Előadást letölteni
Az előadás letöltése folymat van. Kérjük, várjon

1
A sejtadhézió a sejtek kapcsolata
OOOOO ÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖÖBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBBB8ÁÁÁÁÁÁÁÁÁÁÁÖR7I87PÖÜ Dr.Darvas Zsuzsanna PhD Genetikai, Sejt- és Immunbiológiai Intézet

2
Specifikus molekula családok
sejt-sejt sejt-extracelluláris mátrix (ECM) Szerep: embrionális fejlődés, differenciálódás, tumorképződés, metasztázis, migráció
 Szerep: embrionális fejlődés, differenciálódás, tumorképződés, metasztázis, migráció.")
3
Adhéziós molekulák jellemzése
Sejt-sejt vagy sejt-mátrix Ion (Ca++, Mg++) függőség Homofil v. heterofil kapcsolódás Együttműködés a sejtváz komponensel Időleges vagy állandósult struktúra része
 függőség. Homofil v. heterofil kapcsolódás. Együttműködés a sejtváz komponensel. Időleges vagy állandósult struktúra része.")
4
Adhéziós molekula családok főbb típusai ill. képviselői
3 kadherinek

5
kadherinek Homofil kötődés (Ca ion függő)
Kapcsoló fehérjék pl. kateninek mikro- vagy intermedier filamentumokhoz Sejtkapcsoló struktúrákban is Szövetspecifitás: N – neural P – placental E - epithelial motivum : Ca ion kötő domének

6
kadherinek:homofil kötődés
Cisz és transz dimer forma

7
Alfa, béta és gamma katenin
Kapcsoló fehérjék: Aktin filamentum Alfa, béta és gamma katenin Sejtvázelem: aktin/mikrofilamentum Ca 2+ függőség: hiányában felbomlik a kapcsolat sandra.kaminik.de/?page_id=3

8
E kadherin szerepe a morula kialakulásában

9
Bakteriális infekció menete:lysteriosis
L. monocytogenes induces its entry into a non-professional phagocyte. b | Bacteria are internalized in a vacuole (also known as a phagosome). c,d | The membrane of the vacuole is disrupted by the secretion of two phospholipases, PlcA and PlcB, and the pore-forming toxin listeriolysin O. Bacteria are released into the cytoplasm, where they multiply and start to polymerize actin, as observed by the presence of the characteristic actin tails . e | Actin polymerization allows bacteria to pass into a neighbouring cell by forming protrusions in the plasma membrane. f | On entry into the neighbouring cell, bacteria are present in a double-membraned vacuole, from which they can escape to perpetuate the cycle. F-actin, filamentous actin. Electron micrographs a–c,e–f are reproduced with permission from Ref. 113 © (1998) European Molecular Biology Organization, and d is reproduced with permission from Ref. 30 © (1992) Elsevier. Endocitózis – szökés a lizoszómából – aktin citoszkeleton használata további sejtek infekciójához
. c,d | The membrane of the vacuole is disrupted by the secretion of two phospholipases, PlcA and PlcB, and the pore-forming toxin listeriolysin O. Bacteria are released into the cytoplasm, where they multiply and start to polymerize actin, as observed by the presence of the characteristic actin tails . e | Actin polymerization allows bacteria to pass into a neighbouring cell by forming protrusions in the plasma membrane. f | On entry into the neighbouring cell, bacteria are present in a double-membraned vacuole, from which they can escape to perpetuate the cycle. F-actin, filamentous actin. Electron micrographs a–c,e–f are reproduced with permission from Ref. 113 © (1998) European Molecular Biology Organization, and d is reproduced with permission from Ref. 30 © (1992) Elsevier. Endocitózis – szökés a lizoszómából – aktin citoszkeleton használata további sejtek infekciójához.")
10
E kadherin szerepe a folyamatban
internalin A molekula (InlA) segítségével a baktérium az E kadherinhez kötődik és ezután lép be endocitózissal
 segítségével a baktérium az E kadherinhez kötődik és ezután lép be endocitózissal.")
11
CAM=cell adhesion molecules
Immunglobulin szerű sejtadhéziós molekulák

