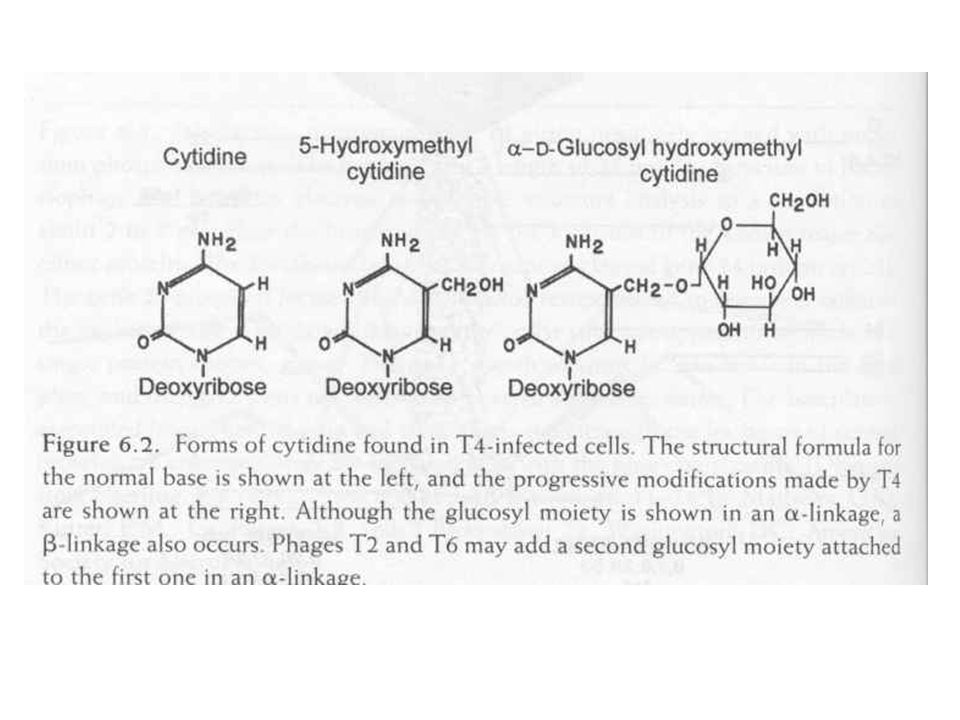
T4 DNS (morfológia és összetétel)
Hidroximetil-citozin Metil csoport glükozilált 70 % kötéssel 30 % kötéssel Így különbözteti meg a saját és gazda DNS-t

16
Fág heterozigóták Het Terminális redundancia
RII mutáns vizsgálatkor 2%-ban „molyhos” plakk, keverék populáció, amely szegregálódik (RII és r+) Vagy rekombináció eredménye vagy terminális redundancia Terminális redundancia Fej nagyobb, mint a DNS 3 kb-sal több DNS pakolódik A végen ismétlődik a DNS Fizikai módszer, DNS denaturálás után „ragadós vég keletkezik”
 Vagy rekombináció eredménye vagy terminális redundancia. Terminális redundancia. Fej nagyobb, mint a DNS. 3 kb-sal több DNS pakolódik. A végen ismétlődik a DNS. Fizikai módszer, DNS denaturálás után „ragadós vég keletkezik")
18
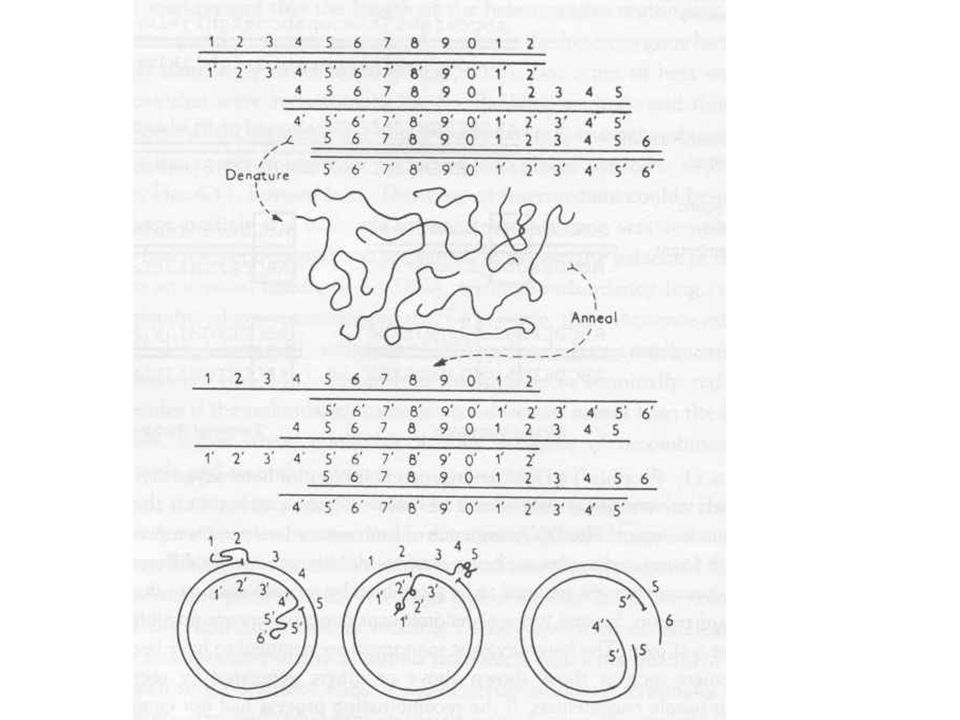
Cirkuláris permutáció
