Előadást letölteni
Az előadás letöltése folymat van. Kérjük, várjon

1
A HALLÓRENDSZER PPKE ITK, 2007

4
Sound as mechanical wave energy requires a medium such as air or water in which to move.
Sound: vibratory energy caused by movement of physical objects Rate of vibration is called frequency What we hear is pitch (high or low) We hear 20-20,000 Hz (cycles/sec) Size (intensity) of vibration is amplitude What we experience is loudness Measured in decibels (dB)
 We hear 20-20,000 Hz (cycles/sec) Size (intensity) of vibration is amplitude. What we experience is loudness. Measured in decibels (dB)")
5
Additive synthesis & Fourier analysis
As in Fourier analysis of patterns of light, the same method can be used for representing and constructing complex sound wave phenomena. Here (d) is a composite of the fundamental (a) plus its second and third harmonics, (b) and (c).
 is a composite of the fundamental (a) plus its second and third harmonics, (b) and (c).")
6
AZ EMBERI HALLÁS FREKVENCIATARTOMÁNYA
kHz -AZ EMBERI HALLÁS 1-4 kHz frekvenciájú rezgésekre a legérzékenyebb- Ekkor az ingerküszöb~ 0 dB=20 µPa

8
A KÖZÉPFÜL KÉPLETEI Kalapács Üllő Kengyel Dobhártya Kerek ablak
Eustach kürt Hallójárat: 25-27 mm. Rezonancia: Hz. Erősítés: 10-15dB

9
A HALLÓCSONTOK MŰKÖDÉSE
Középfül funkciója: Akusztikus impedanciaillesztés: dobhártya-ovális ablak felületének aránya: 17:1 csontok: 1,3x áttételű emelő Σ x erősítés 25-30 dB nyomásnövekedés Dobhártya mozgás: 2 dB suttogás = 10-8 mm

10
Csontvezetés Középfül megkerülése Saját hang felismerése
Vezetéses típusú halláscsökkenésnél a hangrezgések ezen az úton továbbíthatóak a belső fülbe

11
Ossicular reflex (1) Malleus ; - (2) Malleus ligament ; - (3) Incus ; - (4) Incus ligament; - (5) Stapes muscle (stapedius); - (6) Stapes footplate; - (7) Eardrum; - (8) Eustachian tube; - (9) Malleus muscle (tensor tympani); - (10) Nerve (chorda tympani) sectioned
 Malleus ; - (2) Malleus ligament ; - (3) Incus ; - (4) Incus ligament; - (5) Stapes muscle (stapedius); - (6) Stapes footplate; - (7) Eardrum; - (8) Eustachian tube; - (9) Malleus muscle (tensor tympani); - (10) Nerve (chorda tympani) sectioned.")
12
Musculus tensor tympani és a musculus stapedius erős hangok hatására reflexesen összehúzódnak
Bilaterális reflex Hosszú latencia idő Alacsony frekvenciás hangok szűrése-ezáltal a saját hang kiemelése

13
The inner ear Oval window 0,5 mm Round window Cochlea from a human fetus (5 months of gestation)
")
14
A COCHLEA KERESZTMETSZETE
Ductus cochlearis Scala vestibuli Scala tympani Ganglion spirale Nervus acusticus

