Előadást letölteni
Az előadás letöltése folymat van. Kérjük, várjon

1
a tumor-specifikus immunitás kiváltása
Tumor immunoterápia: a tumor-specifikus immunitás kiváltása

2
TUMOR Tumor (neoplázia): növekedési kontrol (immunosurveillance) alól elszabadult sejtek csoportja Benignus (jóindulatú) korlátozott növekedés Malignus (rosszindulatú) korlátlan növekedés (rák) Metastasis: áttét Carcinoma: embrionális ektoderma, vagy endoderma eredetű Sarcoma: mezodermális kötőszövet (szövet, csont) „ Leukémia: hematopoietikus sejt eredetű Limfóma: csontvelői hematopoetikus sejtekből A sejtek malignus transzformációja: Kémiai karcinogén anyagok Besugárzás mutáció , transzformáció Vírusok
 korlátozott növekedés. Malignus (rosszindulatú) korlátlan növekedés (rák) Metastasis: áttét. Carcinoma: embrionális ektoderma, vagy endoderma eredetű. Sarcoma: mezodermális kötőszövet (szövet, csont) „ Leukémia: hematopoietikus sejt eredetű. Limfóma: csontvelői hematopoetikus sejtekből. A sejtek malignus transzformációja: Kémiai karcinogén anyagok. Besugárzás mutáció , transzformáció. Vírusok.")
3
Tumorsejtek immun-átszerkesztése és szelekciója
Cancer immunoediting encompasses three process. (a) Elimination corresponds to immunosurveillance. (b) Equilibrium represents the process by which the immune system iteratively selects and/or promotes the generation of tumor cell variants with increasing capacities to survive immune attack. (c) Escape is the process wherein the immunologically sculpted tumor expands in an uncontrolled manner in the immunocompetent host. In a and b, developing tumor cells (blue), tumor cell variants (red) and underlying stroma and nontransformed cells (gray) are shown; in c, additional tumor variants (orange) that have formed as a result of the equilibrium process are shown. Different lymphocyte populations are as marked. The small orange circles represent cytokines and the white flashes represent cytotoxic activity of lymphocytes against tumor cells. fejlődő tumor sejtek túlélő tumor sejt variánsok Nem transzformált sejtek Ellenőrzés alól elszabadult osztódó tumorsejtek, újabb variánsok
 Elimination corresponds to immunosurveillance. (b) Equilibrium represents the process by which the immune system iteratively selects and/or promotes the generation of tumor cell variants with increasing capacities to survive immune attack. (c) Escape is the process wherein the immunologically sculpted tumor expands in an uncontrolled manner in the immunocompetent host. In a and b, developing tumor cells (blue), tumor cell variants (red) and underlying stroma and nontransformed cells (gray) are shown; in c, additional tumor variants (orange) that have formed as a result of the equilibrium process are shown. Different lymphocyte populations are as marked. The small orange circles represent cytokines and the white flashes represent cytotoxic activity of lymphocytes against tumor cells. fejlődő tumor sejtek. túlélő tumor sejt variánsok. Nem transzformált sejtek. Ellenőrzés alól elszabadult osztódó tumorsejtek, újabb variánsok.")
4
NKT sejtek NK1.1+, abTCR+ sejtek, CD1d-függő, MHC-tól független
korlátozott repertoár: invariáns alfa lánc, béta lánc:Vb8.2,Vb2,Vb7; Vb11 (human) Glycolipidek, lipidek (saját és mikrobiális) hidrofil fejének felismerése (a-glycosphingolipid) a – galactosyl ceramide: IL-4, IL-10, IL-13, IFNg –t termel –> regulator funkció Tumor sejtek pusztítása in vitro Figure 1. Structure of the synthetic α-galactosylceramide.Sphingosine long chain base is shown in green, acyl chain in blue and galactose in red. Galactose is in alpha configuration (see also figure 3). Carbons 1 to 4 on sphingosine are indicated with numbers. In the acyl chain, the alpha carbon position immediately following the carboxyl group is indicated.
 Glycolipidek, lipidek (saját és mikrobiális) hidrofil fejének felismerése (a-glycosphingolipid) a – galactosyl ceramide: IL-4, IL-10, IL-13, IFNg –t termel –> regulator funkció. Tumor sejtek pusztítása in vitro. Figure 1. Structure of the synthetic α-galactosylceramide.Sphingosine long chain base is shown in green, acyl chain in blue and galactose in red. Galactose is in alpha configuration (see also figure 3). Carbons 1 to 4 on sphingosine are indicated with numbers. In the acyl chain, the alpha carbon position immediately following the carboxyl group is indicated.")