Cirkuláris géntérkép, de lineáris molekula Mintha egy kör alakú molekulát különböző helyeken vágnánk fel Random duplex kialakítás Bizonyíték, hogy a molekula cirkulárisan permutált A terminális redundancia és cirkuláris permutáció megmagyarázza, hogy miért cirkuláris a géntérkép Mert nincsen „fix kezdőpontja” a DNS-nek, ezért a géntérképnek sem lehet

20
T4 DNS replikáció Terminális redundancia Cikuláris permutáció Modell
Konkatemerek keletkeznek, melyek a fág fejbe pakolódnak a fej nagysága szerint (több, mint egy genom egység) Cikuláris permutáció Rolling circle replikáció jó lenne, de a T4 fág DNS nem cirkularizálódik Modell Lineáris molekula replikációja nem teljes A lagging strand vége nem tud replikálódni, egyszálú DNS marad, ami rekombinogén
 Cikuláris permutáció. Rolling circle replikáció jó lenne, de a T4 fág DNS nem cirkularizálódik. Modell. Lineáris molekula replikációja nem teljes. A lagging strand vége nem tud replikálódni, egyszálú DNS marad, ami rekombinogén.")
22
T4 adszorpció Farok szálak reverzibilisen kötődnek
Fág mozog a külső membránon Irreverzibilis kötődés a fág receptoron Adhéziós zóna (belső, külső membrán összeér) Farok összehúzódik Proton transzfer segítségével bejut a DNS
 Farok összehúzódik. Proton transzfer segítségével bejut a DNS.")
23
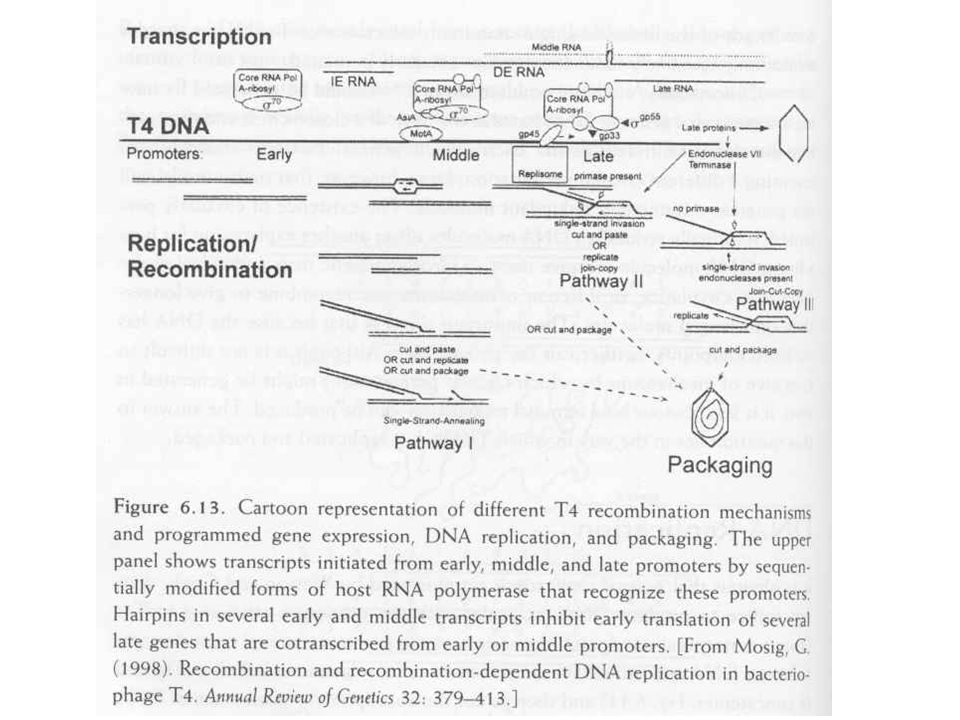
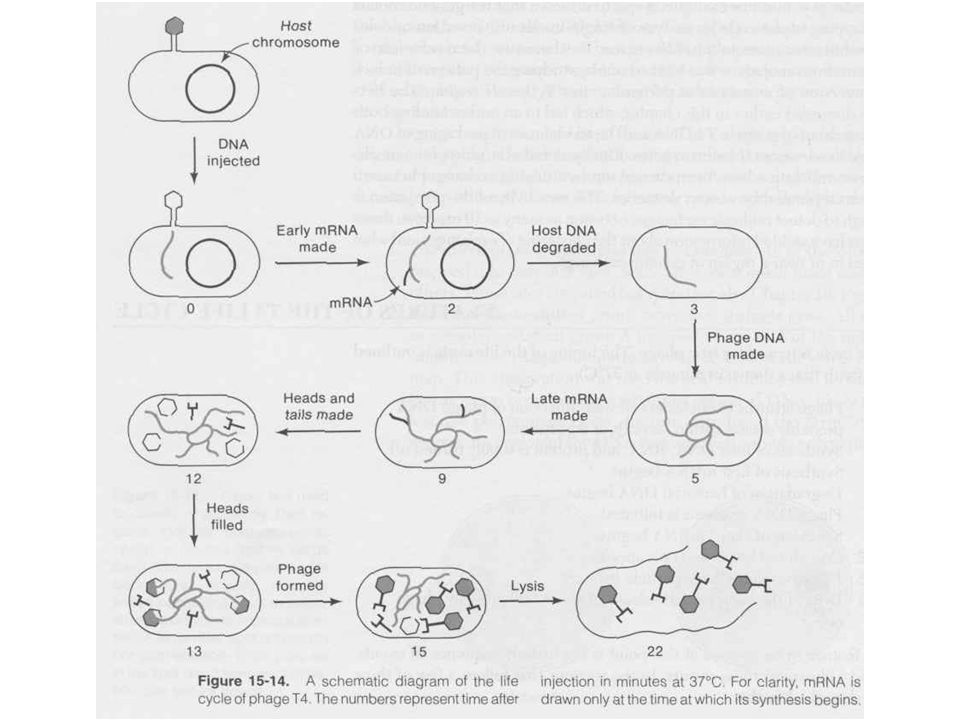
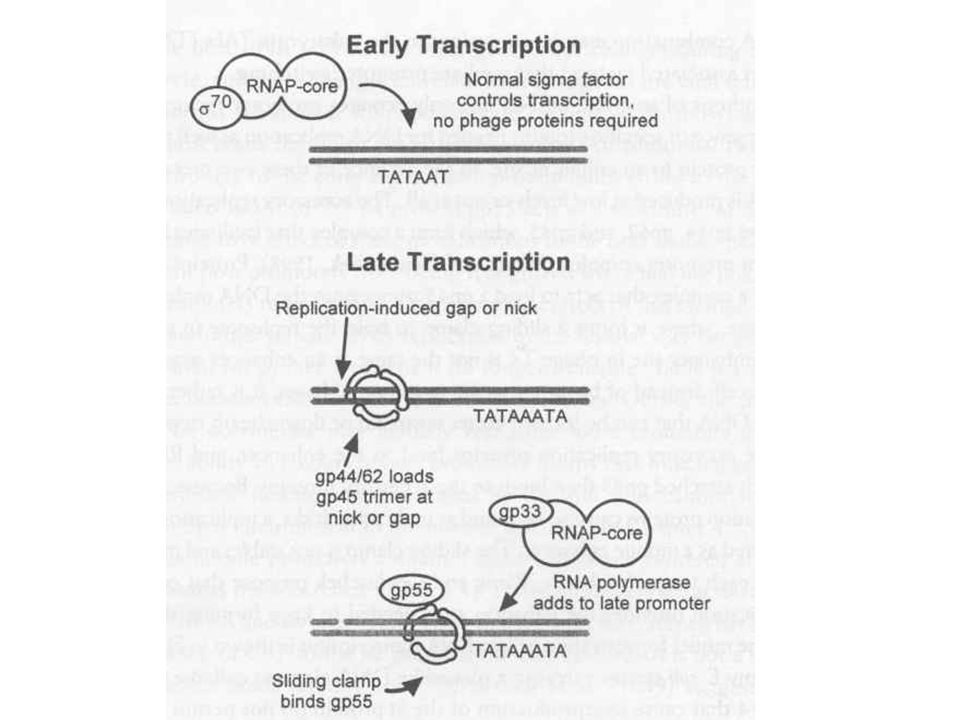
T4 szabályozás Szigorú szabályozás kell a megfelelő „burst size” eléréséhez Immediate early, early, middle, late gének Bekapcsolás a promóterek fizikai szeparációjával (késeiek a 3’ vég felé) Antiterminációval, azaz a termináció gátlásával
 Antiterminációval, azaz a termináció gátlásával.")