15
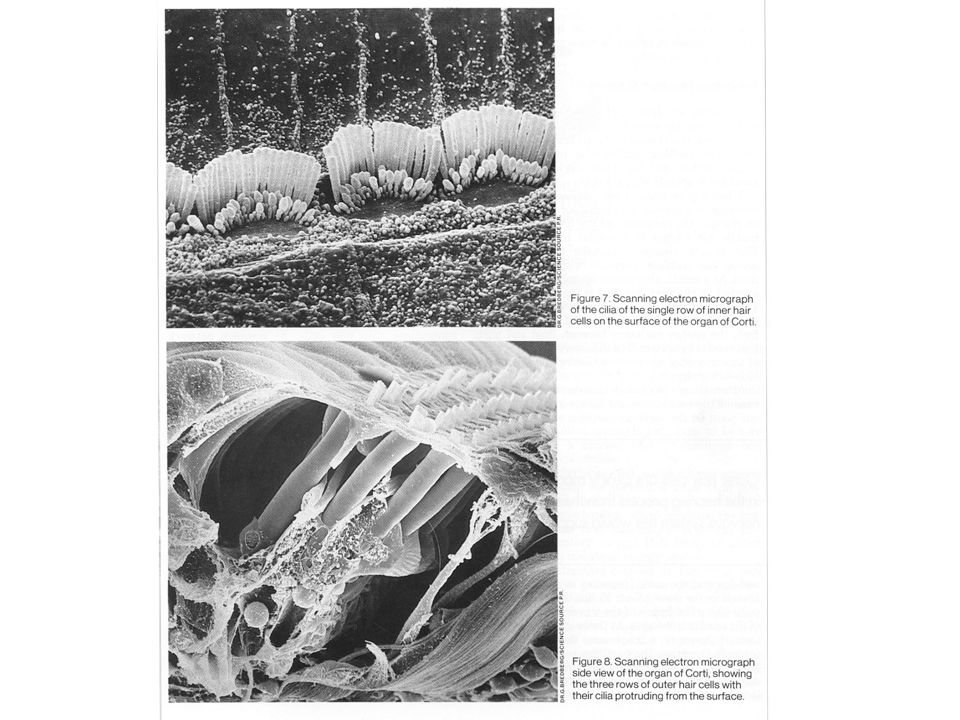
A CORTI SZERV ~15.00 szörsejt 30.000 idegrost ~ 3.500 belső szőrsejt
Az idegrostok 95%-a végződik rajtuk. Szenzoros funkció ~ külső szőrsejt (3 sorban) Az idegrostok 5%-a Hangoló, erősítő funkció
 Az idegrostok 5%-a. Hangoló, erősítő funkció.")
16
Organ of corti Copyright © 2002 Wadsworth Group. Wadsworth is an imprint of the Wadsworth Group, a division of Thomson Learning

17
BELSŐ SZŐRSEJT KÜLSŐ SZŐRSEJT Sejtmag Stereocilia Cuticularis lemez
Radiális afferens idegvégződés (I. típus) Laterális efferens idegvégződés Mediális efferens idegvégződés Spirális afferens idegvégződés (II. típus) KÜLSŐ SZŐRSEJT
 Laterális efferens idegvégződés. Mediális efferens idegvégződés. Spirális afferens idegvégződés. (II. típus) KÜLSŐ SZŐRSEJT.")
18
Hair cells The apical surface of inner support cells, one row of inner hair cells (i: IHCs), pillars of Corti (p), 3 rows of outer hair cells (o: OHCs) and Deiters' cells are visible. At the bottom the surface is fractured and the base of the 3rd row of OHCs is seen, as well as phalangeal processes of Deiters' cell (d). scale bar: 15 µm
, pillars of Corti (p), 3 rows of outer hair cells (o: OHCs) and Deiters cells are visible. At the bottom the surface is fractured and the base of the 3rd row of OHCs is seen, as well as phalangeal processes of Deiters cell (d). scale bar: 15 µm.")
20
Tonotopia-hely elv, frekvenciaanalízis
Cochlea frekvencia analizáló működésének alapja a tonotopia A ductus cochlearis/ membrana basilaris hosszában egymást követő szőrsejtek ingerülete különböző frekvenciájú hangrezgéseket jelez a kp. idegrendszer számára Foramen ovale- magasabb hang/ nagyobb ferkvencia Helicotrema- mélyebb hang/ kisebb ferkvencia

21
Tonotopia 1. cochlea passzív analizátor funkciója
2. külső szőrsejtek aktív funkciója, bidirekcionális transzdukciós mechanizmus 3. az eltérő pozíciójú szőrsejtek különböző mechanikai hangolása

22
1. Passzív frekvencia analízis

23
Membrana basilaris a hangok passzív frekvenciaanalízisét végzi
A kengyel talpa a foramen ovalében mozog Nyomás különbség a S.V. és S.T. között Membrana basilarison vándorló hullám keletkezik Membrana basilaris szerkezete szabja meg, hogy adott frekvenciájú rezgésnél hol mutat maximumot a kilengés Kitérés a hang intenzitásával arányos

24
As shown in this schematic drawing, as the frequency varies from high to low, the site of maximum displacement of the basilar membrane shifts toward the apex. This corresponds to a passive tonotopy .