5
NKT sejtek által közvetített tumor ellenes hatás modellje
Models for NKT-cell-mediated tumor rejection. NKT cells have been implicated in at least three different experimental models of tumor rejection. The diagrams are designed to demonstrate our current understanding of the effector cells and mechanisms involved in these models, based on in vivo studies. This information may be limited by the restricted number of experimental tumors that have been investigated in detail. Macrophages (MØ) and CD8+ T cells are sometimes shown in white, indicating their unconfirmed role as possible effectors. In these models, IFN-γ may act directly to inhibit tumor growth, tumor-associated angiogenesis or the activation of other effector cell types. It is also likely that other, as-yet unknown, NKT-cell derived factors may also be important in these models. (a) In IL-12-mediated rejection, exogenously administered IL-12 (bottom-left of the figure) can act on effector cells including NKT and NK cells to mediate tumor rejection in vivo. IL-12 induces IFN-γ production by NKT and NK cells, and IFN-γ is a key factor in IL-12-induced, NKT-cell-dependent tumor rejection. Evidence exists for both perforin (Pfp)-dependent and IFN-γ-dependent effector mechanisms in IL-12-mediated tumor rejection, and the relative roles of the effector cells and associated molecules are likely to vary in a tumor- and dose-dependent fashion. (b) In α-GalCer-mediated rejection, exogenously administered α-GalCer is presented by CD1d and can induce potent, CD1ddependent,NKT-cell-dependent tumor rejection in vivo. This process is IL-12- and IFN-γ-dependent, and is perforin-independent (indep.). The importance of CD8+ T cells may be model-dependent. (c) In natural rejection, NKT cells can also mediate tumor rejection in the absence of exogenous stimulation via endogenous glycolipid antigen (endog. glycolipid Ag), indicating that tumor immunosurveillance is also a natural, or physiological, function of these cells. Although few examples currently exist, it is known to be IL-12- and IFN-γ-dependent, similar to the α-GalCer induction in (b). In contrast to the previous α-GalCer model, the natural role of NKT cells is perforin-dependent and also depends on CD8+ T-cell recognition of tumor peptides (top-right of the figure). Of potential importance is the dependence on CD1d, suggesting that NKT cells may be capable of responding to tumor-derived glycolipid antigens. a-gal-cer anti-metastatikus hatása: indirekt hatás NK sejtekre, - T sejtek ? B sejtek? ? FasL-, TRAIL -, TNF- hiányos egérben, anti-IL4 mellett nincs hatás. Prolifeláció és vascularizáció gátlása, tumor hypoxia
 and CD8+ T cells are sometimes shown in white, indicating their unconfirmed role as possible effectors. In these models, IFN-γ may act directly to inhibit tumor growth, tumor-associated angiogenesis or the activation of other effector cell types. It is also likely that other, as-yet unknown, NKT-cell derived factors may also be important in these models. (a) In IL-12-mediated rejection, exogenously administered IL-12 (bottom-left of the figure) can act on effector cells including NKT and NK cells to mediate tumor rejection in vivo. IL-12 induces IFN-γ production by NKT and NK cells, and IFN-γ is a key factor in IL-12-induced, NKT-cell-dependent tumor rejection. Evidence exists for both perforin (Pfp)-dependent and IFN-γ-dependent effector mechanisms in IL-12-mediated tumor rejection, and the relative roles of the effector cells and associated molecules are likely to vary in a tumor- and dose-dependent fashion. (b) In α-GalCer-mediated rejection, exogenously administered α-GalCer is presented by CD1d and can induce potent, CD1ddependent,NKT-cell-dependent tumor rejection in vivo. This process is IL-12- and IFN-γ-dependent, and is perforin-independent (indep.). The importance of CD8+ T cells may be model-dependent. (c) In natural rejection, NKT cells can also mediate tumor rejection in the absence of exogenous stimulation via endogenous glycolipid antigen (endog. glycolipid Ag), indicating that tumor immunosurveillance is also a. natural, or physiological, function of these cells. Although few examples currently exist, it is known to be IL-12- and IFN-γ-dependent, similar to the α-GalCer induction in (b). In contrast to the previous α-GalCer model, the natural role of NKT cells is perforin-dependent. and also depends on CD8+ T-cell recognition of tumor peptides (top-right of the figure). Of potential importance is the dependence on. CD1d, suggesting that NKT cells may be capable of responding to tumor-derived glycolipid antigens. a-gal-cer anti-metastatikus hatása: indirekt hatás NK sejtekre, - T sejtek B sejtek FasL-, TRAIL -, TNF- hiányos egérben, anti-IL4 mellett nincs hatás. Prolifeláció és vascularizáció gátlása, tumor hypoxia.")
6
A tumor immunsejtek által történő elpusztításának modelje
. A proposed model for the elimination phase of the cancer immunoediting process. (a) The initiation of the response in which lymphocytes that participate in innate immunity (NKT, NK and T cells) recognize transformed cells that have accumulated above a threshold that has yet to be defined and are stimulated to produce IFN- . (b) The initial IFN- starts a cascade of innate immune reactions that involve (i) the induction of chemokines, including the angiostatic chemokines (CXCL10 (1P10), CXCL9 (MIG) AND CXCL11 (I-TAC)) that block neovascularization in the tumor and that also effect the recruitment of NK cells, dendritic cells, macrophages and other immune effector cells to the tumor site; (ii) an antiproliferative action of IFN- on the developing tumor and (iii) the activation of cytocidal activity in macrophages and NK cells entering the tumor. These events result in some tumor cell death by both immunologic and nonimmunologic mechanisms. Dead tumor cells or tumor cell debris (blue squares) are ingested by dendritic cells and are trafficked to the draining lymph node. (c) Tumor growth is kept in check by the cytocidal activities of NK cells and activated macrophages while CD4+ and CD8+ T cells that are specific for tumor antigens develop in the draining lymph node. (d) Tumor-specific CD4+ and CD8+ T cells home to the tumor along a chemokine gradient where they recognize and destroy tumor cells expressing distinctive tumor antigens. Tumor cells (blue); nontransformed cells (gray); dead tumor cells (white to gray gradient circles surrounded by a dashed black line); lymphocytes, dendritic cells (DC) and macrophages (Mac) are marked and colored appropriately.