25
A T4 átveszi az irányítást
A gazda DNS degradációja Saját DNS replikációjának az alapanyaga HMC (hidroximetil-citozin) szintézis E. coli-ban nincs hasonló Citozin beépülésének megakadályozása T4 dCTPáz, bontja dCTP, dCDP, dCMP keletkezik, ami nem épül be a DNS-be A T4 DNS glükozilálása HMC-t a coli megtámadja HMC glükozilálódik T4 enzimek által, ezt nem támadja meg a coli endonukleáz RNS polimeráz módosítás (T4 szigma faktorok)
 szintézis. E. coli-ban nincs hasonló. Citozin beépülésének megakadályozása. T4 dCTPáz, bontja dCTP, dCDP, dCMP keletkezik, ami nem épül be a DNS-be. A T4 DNS glükozilálása. HMC-t a coli megtámadja. HMC glükozilálódik T4 enzimek által, ezt nem támadja meg a coli endonukleáz. RNS polimeráz módosítás (T4 szigma faktorok)")
28
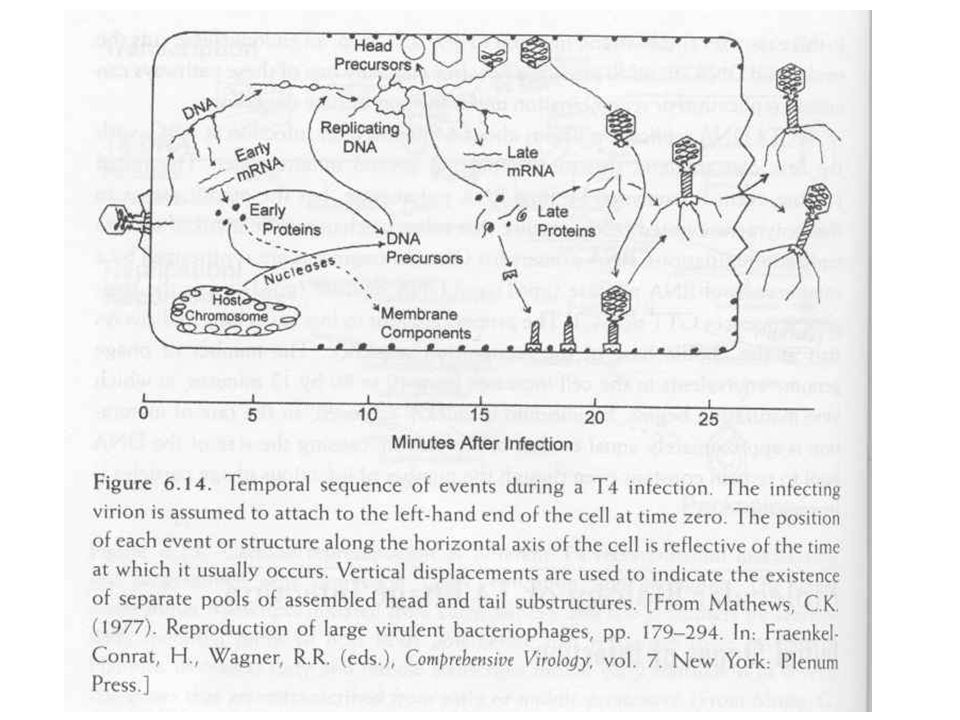
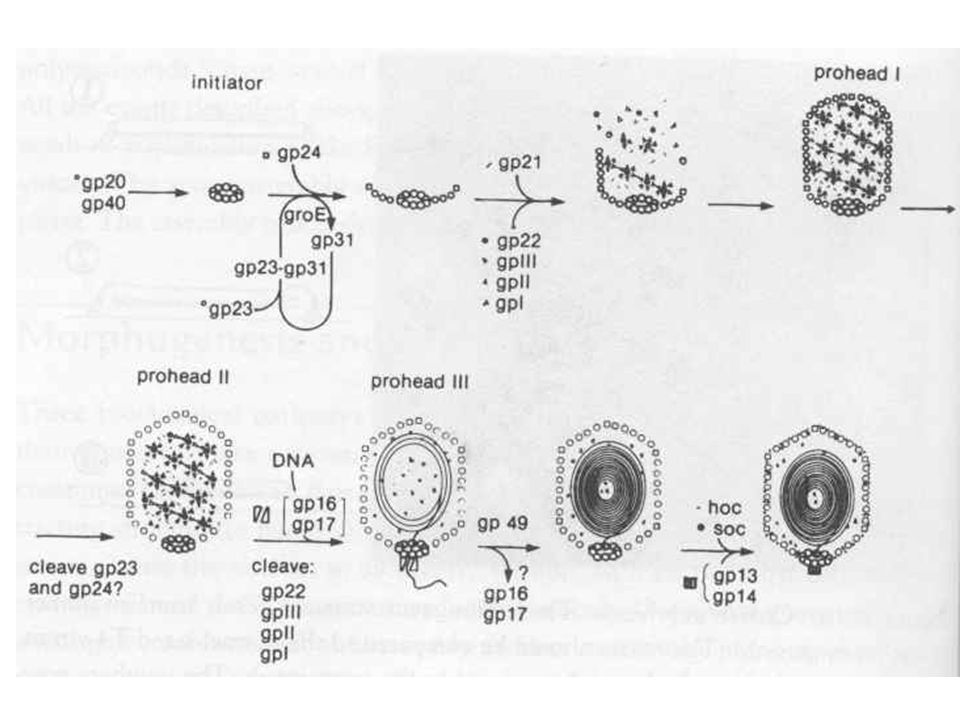
T4 fágrészecskék termelése
Fág alkotórészek szintézise Fej, farok Ahogy a premature lízisből látszott a fej, farok szintézise független folyamat A vírusrészecskék száma lineárisan nő, mintha „gyártósoron” készülnének Fej szintézisében chaperon fehérjék Fág DNS pakolás Fejméretű pakolás, endonukleáz hasítás, termináz enzim Spirális összecsomagolás (ion etching-el kimutatva) Gazdasejt lízis, fágspecifikus lizozim enzimmel
 Gazdasejt lízis, fágspecifikus lizozim enzimmel.")
32
Más virulens fágok T sorozat
Delbrück 7 különböző fágot választott Elektronmikroszkóppal a T párosak és T páratlanok hasonlítottak egymáshoz T páros, kontraktilis farok, hosszúkás fej T páratlan, nem kotraktilis farok, oktaéderes fej

33
T2, T6 fágok A T4 fággal homológok HMC glükoziláció különbözik