25
The wave displayed here, generated on the basilar membrane by a 400 Hz input tone, is the solution of an integro-differential equation which describes the membrane motion in the linear approximation.Waves peak at a frequency-dependent location and form because: i) each membrane segment interacts instantly with each other through the fluid filling the cochlear duct and ii) membrane stiffness is graded from base to apex.
 each membrane segment interacts instantly with each other through the fluid filling the cochlear duct and ii) membrane stiffness is graded from base to apex.")
26
2. A KÜLSŐ SZŐRSEJTEK AKTÍV MŰKÖDÉSÉNEK HATÁSA
10 kHz-es tiszta hang által okozott haladó hullám tengeri malac membrana basilarisán. A külső szőrsejtek kb. ½ oktávval eltolják a hullám maximumának helyét és kb. 50 dB-el növelik amplitúdóját.

27
Four-image set of the cochlear partition (resolution 356 nm/pixel) viewed from scala media at angle bringing full length of outer hair cells into focus. Cell stereocilia are visible, through the intact tectorial membrane, near frame bottom. A patch pipette (entering from R.) is attached to a third row outer hair cell, incrementing cell potential by 25 mV between successive frames. The cell responds by shortening visibly when depolarized within the intact organ of Corti. The mechanism depends on an integral membrane protein called prestin acting as a molecular motor.
![]()
28
Four-image set of the cochlear partition (256x256 pixels, resolution 356 nm/pixel) viewed from scala media at angle bringing full length of outer hair cells into focus. Cell stereocilia are visible, through the intact tectorial membrane, near frame bottom. A patch pipette (entering from R.) is attached to a third row outer hair cell, incrementing cell potential by 25 mV between successive frames. The cell responds by shortening visibly when depolarised within the intact organ of Corti.
![]()
29
Active movement of the OHC
Displacement of the stereocilia towards the tallest produces a cell depolarisation as ionic current flows in through the mechanoelectric transducer channels. The outer hair cell responds by changing length (vertical arrows). Ultrastructural analysis of cell lateral plasma membrane indicates that electromotility may be based upon conformational changes of a dense array of integral membrane proteins whose molecular identity is not yet known. Changes in membrane potential may act as the driving force for the re-arrangement of the proteins in the plane of the membrane. Membrane surface forces are converted into cell axial forces by interaction of the motor proteins with the cytoskeleton (inset). . |
. Ultrastructural analysis of cell lateral plasma membrane indicates that electromotility may be based upon conformational changes of a dense array of integral membrane proteins whose molecular identity is not yet known. Changes in membrane potential may act as the driving force for the re-arrangement of the proteins in the plane of the membrane. Membrane surface forces are converted into cell axial forces by interaction of the motor proteins with the cytoskeleton (inset). . |")
30
Tip links

31
Coupling between outer hair cells and Deiters' cells Here we patch-clamped simultaneously one outer hair cell (OHC) in raw 3 of cochlear turn 3 and its attached Deiters' cell (DC). The pipette contacting the DC near the frame bottom was out of focus and does not show. 100 mV potentials steps were repeatedly applied to the OHC to study the mechanical behavior of the OHC-DC junction. It is quite clear that this cell couple was firmly connected and that the OHC did not slide in the DC cup while changing length. On the contrary, the force applied by the OHC produced a visible deformation in the DC, acting against its visco-elastic load

32
Organ of Corti and Internal Forces
Understanding how the organ of Corti reacts to cellular forces is central to cochlear micromechanics. One simple scheme is that, under outer hair cell contraction, the organ of Corti would distort as shown here. This hypothesis is consistent with experiments. Cell motility feeds back to the basilar membrane motion enhancing the travelling wave amplitude and making the cochlea active. Reference Mammano, F. and Nobili, R. (1993) J. Acoust. Soc. Am. 93, Organ of Corti and Internal Forces
 J. Acoust. Soc. Am. 93, Organ of Corti and Internal Forces.")
33
2) Mechano-transduction at OHCs.
Bending of OHC stereocilia leads to K+ entry and OHC depolarisation 3) Electromotility and reverse transduction. Depolarisation of its membrane results in OHC contraction which magnifies the motion 4) Mechano-transduction at IHC OHC contraction enhances the motion, IHC stereocilia are bent 5) IHC-auditory nerve synapse activation as a result of the IHC depolarisation: an auditory message is sent via the type I afferent (blue)
 Electromotility and reverse transduction. Depolarisation of its membrane results in OHC contraction which magnifies the motion. 4) Mechano-transduction at IHC OHC contraction enhances the motion, IHC stereocilia are bent. 5) IHC-auditory nerve synapse activation as a result of the IHC depolarisation: an auditory message is sent via the type I afferent (blue)")
34
A STEREOCILIUMOK MŰKÖDÉSE
Mechanoelektromos transzdukció: A stereocilium elhajlásakor a kation csatornák megnyílnak, K+ ion áramlik a sejtbe, a sejt depolarizálódik.