 The initiation of the response in which lymphocytes that participate in innate immunity (NKT, NK and T cells) recognize transformed cells that have accumulated above a threshold that has yet to be defined and are stimulated to produce IFN- . (b) The initial IFN- starts a cascade of innate immune reactions that involve (i) the induction of chemokines, including the angiostatic chemokines (CXCL10 (1P10), CXCL9 (MIG) AND CXCL11 (I-TAC)) that block neovascularization in the tumor and that also effect the recruitment of NK cells, dendritic cells, macrophages and other immune effector cells to the tumor site; (ii) an antiproliferative action of IFN- on the developing tumor and (iii) the activation of cytocidal activity in macrophages and NK cells entering the tumor. These events result in some tumor cell death by both immunologic and nonimmunologic mechanisms. Dead tumor cells or tumor cell debris (blue squares) are ingested by dendritic cells and are trafficked to the draining lymph node. (c) Tumor growth is kept in check by the cytocidal activities of NK cells and activated macrophages while CD4+ and CD8+ T cells that are specific for tumor antigens develop in the draining lymph node. (d) Tumor-specific CD4+ and CD8+ T cells home to the tumor along a chemokine gradient where they recognize and destroy tumor cells expressing distinctive tumor antigens. Tumor cells (blue); nontransformed cells (gray); dead tumor cells (white to gray gradient circles surrounded by a dashed black line); lymphocytes, dendritic cells (DC) and macrophages (Mac) are marked and colored appropriately.")
7
Replikációs életszakasz Gén expresszió változás
Az önfenntartó sejtszaporodáshoz vezető molekuláris útvonalak Sejtciklus Replikációs életszakasz Gén expresszió változás Apoptózis Cancer arises from the stepwise accumulation of genetic changes that confer upon an incipient neoplastic cell the properties of unlimited, self-sufficient growth and resistance to normal homeostatic regulatory mechanisms. Advances in human genetics and molecular and cellular biology have identified a collection of cell phenotypes — the main destinations in the subway map below — that are required for malignant transformation1. Specific molecular pathways (subway lines) are responsible for programming these behaviours. Although the connections between cancer-cell wiring and function remain incompletely explored and specified — hence the many lines under construction — the broad outlines of the molecular circuitry of the cancer cell can now be sketched. Further advances in understanding these pathways and their interconnections will accelerate the development of molecularly targeted therapies that promise to change the practice of oncology. Anyagcsere
 are responsible for programming these behaviours. Although the connections between cancer-cell wiring and function remain incompletely explored and specified — hence the many lines under construction — the broad outlines of the molecular circuitry of the cancer cell can now be sketched. Further advances in understanding these pathways and their interconnections will accelerate the development of molecularly targeted therapies that promise to change the practice of oncology. Anyagcsere.")
8
Sejt ciklus: tumor szuppresszor fehérjék ellenőrzése alatt áll
G1 fázisban külső+belső jelek alapján döntenek a nyugvó állapot vagy a sejtciklusba lépés között További ellenőrzési pontok későbbi fázisban Tumor szuppressziós útvonalak gátoltak retonoblastoma (RB) út: E2F transzkripciós faktorok által ellenőrzött gének(sejtciklus) expresszióját gátolja, foszforilációtól függően inaktiválódik, cyclin dependens kinázok szabályozzák P53 út: apoptózist aktivál, gátolja a sejtciklusba lépést (CDK inhibitort aktivál) Normális sejt: RAS út tartós aktiválása aktiválja az RB és p53 utat-> befagyasztja a sejtciklust - védekező mechanizmus TGFb út: ser/thr kinázok aktiválása -> SMAD transzkripciós faktorok aktiválása sejtciklust szabályozó gének (CDK gátlók, Myc onkogén).
 út: E2F transzkripciós faktorok által ellenőrzött gének(sejtciklus) expresszióját gátolja, foszforilációtól függően inaktiválódik, cyclin dependens kinázok szabályozzák. P53 út: apoptózist aktivál, gátolja a sejtciklusba lépést (CDK inhibitort aktivál) Normális sejt: RAS út tartós aktiválása aktiválja az RB és p53 utat-> befagyasztja a sejtciklust - védekező mechanizmus. TGFb út: ser/thr kinázok aktiválása -> SMAD transzkripciós faktorok aktiválása sejtciklust szabályozó gének (CDK gátlók, Myc onkogén).")