12
Új név: Ig-szerű CAM-ok
Ált. homofil kötődés Sejt-sejt Ca++ ion független Kapcsoló fehérjék mikrofilamentumokhoz Sejt/szövet specifikus VCAM1, ICAM, NCAM vascular/endothelial intercellular neuronal NCAM: 20 féle variáns alternativ splicing-al

13
Ig szerű adhéziós molekulák: extracelluláris domén motivumok „zsebek"
Ig domén: antiparalell béta redők diszulfid hidakkal

14
Hisztogenezis folyamatában fontos mechanizmusok
A velőcsőről lefűződő sejtek vándorlása során integrinek és NCAM szerepe

15
NCAM szerepe az embrionális fejlődésben
Neural crest (velőléc): a velőcsőből lefűződő sejtek- ezek a sejtek a vándorlásuk során változtatják az NCAM expressziót
: a velőcsőből lefűződő sejtek- ezek a sejtek a vándorlásuk során változtatják az NCAM expressziót.")
16
Szelektin adhéziós molekulák

17
Sejt/szövet specifikus E – epithelial L – lymphoid P - platelet
Szelektinek Sejt-sejt Heterofil kötődés Ca++ ion függő Sejt/szövet specifikus E – epithelial L – lymphoid P - platelet

18
CRD: calcium-dependent carbohydrate recognition domain
CRD: calcium-dependent carbohydrate recognition domain C lektin Heterofil kötődés a másik sejt plazmamembránjának egyes cukorkomponenseihez

19
P szelektin ligandja: PSGL-1 ( P selectin glycoprotein ligand 1)
bme.virginia.edu/ley/psgl-1.html P-selectin glycoprotein ligand-1 is a 240 kDa homodimer consisting of two 120kDa polypeptide chains. PSGL-1 is constitutively expressed on all leukocytes. PSGL-1 is primarily found on the tips of the microvilli. PSGL-1 can bind to P-selectin on the endothelium when decorated with appropriate sugars. The structure of functional PSGL-1 includes the sialyl-Lewisx component. The PSGL-1 gene encodes a transmembrane polypeptide rich in proline, serine and threonine residues typical of mucin-type glycoproteins. The O-linked glycans displayed by PSGL-1 must undergo two specific post-translational modifications in order for it to function as a counter-receptor for P-selectin: a(1,3) fucosylation and a(2,3) sialylation. Bonds between P-selectin and PSGL-1 primarily mediate the rolling phase of the adhesion cascade
 fucosylation and a(2,3) sialylation. Bonds between P-selectin and PSGL-1 primarily mediate the rolling phase of the adhesion cascade.")
20
Szelektin: specifikus kötődés sziálsavhoz és fukózhoz
Selectins bind to carbohydrate structures which contain the monosaccharides fucose and sialic acid (neuraminic acid). The prototype structure is the tetrasaccharide Sialyl-Lewis X (sLex) Leukocyte Adhesion Deficiency II (LAD II) Immundeficiencia, neutrofilia, pszihomotoros és növekedési retardáció
. The prototype structure is the tetrasaccharide Sialyl-Lewis X (sLex) Leukocyte Adhesion Deficiency II (LAD II) Immundeficiencia, neutrofilia, pszihomotoros és növekedési retardáció.")
21
Szelektinek szerepe: rolling
employees.csbsju.edu/.../complexoligosacch.htm