T2 25% nem glükozilált, di-glükozidok (minkettő ) T6 di-glükozidos, az első , a második glükozidos kötéssel 85% homológia T2 host range mutáció Farok szálak módosulnak E. coli mutálódhat T3, T7-nél host range mutációra rezisztens baktérium T2 farok szál poszt-transzlációsan módosul (180 AA levágás a C-terminálisról Ha hiányzik a szignál a fehérjéről, akkor nincs módosítás
 T6 di-glükozidos, az első , a második glükozidos kötéssel. 85% homológia. T2 host range mutáció. Farok szálak módosulnak. E. coli mutálódhat. T3, T7-nél host range mutációra rezisztens baktérium. T2 farok szál poszt-transzlációsan módosul (180 AA levágás a C-terminálisról. Ha hiányzik a szignál a fehérjéről, akkor nincs módosítás.")
34
T1 fág Legkorábban használt fág (Luria-Delbrück fluktuációs teszt)
Perzisztens, nehéz kiirtani, mindenhol megjelenik, ezért nem vizsgálták Valószínű a fág virulens mutánsa T1 fág mutánsok története (PCR hasonló)
")
35
T5 fág T1-hez hasonlít, kevesebb farok szál (4)
Nagy (9%) terminális redundancia Nick-ek a DNS-en több 4-5-ös sorozatban Gének szabályozása (5’-től a 3’-ig, pre-early, early, late) A pre-early gének jutnak be előbb DNS bejutás megáll Csak akkor indul újra, ha a már bejutott gének FST (first step transfer) expresszálódnak Néhány T5 promóter nagyon erős (90% az össz transzkripciónak innen indul), 7-metil-guanozin (CAP), klónozó vektorokban használt
 terminális redundancia. Nick-ek a DNS-en több 4-5-ös sorozatban. Gének szabályozása (5’-től a 3’-ig, pre-early, early, late) A pre-early gének jutnak be előbb. DNS bejutás megáll. Csak akkor indul újra, ha a már bejutott gének FST (first step transfer) expresszálódnak. Néhány T5 promóter nagyon erős (90% az össz transzkripciónak innen indul), 7-metil-guanozin (CAP), klónozó vektorokban használt.")