9
Tumor kialakulásában szerepet játszó gének:
tumor gének 291 tumor gént azonosítottak, ez több mint 1 %-a a teljes genomnak Ezek 90 %-a szomatikus mutáción esett át, 20 % embrionális mutációt, 10 % mindkettőt tartalmaz Kromoszóma transzlokáció a leggyakoribb – kiméra gén A legtöbb tumor gént azonosították leukémia, limfóma és szarkóma esetében. Ezek csak 10 %-át adják az összes tumoros betegségeknek A legközönségesebb domén, amely a tumor gének által kódolt: protein kináz 2004-es felmérés

10
Kromoszóma transzlokáció Burkitt limfómában
EBV fertőzés: c-myc az áthelyeződik az Ig H lánc enhancer régiójához – B sejtek korlátlan növekedése

11
A tumorspecifikus antigén kialakulása
karcinogén „privát antigének”

12
a sejttranszformációt kiváltó fehérjét kódoló gén
Onkogén: a sejttranszformációt kiváltó fehérjét kódoló gén 1910: Rous sarcoma virus: v-src (c-src) 1966 Nobel díj (csirke szarkóma sejtek szűrlete RNS vírust tartalmaz) Normális sejtekben: proto-onkogének, a sejtproliferációt, sejtciklust, túlélést, illetve apoptózist szabályozó fehérjék Mutáció, transzlokáció következtében megváltozik: aktivitásuk, mennyiségük, funkciójuk
 1966 Nobel díj. (csirke szarkóma sejtek szűrlete RNS vírust tartalmaz) Normális sejtekben: proto-onkogének, a sejtproliferációt, sejtciklust, túlélést, illetve apoptózist szabályozó fehérjék. Mutáció, transzlokáció következtében megváltozik: aktivitásuk, mennyiségük, funkciójuk.")
13
A tumor és az immunrendszer

14
Tumor „megszökése” az immunrendszer elől:
· alacsony MHCI expresszió, csökkent prezentáció · nincs MHCII expresszió - TH sejteket nem aktivál · kostimulátor molekulák (B7) hiánya · tumorsejt terméke elnyomja immunválaszt (TGF, IL-10) · a gazdaszervezet toleranciája · mutáns tumor sejtek elszaporodása, amelyek nem expresszálnak peptid- MHCI komplexet · antigén moduláció: tumor antigének elveszitése, vagy elrejtése ellenanyagok által · rezisztens tumor kialakulása · tumor antigének elfedése sziálsav tartalmu mukopoliszaharidok által - „antigen masking”
 hiánya. · tumorsejt terméke elnyomja immunválaszt (TGF, IL-10) · a gazdaszervezet toleranciája. · mutáns tumor sejtek elszaporodása, amelyek nem expresszálnak peptid- MHCI komplexet. · antigén moduláció: tumor antigének elveszitése, vagy elrejtése ellenanyagok által. · rezisztens tumor kialakulása. · tumor antigének elfedése sziálsav tartalmu mukopoliszaharidok által - „antigen masking")
15
A tumor felismerésének közvetett és közvetlen útjai
Figure 2 | Direct and indirect pathways of cancer recognition. a | Innate immune cells recognize cancers through germ-line-encoded pattern-recognition receptors and other cell-surface molecules. Cancers express various stress-induced genes such as MICA and MICB that trigger NKG2D receptors on natural killer (NK) cells, macrophages and some cytotoxic T lymphocytes (CTLs). NK cells also use a combination of inhibitory and activating receptors such as the killer-cell immunoglobulin-like receptors (KIRs) to detect the loss of major histocompatibility complex (MHC) class I expression on cancers. Dendritic cells (DCs) phagocytose apoptotic tumours through CD36 and v 5. Macrophages and DCs also use scavenger receptors and CD91 to ingest heat-shock proteins (HSPs) complexed with tumour-derived peptides that are released from necrotic cancer cells. b | Adaptive immune cells are initially stimulated to recognize cancer cells through cross-priming by DCs. As tumour cells do not express co-stimulatory molecules that are important for T-cell activation, they generally cannot prime cellular responses efficiently. Instead, DCs capture dying tumour cells or debris, migrate to regional lymph nodes and process the tumour-derived material onto CD1D, for presentation to NKT cells (not shown), and MHC class I and class II molecules for presentation to CD4+ and CD8+ T cells. Mature DCs express co-stimulatory molecules such as B7-1 and B7-2 to enhance T-cell activation. DCs secrete IL-12 and IL-18 to promote T helper 1 (TH1) CD4+ T-cell responses and cytotoxic CD8+ T cells. Activated CD4+ T cells and NKT cells express CD40 ligand, which further stimulates DC maturation through CD40 signalling. CD4+ T cells and DCs also help trigger B cells to produce antibodies that are reactive with tumour proteins.
 cells, macrophages and some cytotoxic T lymphocytes (CTLs). NK cells also use a combination of inhibitory and activating receptors such as the killer-cell immunoglobulin-like receptors (KIRs) to detect the loss of major histocompatibility complex (MHC) class I expression on cancers. Dendritic cells (DCs) phagocytose apoptotic tumours through CD36 and v 5. Macrophages and DCs also use scavenger receptors and CD91 to ingest heat-shock proteins (HSPs) complexed with tumour-derived peptides that are released from necrotic cancer cells. b | Adaptive immune cells are initially stimulated to recognize cancer cells through cross-priming by DCs. As tumour cells do not express co-stimulatory molecules that are important for T-cell activation, they generally cannot prime cellular responses efficiently. Instead, DCs capture dying tumour cells or debris, migrate to regional lymph nodes and process the tumour-derived material onto CD1D, for presentation to NKT cells (not shown), and MHC class I and class II molecules for presentation to CD4+ and CD8+ T cells. Mature DCs express co-stimulatory molecules such as B7-1 and B7-2 to enhance T-cell activation. DCs secrete IL-12 and IL-18 to promote T helper 1 (TH1) CD4+ T-cell responses and cytotoxic CD8+ T cells. Activated CD4+ T cells and NKT cells express CD40 ligand, which further stimulates DC maturation through CD40 signalling. CD4+ T cells and DCs also help trigger B cells to produce antibodies that are reactive with tumour proteins.")