22
Integrinek

23
Integrinek Ált. sejt-ECM között de van sejt-sejt is
Heterodimer ( és ) sok variáció ECM receptorok pl. fibronektin, laminin, kollagén Ca2+ ion függő Kapcsoló fehérjéken keresztül mikro- vagy intermedier filamentumhoz Integrins comprise and subunits that span the cell membrane and reach into the extracellular space. Both and subunits have a short cytoplasmic tail on the inner side of the plasma membrane and a long extracellular domain protruding from the outer surface of the plasma membrane, which contacts the extracellular matrix (ECM). Non-covalent interactions between their extracellular and membrane-proximal regions bring and subunits together in a heterodimer. The short cytoplasmic tails of both and subunits of integrins interact with a cohort of intracellular proteins. These intracellular integrin-binding proteins also link to the cytoskeleton, thereby subjecting integrins to the mechanical forces that are generated by cytoskeletal proteins (right side of figure). A non-covalent interaction between the cytoplasmic tails of an inactive integrin prevents integrin activation (left side of figure). During 'inside–out' signalling, which influences integrin activation, certain signals or activators change the relationships of the and cytoplasmic tails with each other and/or the membrane, which leads to a propagated allosteric rearrangement.
 sok variáció. ECM receptorok pl. fibronektin, laminin, kollagén. Ca2+ ion függő. Kapcsoló fehérjéken keresztül. mikro- vagy intermedier. filamentumhoz. Integrins comprise and subunits that span the cell membrane and reach into the extracellular space. Both and subunits have a short cytoplasmic tail on the inner side of the plasma membrane and a long extracellular domain protruding from the outer surface of the plasma membrane, which contacts the extracellular matrix (ECM). Non-covalent interactions between their extracellular and membrane-proximal regions bring and subunits together in a heterodimer. The short cytoplasmic tails of both and subunits of integrins interact with a cohort of intracellular proteins. These intracellular integrin-binding proteins also link to the cytoskeleton, thereby subjecting integrins to the mechanical forces that are generated by cytoskeletal proteins (right side of figure). A non-covalent interaction between the cytoplasmic tails of an inactive integrin prevents integrin activation (left side of figure). During inside–out signalling, which influences integrin activation, certain signals or activators change the relationships of the and cytoplasmic tails with each other and/or the membrane, which leads to a propagated allosteric rearrangement.")
24
Integrinek: funkcionális forma: heterodimer
Heterodimer α integrin tagja: Ca2+ ion kötőhelyek

25
Integrin heterodimer kombinációk
ECM molekuláival kapcsolat RGD szekvencia imgt.cines.fr/.../Tutorials/Integrins/

26
Integrin és ECM molekula kapcsolat
szabályozása: kis G fehérjék Integrin α and β adhesiveness to extracellular matrix (ECM) components is regulated by altering the integrin's affinity for its ECM ligands and/or by altering the integrin's avidity. Small GTP-binding proteins seem to have a prevalent role in these processes. The activation of the small GTP-binding protein RRAS can lead to increases in integrin affinity for the ECM, whereas HRAS activation can lead to decreases in integrin affinity (no ECM binding). Similarly, activation of the small GTP-binding proteins RAC and CDC42, and protein kinase C (PKC), can lead to clustering of integrins, thereby increasing integrin avidity. The careful regulation of each of these events is necessary for efficient cell migration.
 components is regulated by altering the integrin s affinity for its ECM ligands and/or by altering the integrin s avidity. Small GTP-binding proteins seem to have a prevalent role in these processes. The activation of the small GTP-binding protein RRAS can lead to increases in integrin affinity for the ECM, whereas HRAS activation can lead to decreases in integrin affinity (no ECM binding). Similarly, activation of the small GTP-binding proteins RAC and CDC42, and protein kinase C (PKC), can lead to clustering of integrins, thereby increasing integrin avidity. The careful regulation of each of these events is necessary for efficient cell migration.")
27
Integrinek: speciális funkció
kötődhetnek szolubilis ligandhoz is pl. fibrinogén

28
Egyéb adhéziós molekulák….
bioweb.wku.edu/.../L21Adhesion SigTransdctn.html Sejtadhéziós molekulákkal kapcsolódnak ECM molekulákkal kapcsolódnak