35
Oto-acoustic emissions
Discovered back in 1978, oto-acoustic emissions (OAEs) were not fully explained until a few years later, after the OHC active mechanism had been understood. At least in the mammalian cochlea, OAEs reflect OHC electromotility. OHC contractions/elongations themselves vibrate cochlear fluids and the middle ear conducting mechanism transfers this vibration back to the air of the external auditory canal: there, the emissions can be registered by a microphone. Schematic drawing of a probe for recording evoked OAEs. It contains a speaker emitting the stimulating sound and a microphone to record the returning OAEs
 were not fully explained until a few years later, after the OHC active mechanism had been understood. At least in the mammalian cochlea, OAEs reflect OHC electromotility. OHC contractions/elongations themselves vibrate cochlear fluids and the middle ear conducting mechanism transfers this vibration back to the air of the external auditory canal: there, the emissions can be registered by a microphone. Schematic drawing of a probe for recording evoked OAEs. It contains a speaker emitting the stimulating sound and a microphone to record the returning OAEs.")
36
3. Az eltérő pozíciójú szőrsejtek különböző mechanikai hangolása
A cochlea bázisához közelebbi szőrsejtek stereociliumai rövidebbek, feszesebbek-nagy frekvenciájú rezgések átvétele Apexhez közeli szőrsejtek stereociliumai hosszabbak, kevésbé feszesek-alacsonyabb frekvenciájú rezgésekre válaszolnak

37
Külső szőrsejtek: mechanoelektromos transzdukció, elektromechanikai transzdukció
Külső szőrsejtek alakváltozása elmozdítja a környező struktúrákat, elsősorban a membrana tectoriát-passzív hullám kiemelése A bidirekcionális transzdukció hat a belső szőrsejtekre is-nagyobb mértékű stereocilium kitérés Fiziológiás hallásküszöb a külső szőrsejtek erősítő funkciójának következménye (50-60dB SPL magasabb küszöb)
")
38
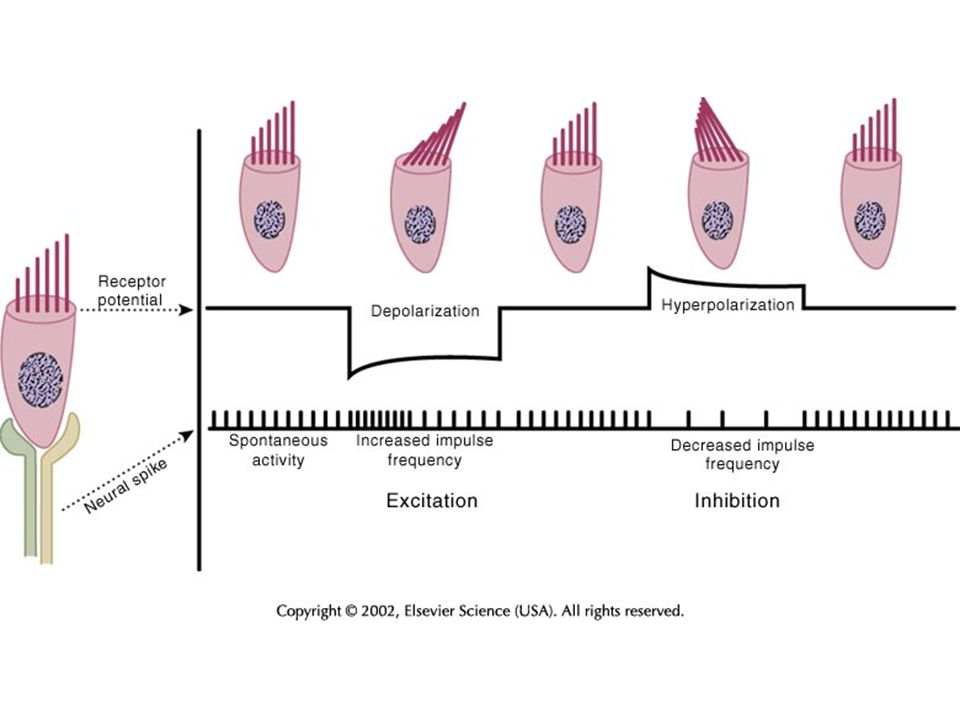
ELEKTROMOS POTENCIÁLOK A BELSŐ FÜLBEN
Nyugalmi potenciál A szőrsejtek nyugalmi potenciálja: -60 mV. A csatornák 15%-a nyitott. endolimpha - perilimphával = +80mV endolimpha – intracell. tér = ~150 mV Receptor potenciál A stereocilium 100nm kitérése megfelel a választerjedelem 90%-ának. Normális inger: ~±1° kitérés (kevesebb mint az átmérője) Termális zaj: 3nm. Hallásküszöb: 0,3 nm → 100μV receptor pot. .
 Termális zaj: 3nm. Hallásküszöb: 0,3 nm → 100μV receptor pot. .")
40
Overview of the function of the Organ of Corti
For low and mid intensities sounds the function of the organ of Corti may be schematically summarised in 5 stages: (1) Sounds waves move the basilar membrane (BM) up and down. (2) Stereocilia of the OHCs , implanted in the tectorial membrane (TM), are bent and the cell is depolarised. (3) Excited (depolarised) OHCs react by contracting (= electromotility): this is an active mechanism. (4) Due to the tight coupling of OHCs with the BM and reticular lamina this active mechanism feeds energy back into the organ of Corti and IHCs are excited possibly via TM (Hensen's stripe) activation of IHC stereocilia. (5) The IHC-auditory nerve synapse is activated and a message is sent to the CNS.
 Sounds waves move the basilar membrane (BM) up and down. (2) Stereocilia of the OHCs , implanted in the tectorial membrane (TM), are bent and the cell is depolarised. (3) Excited (depolarised) OHCs react by contracting (= electromotility): this is an active mechanism. (4) Due to the tight coupling of OHCs with the BM and reticular lamina this active mechanism feeds energy back into the organ of Corti and IHCs are excited possibly via TM (Hensen s stripe) activation of IHC stereocilia. (5) The IHC-auditory nerve synapse is activated and a message is sent to the CNS.")
41
Szenzoros transzdukció
Stereociliumok kitérése gradált –AC összetevő Membránpotenciál pozitívabb lesz- DC komponens Hanghullám-depolarizáció+rátevődött oszcilláció-pulzáló Ca beáramlás- változó intenzítású transzmitter leadás A nervus cochlearis akciós potenciál sorozatának frekvenciája a belső szőrsejtek transzmitter leadásától függ, karakterisztika hasonló