16
Koordinált sejtes és humorális reakciók közvetítik a tumor sejtek pusztulását
Figure 3 | A coordinated cellular and humoral reaction mediates tumour destruction. Following stimulation, natural killer (NK) cells can lyse tumours through the perforin/granzyme pathway or apoptosis-inducing ligands such as tumour-necrosis factor (TNF)-related apoptosis-inducing ligand (TRAIL). NK cells secrete interferon- (IFN- ), which inhibits tumour-cell proliferation, enhances tumour-cell apoptosis, improves tumour antigen presentation and inhibits angiogenesis. NKT cells also execute cytotoxicity and cytokine production. Cytotoxic CD8+ T cells lyse tumours through death ligands, such as TRAIL, and the perforin/granzyme pathway. CD4+ T cells can differentiate into T helper 1 (TH1) cells that secrete IFN- and TNF- or T helper 2 (TH2) cells that secrete interleukin (IL)-4, IL-5, IL-6, IL-10 and IL-13. The latter cytokines enhance eosinophil function and increase antibody production by B cells. Antibodies to cancer cell-surface molecules can inhibit oncogenic signalling and/or stimulate tumour destruction through engaging Fc receptors on macrophages, granulocytes and NK cells (not shown). Antibodies can further promote tumour antigen presentation by dendritic cells (DCs) through immune-complex formation. Macrophages can lyse tumours through the production of nitric oxide and reactive oxygen species. Tumour blood vessels can also be attacked by lymphocytes and granulocytes.
 cells can lyse tumours through the perforin/granzyme pathway or apoptosis-inducing ligands such as tumour-necrosis factor (TNF)-related apoptosis-inducing ligand (TRAIL). NK cells secrete interferon- (IFN- ), which inhibits tumour-cell proliferation, enhances tumour-cell apoptosis, improves tumour antigen presentation and inhibits angiogenesis. NKT cells also execute cytotoxicity and cytokine production. Cytotoxic CD8+ T cells lyse tumours through death ligands, such as TRAIL, and the perforin/granzyme pathway. CD4+ T cells can differentiate into T helper 1 (TH1) cells that secrete IFN- and TNF- or T helper 2 (TH2) cells that secrete interleukin (IL)-4, IL-5, IL-6, IL-10 and IL-13. The latter cytokines enhance eosinophil function and increase antibody production by B cells. Antibodies to cancer cell-surface molecules can inhibit oncogenic signalling and/or stimulate tumour destruction through engaging Fc receptors on macrophages, granulocytes and NK cells (not shown). Antibodies can further promote tumour antigen presentation by dendritic cells (DCs) through immune-complex formation. Macrophages can lyse tumours through the production of nitric oxide and reactive oxygen species. Tumour blood vessels can also be attacked by lymphocytes and granulocytes.")
17
Tumor specifikus memória T sejtek szaporodása
GM-CSF szekretáló tumor sejt vakcina + anti-CTLA4 ellenanyag blokád szinergizáló anti-tumor hatást mutat Figure 4 | GM-CSF-secreting tumour-cell vaccines and CTLA-4 antibody blockade show synergistic antitumour effects. Vaccination with irradiated tumour cells that are engineered to secrete granulocyte–marcrophage colony-stimulating factor (GM-CSF) stimulates infiltration of dendritic cells (DCs), macrophages and granulocytes at the immunization site. This coordinated cellular reaction promotes the efficient phagocytosis of tumour debris by DCs. G further induces DCs to mature and migrate to regional lymph nodes to prime tumour-specific T and B cells. Activated T cells then upregulate surface cytotoxic-T-lymphocyte-associated protein 4 (CTLA-4), which, following engagement by B7 molecules, signals to limit T-cell expansion and cytokine production. CTLA-4 antibody blockade attenuates this inhibition and thereby augments T-cell proliferation and effector functions. CTLA-4 antibody blockade can also inhibit regulatory T cells (which constitutively express surface CTLA-4) and thereby promote the expansion of tumour-specific memory T cells. Tumor specifikus memória T sejtek szaporodása
 stimulates infiltration of dendritic cells (DCs), macrophages and granulocytes at the immunization site. This coordinated cellular reaction promotes the efficient phagocytosis of tumour debris by DCs. G further induces DCs to mature and migrate to regional lymph nodes to prime tumour-specific T and B cells. Activated T cells then upregulate surface cytotoxic-T-lymphocyte-associated protein 4 (CTLA-4), which, following engagement by B7 molecules, signals to limit T-cell expansion and cytokine production. CTLA-4 antibody blockade attenuates this inhibition and thereby augments T-cell proliferation and effector functions. CTLA-4 antibody blockade can also inhibit regulatory T cells (which constitutively express surface CTLA-4) and thereby promote the expansion of tumour-specific memory T cells. Tumor specifikus memória T sejtek szaporodása.")