29
Az igazi csapatmunka…….
Sequential steps in leukocyte emigration are controlled by specific adhesion molecules on leukocytes and endothelial cells. The various steps of leukocyte emigration described in the text are depicted schematically here. For each step the interacting pairs of adhesion molecules, ligands, or counter-receptors expressed by the leukocytes and endothelial cells or extracellular matrix are shown in the same color. This diagram is not inclusive, and other molecules may mediate each of these events for distinct leukocyte types under different inflammatory conditions. For the Capture/Tethering step, the protrusions on the leukocyte surface are meant to represent the microvilli that bear L-selectin or VLA-4. The lightning bolt at the Activation step represents the triggering of inside-out activation of leukocyte integrins by signals from the endothelium and endothelial surface via G protein coupled receptors. The basal lamina (orange strip) is depicted as separate from the remainder of the extracellular matrix (ECM), because migration across the subendothelial basal lamina seems to be a separate step controlled by distinct molecules (Liao et al, 1995; Wakelin et al, 1996). However, these steps may not be molecularly distinct (see text). Because the exact 1 integrin(s) that are involved in migration through ECM via fibronectin interactions have not been defined, the leukocyte molecules are designated simply as 1. ESL, E-selectin ligand; HSPG, heparan sulfate proteoglycan; ICAM, intercellular adhesion molecule; LFA, leukocyte function associated antigen; PAF, platelet activating factor; PAF-R, PAF receptor; PECAM-1, platelet/endothelial cell adhesion molecule-1; PECAM-1 (d1/2), interaction involves immunoglobulin domains 1 and/or 2 of PECAM-1; PECAM-1 (d6), interaction involves immunoglobulin domain 6 of PECAM-1; PSGL, P-selectin glycoprotein ligand; s-Lex, sialyl-Lewisx carbohydrate antigen; VLA, very late antigen.
 is depicted as separate from the remainder of the extracellular matrix (ECM), because migration across the subendothelial basal lamina seems to be a separate step controlled by distinct molecules (Liao et al, 1995; Wakelin et al, 1996). However, these steps may not be molecularly distinct (see text). Because the exact 1 integrin(s) that are involved in migration through ECM via fibronectin interactions have not been defined, the leukocyte molecules are designated simply as 1. ESL, E-selectin ligand; HSPG, heparan sulfate proteoglycan; ICAM, intercellular adhesion molecule; LFA, leukocyte function associated antigen; PAF, platelet activating factor; PAF-R, PAF receptor; PECAM-1, platelet/endothelial cell adhesion molecule-1; PECAM-1 (d1/2), interaction involves immunoglobulin domains 1 and/or 2 of PECAM-1; PECAM-1 (d6), interaction involves immunoglobulin domain 6 of PECAM-1; PSGL, P-selectin glycoprotein ligand; s-Lex, sialyl-Lewisx carbohydrate antigen; VLA, very late antigen.")
30
Adhéziós molekula kapcsolatok kadherinekkel és
integrinekkel:Sejtkapcsoló struktúrák