42
Belső szőrsejtek ingerülete a ganglion spirale bipoláris sejtjeinek végződéseire tevődik át
Egy oldal ~3500 belső szőrsejt Egy afferens rost egy szőrsejthez kapcsolódik Minden egyes szőrsejthez ~10 afferens rost kapcsolódik Legjobb frekvencia válasz jellemző Hangintenzitás növelése-karakterisztikus frekvencia feletti és alatti frekvenciák is afferens aktiválódást váltanak ki

43
Fáziscsatolás, frekvenciakód és populációs kód
AP Frekvencia maximum a hangnyomás változások maximumával esik egybe (!) Adott hangfrekvenciára válaszoló afferens idegek AP frekvenciája a hangnyomás szintől függ Nagyobb intenzitású hangok a membrana basilaris nagyobb kitérésével járnak-hangintenzitást emelve azon axonokban is fokozódik az AP frekvenciája amelyek karakterisztikus frekvenciája alacsonyabb ill. magasabb. Több ingerületbe került rost nagyobb hangintenzitást jelez- Populációs kód
 Adott hangfrekvenciára válaszoló afferens idegek AP frekvenciája a hangnyomás szintől függ. Nagyobb intenzitású hangok a membrana basilaris nagyobb kitérésével járnak-hangintenzitást emelve azon axonokban is fokozódik az AP frekvenciája amelyek karakterisztikus frekvenciája alacsonyabb ill. magasabb. Több ingerületbe került rost nagyobb hangintenzitást jelez- Populációs kód.")
44
SENSORY NEURONS ARE DRIVEN BY INNER HAIR CELLS
Phase Locking Consistent firing of cell at same sound wave phase for low frequency sounds