18
A tumorsejtek transzfektálása B7 –tel, vagy GM-CSF –el növeli a tumor immunogenitását
Transfection of tumors with the gene for B7 or for GM-CSF enhances tumor immunogenicity. A tumor that does not express co-stimulatory molecules will not induce an immune response, even though it might express tumor rejection antigens (TRAs), because naive CD8 T cells specific for the TRA cannot be activated by the tumor. The tumor therefore grows progressively in normal mice and eventually kills the host (top panels). If such tumor cells are transfected with a co-stimulatory molecule, such as B7, TRA-specific CD8 T cells now receive both signal 1 and signal 2 from the same cell (see Section 8-5) and can therefore be activated (center panels). The same effect can be obtained by transfecting the tumor with the gene encoding GM-CSF, which attracts and stimulates the differentiation of dendritic cell precursors (bottom panels). Both these strategies have been tested in mice and shown to elicit memory T cells, although results with GM-CSF are more impressive. Because TRA-specific CD8 cells have now been activated, even the original B7-negative or GM-CSF negative tumor cells can be rejected.
, because naive CD8 T cells specific for the TRA cannot be activated by the tumor. The tumor therefore grows progressively in normal mice and eventually kills the host (top panels). If such tumor cells are transfected with a co-stimulatory molecule, such as B7, TRA-specific CD8 T cells now receive both signal 1 and signal 2 from the same cell (see Section 8-5) and can therefore be activated (center panels). The same effect can be obtained by transfecting the tumor with the gene encoding GM-CSF, which attracts and stimulates the differentiation of dendritic cell precursors (bottom panels). Both these strategies have been tested in mice and shown to elicit memory T cells, although results with GM-CSF are more impressive. Because TRA-specific CD8 cells have now been activated, even the original B7-negative or GM-CSF negative tumor cells can be rejected.")
19
A tumor antigéneket felismerheti mind a természetes, mind a szerzett immunrendszer.
A gazdaszervezet citokin termelése elnyomhatja a tumor képződést az infekció, gyulladás és az immunválasz szabályozása révén A tumor sejtek felhasználhatják a gazda citokinjeit a szaporodás elősegítésére, az apoptózistól való megmenekülésre és a szétterjedésre Citokinek szisztémás alkalmazása anti-tumor hatású, de súlyos toxikus mellékhatásokkal jár, mint pl. infekció, ez behatárolja az alkalmazást. Tumor sejteket genetikai manipulációval módosítani lehet immunstimuláló citokinek termelésére, ezek tumor vakcinák alapjait képezhetik CTLA-4 blokkoló receptor gátlása ellenanyaggal ígéretes stratégia a tumor vakcina hatékonyságának növelésére, de rizikó faktor –saját antigénekkel szembeni toleranciát is elnyomhatja.

20
A tumor és az immunrendszer egymásrahatása - a tumor ellenes immunválasz kialakulása
CD8+ Tc-sejt Tumor-specifikus CTL indukció Citokinek Tumor antigének szeparálása Dendritikus sejtek feltöltése Tumor-specifikus CTL Genetikai módosítás (MHCII expresszió)
")
22
CÉL: a gazdaszervezet tumorspecifikus immunválaszának fokozása
IMMUNTERÁPIA CÉL: a gazdaszervezet tumorspecifikus immunválaszának fokozása Tumor antigének: Tumor sejtekben a mutációk gyakorisága nagy => fehérje expresszió a normálistól eltér Oncogén virusok által indukált tumor: vírus fehérjék expressziója (CTL aktiválás) Abnormális szerkezetű lipidek, szénhidrátok szintézise (B sejtek aktiválása) Tumor specifikus antigének -normális sejten nincs Tumor asszociált antigének - normális sejten is van -tolerogén
 Abnormális szerkezetű lipidek, szénhidrátok szintézise (B sejtek aktiválása) Tumor specifikus antigének -normális sejten nincs. Tumor asszociált antigének - normális sejten is van -tolerogén.")
23
Stratégiák a kísérletes tumor immunterápiára
Fig. 1. Current strategies in experimental immunotherapy of tumors. Tumor-specific mAbs can mediate cytolysis either by engaging NK cells via Fc receptors (ADCC), or by complement activation. To reduce immunogenicity of xenogeneic antibodies, constant regions are humanized by recombinant technology. Bispecific antibody constructs are designed to bring immune effector cells into contact with tumor cells and to simultaneously stimulate their cytotoxic activity. Examples include antibodies that recognize a tumor surface antigen and CD16 to activate NK cells, or CD3 to activate T cells. Recombinant fusion products of antitumor antibodies and cytokines can concentrate immune effector functions at the tumor site. CTLs can be activated against tumor antigens by tumor cells rendered immunogenic by expression of either costimulatory molecules such as CD80/86 or cytokines. Effective stimulation of tumor-specific CTLs is achieved by dendritic cells (DCs) presenting major histocompatibility complex class I-bound tumor peptides. DCs are either directly loaded with peptides or exposed to tumor-cell lysates, tumor proteins or DNA. Intracellular synthesis and processing of tumor proteins in DCs is achieved by transfection of cDNA in an expression vector. Abbreviations: ADCC, antibody-mediated cellular cytotoxicity; CTL, cytotoxic T lymphocyte; IL-2, interleukin 2; mAb, monoclonal antibody; NK, natural killer.