36
T7, T3 T7 RNS polimeráz csak a saját promóterét ismeri fel
T7 promóter egy része nagyon konzervált így valószínűtlen, hogy máshol is előfordul Klónozó vektorokban használják, mert a T7 promóterről csak akkor van átírás, ha a T7 polimeráz is a sejtben van T7 terminálisan redundáns, de cirkulárisan nem permutált (hasítás restrickiós enzimhez hasonló) Fejméretű pakolás nem magyarázza az egyedi DNS molekulát
 Fejméretű pakolás nem magyarázza az egyedi DNS molekulát.")
37
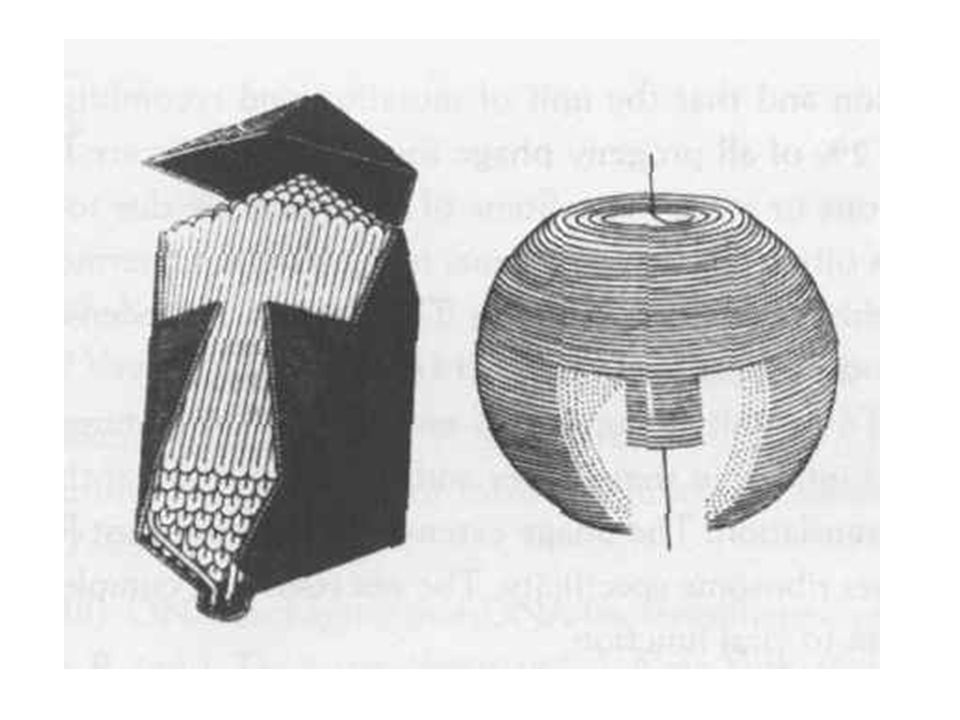
Egyszálú DNS-t tartalmazó fágok (Ff, filamens fág csoport)
Cirkuláris egyszálú DNS (~6 kb) Hosszúkás alak, melyben a DNS köré sok azonos alegységű fehérje tekeredik F pílushoz tapadnak a virionok (male specifikus fágok)
 Hosszúkás alak, melyben a DNS köré sok azonos alegységű fehérje tekeredik. F pílushoz tapadnak a virionok (male specifikus fágok)")
38
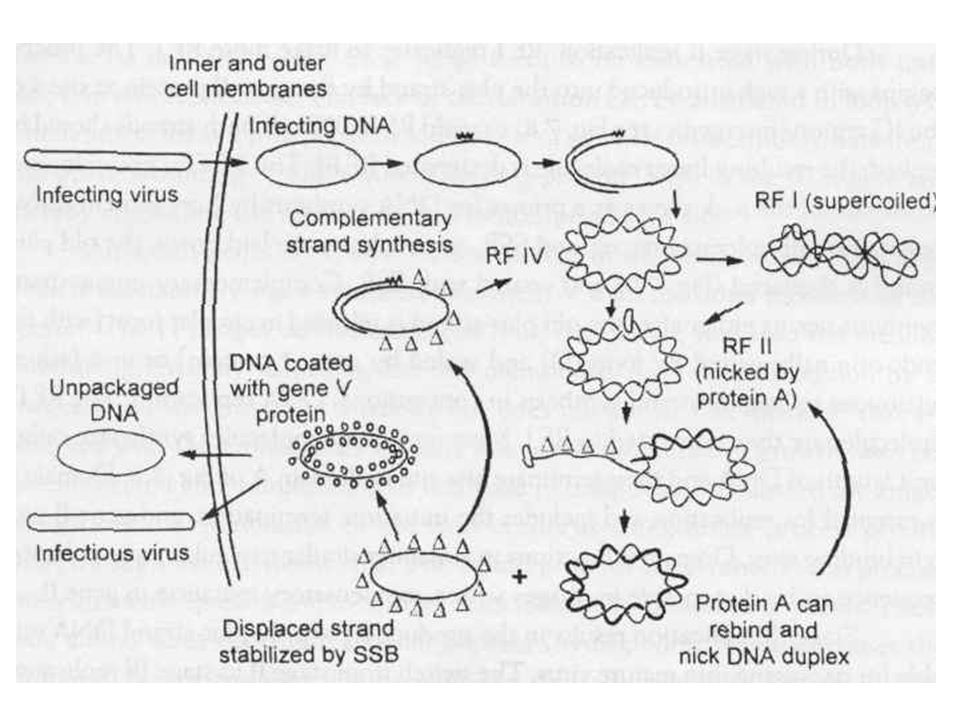
Ff DNS replikáció RNS priming RF IV forma kovalensen zárt cirkuláris