45
Phase-locking Inter-spike Intervals Response to Low Frequency tones
-1 -0.5 0.5 1 0.2 0.4 0.6 0.8 Inter-spike Intervals Response to Low Frequency tones time (t) Response to High Frequency tones > 5kHz Random intervals 2 periods 1 period nerve spike
 Response to High Frequency tones > 5kHz. Random intervals. 2 periods. 1 period. nerve spike.")
46
INDIVIDUAL NEURONS ARE TUNED TO RESPOND TO A NARROW RANGE OF FREQUENCIES

47
A központi idegrendszer a cochlea szintjén csökkentheti a hallás érzékenységét
Laterális olivocochleáris köteg: cochleáris afferensek érzékenységét állíthatják be Medialis olivocochleáris köteg: szintén az afferens aktivitást csökkentik, de a külső szőrsejtek bidirek-cionális transzdukciójának gátlásával Zajszűrés, kiemelés Erős hangoktól védelem PNS Fig 30-10

48
Auditory Nerve (VIII cranial nerve)
Neural information from inner hair cells: carried by cochlear division of the VIII Cranial Nerve. Bipolar neurons, cell bodies in spiral ganglion, proximal processes on hair cell, distal in cochlear nucleus. PNS Chapter 30

49
Auditory System: Central Pathways
Nagyon komplex Hanginger kialakulása-megszűnése Hangmintázat analízisből tartalmi információ PNS Fig 30-12

50
Auditory System: Central Pathways
Már az első szinapszistól párhuzamos csatornák/ felszállópályák Növekvő komplexitás Különböző sejttípusok különböző projekció és tüzelésimintázat Reciprok összeköttetések Valamennyi állomáson érvényesül a tonotópia-izofrekvenciás lemezek Binauralis interakciók

51
Cochlear Nucleus: VIII nerve: branches -> 3 cochlear nuclei.
Dorsal Cochlear Nucleus (DCN) Posteroventral Cochlear Nucleus (PVCN) Anteroventral Cochlear Nucleus (AVCN) Tonotopy (through innervation order) PNS Fig 30-13 Start of true auditory feature processing. Distinct cell classes: stellate (encode frequency), bushy (encodes sound onset) Different cell types project to different relay nuclei. PNS Fig 30-14
 Posteroventral Cochlear Nucleus (PVCN) Anteroventral Cochlear Nucleus (AVCN) Tonotopy (through innervation order) PNS Fig Start of true auditory feature processing. Distinct cell classes: stellate (encode frequency), bushy (encodes sound onset) Different cell types project to different relay nuclei. PNS Fig")
52
Superior Olive: hangforrás térbeli elhelyezkedésének felismerése
Medial Superior Olive: interaural time differences: Szűk frekvenciatartományra érzékenyek Időbeli különbségnek megfelelően adnak le impulzusokat Jellemző késés PNS Fig 30-15

53
Superior Olive: hangforrás térbeli elhelyezkedésének felismerése
Bilaterális információ Lateral Superior Olive, corpus trapezoideum: interaural intensity differences. A laterális olivamag a két cochleát érő, azonos frekvenciájú hangok intenzitásbeli különbségére érzékeny Aktiváció: ha az azonos oldali intenzitás nagyobb, mint az ellenoldali (corpus trapezoideumból jövő neuronok gátlók) Az oliva complex sérülése megszünteti a hangforrás lokalizációt Principles of Neural Science, Chapter 30
 Az oliva complex sérülése megszünteti a hangforrás lokalizációt. Principles of Neural Science, Chapter 30.")
54
Interaural intensity differences (High Frequency Sounds)
Intensity reaching contralateral ear is lower because of masking by head. Sound Wave Right Ear Thickness of arrow represents intensity reaching each ear.