, or by complement activation. To reduce immunogenicity of xenogeneic antibodies, constant regions are humanized by recombinant technology. Bispecific antibody constructs are designed to bring immune effector cells into contact with tumor cells and to simultaneously stimulate their cytotoxic activity. Examples include antibodies that recognize a tumor surface antigen and CD16 to activate NK cells, or CD3 to activate T cells. Recombinant fusion products of antitumor antibodies and cytokines can concentrate immune effector functions at the tumor site. CTLs can be activated against tumor antigens by tumor cells rendered immunogenic by expression of either costimulatory molecules such as CD80/86 or cytokines. Effective stimulation of tumor-specific CTLs is achieved by dendritic cells (DCs) presenting major histocompatibility complex class I-bound tumor peptides. DCs are either directly loaded with peptides or exposed to tumor-cell lysates, tumor proteins or DNA. Intracellular synthesis and processing of tumor proteins in DCs is achieved by transfection of cDNA in an expression vector. Abbreviations: ADCC, antibody-mediated cellular cytotoxicity; CTL, cytotoxic T lymphocyte; IL-2, interleukin 2; mAb, monoclonal antibody; NK, natural killer.")
24
TUMOR ANTIGÉNEK Tumor ellen gazdaszervezet immunválasszal reagál: limfocita függő Specifikus tumor ellenes ellenanyagok memória van „Immunsurveilance” - tumor elpusztítására tumor specifikus antigének (TSA) - csak az adott tumorra jellemző MHC-hez kötődött peptidek tumorhoz asszociált antigének (TAA) - embrionális gének termékei- mennyiségi növekedés 50x- 100x Tumor antigének izolálása: cDNS könyvtárból - CTL segítségével, tumor sejtek felszínéről savas elucióval
 - csak az adott tumorra jellemző. MHC-hez kötődött peptidek. tumorhoz asszociált antigének (TAA) - embrionális gének termékei- mennyiségi növekedés 50x- 100x. Tumor antigének izolálása: cDNS könyvtárból - CTL segítségével, tumor sejtek felszínéről savas elucióval.")
25
Citotoxikus T sejt klónok előállítása:
in vitro: melanoma sejt tenyészet (tumorból) + limfociták (betegből) Tumor specifikus CTL Tumor cDNS könyvtár transzfektálás MHCI+ célsejtekbe : Tumor antigént bemutató sejtek Együtt tenyésztés, amelyik sejt elpusztult az tartalmazta a megfelelő tumor peptidet Izolálás, szekvenálás tumor ag.
 + limfociták (betegből) Tumor specifikus CTL. Tumor cDNS könyvtár transzfektálás MHCI+ célsejtekbe : Tumor antigént bemutató sejtek. Együtt tenyésztés, amelyik sejt elpusztult az tartalmazta a megfelelő tumor peptidet. Izolálás, szekvenálás tumor ag.")
26
Természetes módon processzált tumor peptidek előállításának módjai
Peptid bejuttatása a sejtbe: Liposzóma (MHCII) Kiméra baktérium (MHCII) Vírus kiméra –minigen (MHCI) Fig. 1. Methods of purification of naturally processed tumor peptides. The steps for the purification of strong extracts (left) and mild extracts (right) are depicted. For mild extracts, a further step of enrichment of material of molecular weight,5000 can be added after the lyophilization.
 Kiméra baktérium (MHCII) Vírus kiméra –minigen (MHCI) Fig. 1. Methods of purification of naturally processed tumor peptides. The steps for the purification. of strong extracts (left) and mild extracts (right) are depicted. For mild extracts, a further step of. enrichment of material of molecular weight,5000 can be added after the lyophilization.")
27
Tumor elleni vakcináció
A processzálás hiányában nincs CTL felismerés - tumor epitópok (pl. HLA-A2 korlátozott melanoma differenciálódási epitóp) rosszul processzálódnak Anti-tumor válasz kiváltható a peptiddel (tumor epitóp) Impact of the lack of processing of certain tumor epitopes by the immunoproteasome on vaccination efficiency. For epitopes that are poorly processed by the immunoproteasome,such as the HLA-A2-restricted melanoma differentiation epitope Melan-A26–35, CTLresponses should (a) be induced more efficiently with immunization modalities that do not depend on processing by dendritic cells,such as peptides or recombinant viruses encoding only the peptide, than (b) with proteins or recombinant viruses encoding full-length proteins.
 rosszul processzálódnak. Anti-tumor válasz kiváltható a peptiddel (tumor epitóp) Impact of the lack of processing of certain tumor epitopes by the immunoproteasome on vaccination efficiency. For epitopes that are poorly processed by the immunoproteasome,such as the HLA-A2-restricted melanoma differentiation epitope Melan-A26–35, CTLresponses should (a) be induced more efficiently with immunization modalities that do not depend on processing by dendritic cells,such as peptides or recombinant viruses encoding only the peptide, than (b) with proteins or recombinant viruses encoding full-length proteins.")