RFI supercoiled RF IV RFII nick a DNS-en Komplementer szál szintetizálódik Az eredeti szál áthelyeződik SSB (egyszálú DNS kötő fehérje) stabilizálja „pakolódik” Kijutás szignál peptid akcióként, nincs lízis Nagy titer 1012 /ml, plakk lassúbb növekedésű rész
 stabilizálja. „pakolódik Kijutás szignál peptid akcióként, nincs lízis. Nagy titer 1012 /ml, plakk lassúbb növekedésű rész.")
40
M13 fág lacZ gén beépítése a klónozáshoz
Restrikciós helyek a klónozáshoz Primer hely RF használható, mint egy plazmid klónozás Egyszálú DNS szekvenáláshoz használható

41
X174 Sejt lízis Kis genom méret, egymásba ágyazott gének

43
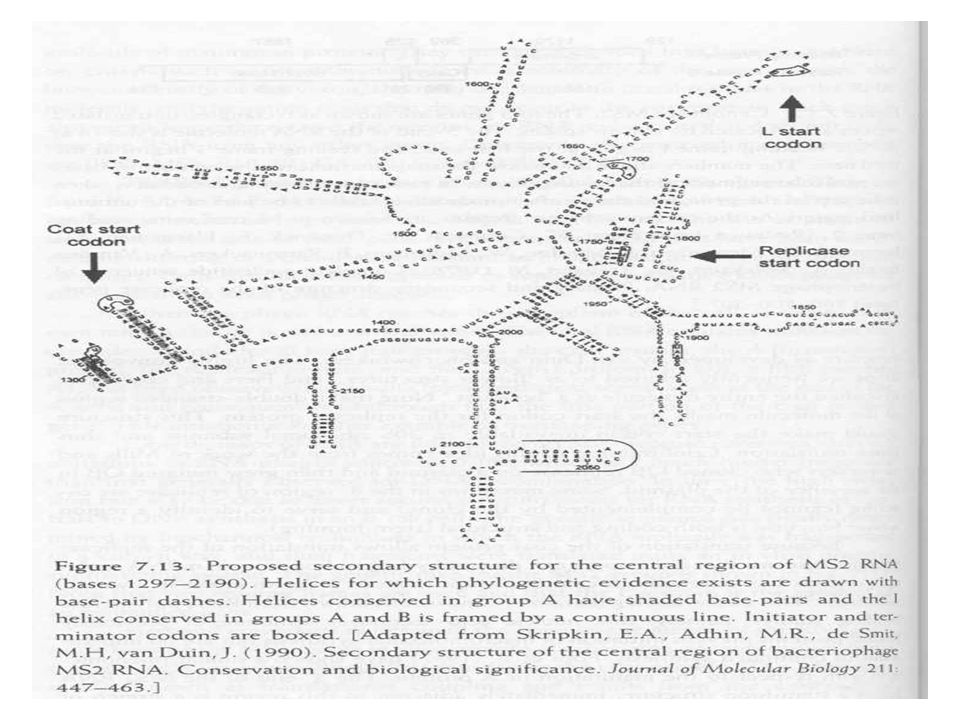
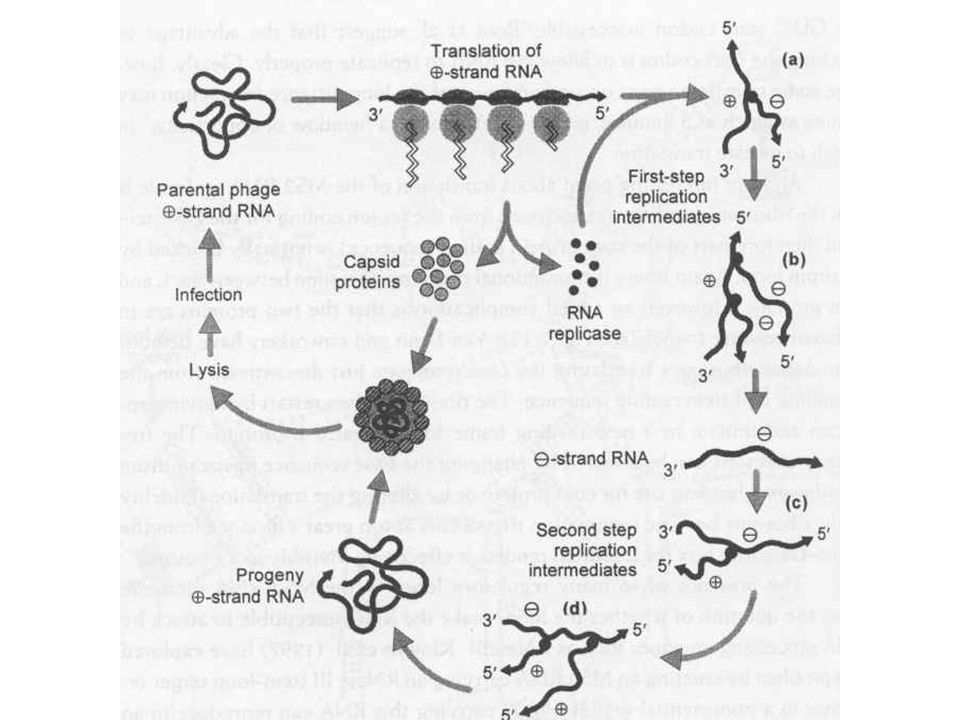
RNS fágok Lineáris egyszálú RNS
Male specifkus fágok, píluson keresztül Nagy reverziós ráta, mert az RNS szintézis pontossága kisebb Transzlációs csatolás RNS másodlagos szerkezetéből adódik (replikáz csak a köpenyfehérje transzlációja után indulhat RNS replikációhoz 4 fehérje szükséges 3 gazda fehérje mely az RNS transzlációs rendszerben található Virális replikáz Ez a komplex az RNS függő RNS polimeráz, két lépcsős replikáció, először a + szál, majd a – szál szintetizálódik