55
Lateral Superior Olive Circuit Diagram

56
The balance between excitation and inhibition determines response
Ipsilateral input from AVCN Response = excitation - inhibition This picture illustrates the inputs to one LSO neuron. LSO neuron If ipsilateral AVCN is responding more than contralateral AVCN (adjusted by MNTB), respond. Contralateral input from MNTB
, respond. Contralateral input from MNTB.")
57
Auditory System: Midbrain
From superior olives via lateral lemniscus to the inferior colliculus (IC). Separate path from DCN. Dorsal IC: auditory, touch Central Nucleus of IC: combines LSO, MSO inputs to 2-D spatial map; passed on to Superior Colliculus to match visual map Medial geniculate body: Principal nucleus: thalamic relay of auditory system. Tonotopic. Other nuclei: multimodal: visual, touch, role in plasticity?
. Separate path from DCN. Dorsal IC: auditory, touch. Central Nucleus of IC: combines LSO, MSO inputs to 2-D spatial map; passed on to Superior Colliculus to match visual map. Medial geniculate body: Principal nucleus: thalamic relay of auditory system. Tonotopic. Other nuclei: multimodal: visual, touch, role in plasticity")
58
Anatomy source : Palmer & Hall, 2002
Right hemisphere Primary & non-primary auditory cortex Sylvian Fissure Medial Temporal Gyrus planum polare (nonprimary AC) Superior Temporal Gyrus Heschl’s gyrus (primary AC) Superior Temporal Sulcus planum temporale (nonprimary AC)
 Superior. Temporal Gyrus. Heschl’s gyrus. (primary AC) Superior Temporal Sulcus. planum temporale. (nonprimary AC)")
59
Auditory Cortex: Complex patterns
Superior temporal gyrus Like other sensory cortex: Input layer: IV, V: back project to MGB. VI: back project to IC Some 15 distinct tonotopic areas PNS Fig 30-12

60
Sound intensity and activation
Loud sounds (90 dB) activated posterior and medial temporal gyrus (red) Soft (70 dB) sounds activated area (yellow) is found most laterally of TTG Medium intensity (82 dB) sounds activated intermediate area (green). (NeuroImage 2002;17: 710)
 activated posterior and medial temporal gyrus (red) Soft (70 dB) sounds activated area (yellow) is found most laterally of TTG. Medium intensity (82 dB) sounds activated intermediate area (green). (NeuroImage 2002;17: 710)")
61
Cross-modal plasticity in congenitally deaf
These PET/MR images show increased neural activity in the superior temporal gyrus in congenitally deaf subjects when they viewed signs or sign-like movements, suggesting that auditory cortical regions may contribute to the processing of visual information in the deaf

62
... and in congenitally blind

63
Multisensory DSC neurons exhibit the properties of multisensory enhancement and inverse effectiveness Stein and Meredith (1993) The Merging of the Senses, fig
 The Merging of the Senses, fig")
64
Characteristics of multisensory convergence: facilitation (1)
Enhancement of the multimodal neuron’s response with more than one modality

65
Characteristics of multisensory convergence : inhibition (1)
With two modalities, the multisensory neuron’s response is attenuated compared with the response of one modality Excitatory – inhibitory multisensory convergence

66
the peripheral system=the cochlea
Most important to you since this is where damage results in irreversible hearing loss studied in chick show that may not be irreversible! drugs can injure as well as loud sounds Aspirin salicylates quinine (tonic water) streptomycin frequent ear infections
 streptomycin. frequent ear infections.")
67
In this transmission electron micrograph, the tallest stereocilium of an OHC is just detached (by the fixation procedure) from the tectorial membrane. scale bar: 1 µm

68
Acoustic trauma and hair cells 1
Depending on the level of exposure, damage to hair cells may: 1) only involve stereocilia, and some repair mechanism are possible, or 2) involve the entire hair cell, which undergoes apoptosis and dies. Mild damage, which is restricted to stereocilia : ranging from disarray, broken tip links and broken roots to fused and giant stereocilia; in such a case, electromechanical transduction is altered, or stopped, but slow repair may occur.
 only involve stereocilia, and some repair mechanism are possible, or 2) involve the entire hair cell, which undergoes apoptosis and dies. Mild damage, which is restricted to stereocilia : ranging from disarray, broken tip links and broken roots to fused and giant stereocilia; in such a case, electromechanical transduction is altered, or stopped, but slow repair may occur.")
69
Acoustic trauma and hair cells 2
2) Severe and definitive damage, when the hair cell itself is altered and disappears. On the left, TEM of a dying OHC. The flat preparation on the right shows a traumatised cochlea where 7 to 8 OHC have disappeared, together with a pillar cell (damaged area outlined in blue). scale bar: 20 µm.
 Severe and definitive damage, when the hair cell itself is altered and disappears. On the left, TEM of a dying OHC. The flat preparation on the right shows a traumatised cochlea where 7 to 8 OHC have disappeared, together with a pillar cell (damaged area outlined in blue). scale bar: 20 µm.")
Hasonló előadás






 -- sàvszűrő ~àllandó Q-val ? inhibició, komodulalt takaras leengedese (CMR) -- mindez.>")