28
Peptidek terápiás alkalmazása
Szintetikus peptidek Természetes peptidek Előny Immunválasz specifitása sokféle ag keveréke Korlátlan mennyiség több betegnél Nagy tisztaság CD4 CD8 T akt. Kevés tumor sejten ellenőrizhető CD4, CD8 T aktiválás Hátrány Tumorspecifitásra nincs bizonyíték, sok tumor sejt kell Néhány betegnél, alacsony koncentráció Mutáns sejtek megszökhetnek, autoimmunitás kialakulhat Autoimmunitás kialakulhat

29
Ellenanyag terápiák

Hasonló előadás
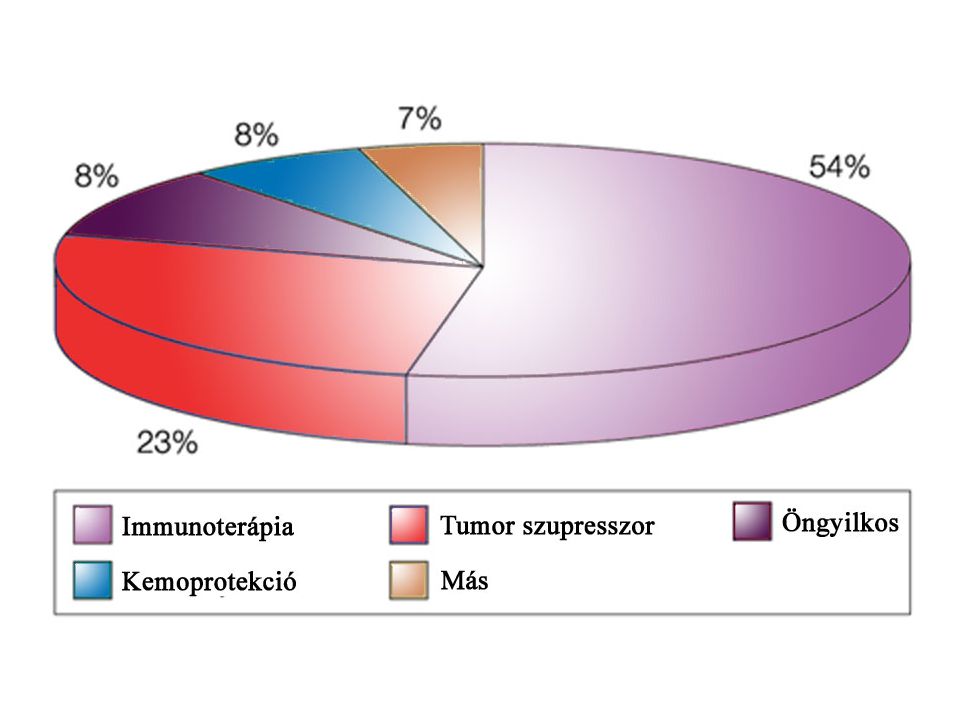

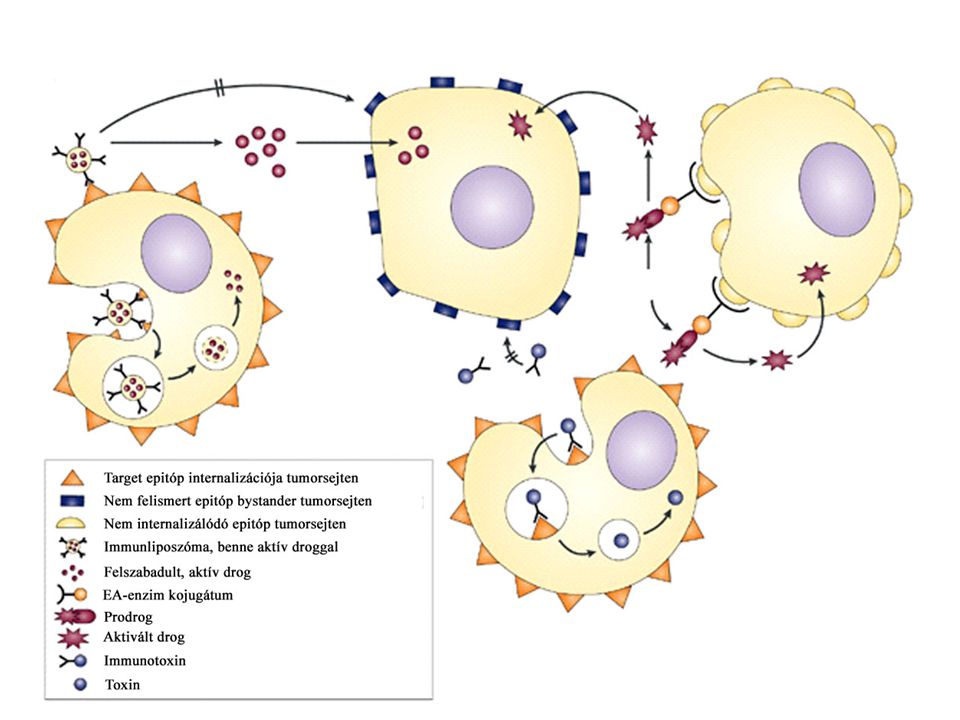
 daganatkeltő.>")














>")
>")