46
Bacillus subtilis fágok
SPO1 T4-hez hasonló morfológia Terminálisan redundáns, de egyedülálló szekvencia Timin helyett 5-hidroxi-metil-uracilt tartalmaz A gazda RNS polimeráz megváltozik a fág szigma faktorok hatására 2 replikációs origó van, 20x genomnyi konkatemer jöhet létre A gazda sporulációja gátolja a SPO1 fejlődését, ezért az endospórában virális DNS található, amely lehetővé teszi a szaporodást

47
29 Kisméretű dupla szálú DNS, gyűrűs
Két korai gén a DNS két végén (cirkuláris DNS!) DNS középső részén Az átmenet a gazda transzkripcióból a fág transzkripcióba, fág szigma faktorok termelődése révén valósul meg
 DNS középső részén. Az átmenet a gazda transzkripcióból a fág transzkripcióba, fág szigma faktorok termelődése révén valósul meg.")
48
Összefoglalás Fág vizsgálati módszerek T4 fág T sorozat fágjai
Egyszálú DNS fágok RNS fágok Bacillus fágok









>")


 szív vizsgálat>")



 – méreg gör.- phag Dimitrij Ivanovszkij>")


 „késői” gének (late - L) 4.7 kb SV40 genom - kicsiny „tanulóvírus” fertőzést követően először.>")


>")