31
Sejtkapcsoló struktúrák kialakulása
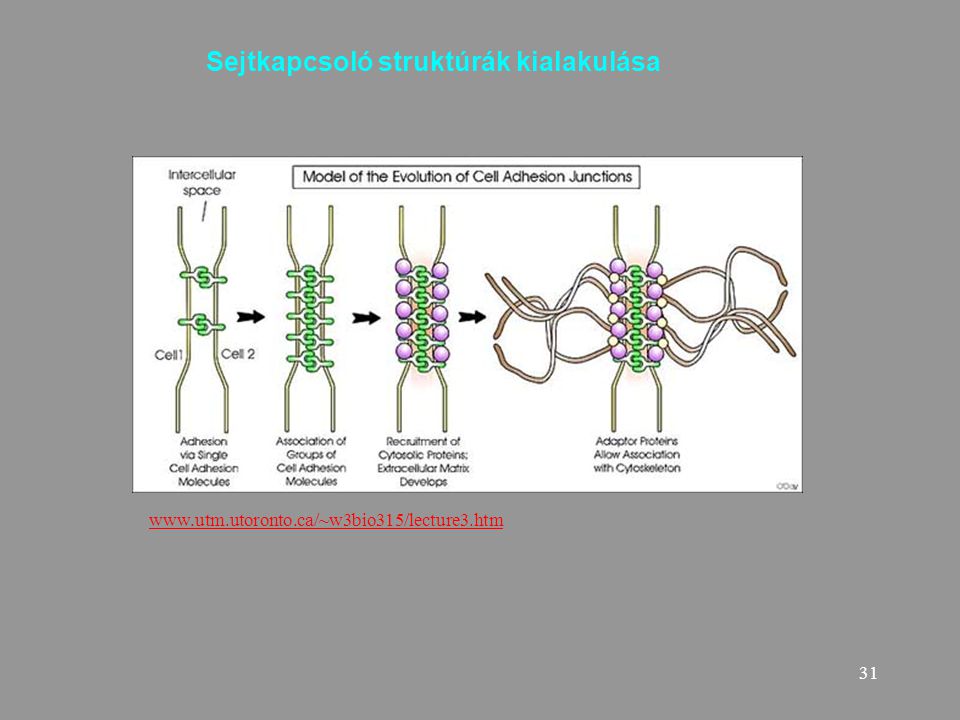
32
A sejtkapcsoló struktúrák jellemzése
Sejt-sejt, sejt-mátrix Ion (Ca++, Mg++) függőség Állandó vagy időleges Kapcsolódó sejtváz elemek Funkció: Lezárás és mechanikai kapcsolás Mechanikai kapcsolás Kommunikáció
 függőség. Állandó vagy időleges. Kapcsolódó sejtváz elemek. Funkció: Lezárás és mechanikai kapcsolás. Mechanikai kapcsolás. Kommunikáció.")
33
sejtkapcsoló struktúrák
Hámsejtek laterális és bazális felszínén

34
Sejt-sejt kapcsoló struktúrák
Szoros illeszkedés (tight junction) Zonula occludens Adherens kapcsolat (adherent junction) Zonula adherens Dezmoszóma + interdigitáció Macula adherens Réskapcsolat Gap junction
 Zonula occludens. Adherens kapcsolat (adherent junction) Zonula adherens. Dezmoszóma + interdigitáció. Macula adherens. Réskapcsolat. Gap junction.")
35
Sejt-sejt és sejt - mátrixkapcsoló struktúrák
hemidezmoszóma Fokális kontaktus

36
Szoros illeszkedés Adhéziós molekulák: okkludin, klaudin, JAM-ok
Figure 2. Schematic diagram of the molecular organization of epithelial TJ. TJ restrict the free diffusion of molecules across the space between the membranes of adjoining cells, and maintain a separation between apical and basolateral membrane domains. TJ membrane proteins are linked to the cytoskeleton (F-actin) via a complex network of adaptor proteins. Several TJ proteins interact with regulators of small GTPases (Cdc42, RhoA and Rac1). Some transcription factors and cytoplasmic TJ proteins shuttle between junctions and nucleus (arrows) depending on the state of confluence of the epithelial monolayer, indicating a junction-assembly-dependent mechanism of transcriptional regulation. Adhéziós molekulák: okkludin, klaudin, JAM-ok Kapcsoló fehérjék: ZO1,2, cingulin Sejtváz elem: mikrofilamentum (aktin)
 via a complex network of adaptor proteins. Several TJ proteins interact with regulators of small GTPases (Cdc42, RhoA and Rac1). Some transcription factors and cytoplasmic TJ proteins shuttle between junctions and nucleus (arrows) depending on the state of confluence of the epithelial monolayer, indicating a junction-assembly-dependent mechanism of transcriptional regulation. Adhéziós molekulák: okkludin, klaudin, JAM-ok. Kapcsoló fehérjék: ZO1,2, cingulin. Sejtváz elem: mikrofilamentum (aktin)")
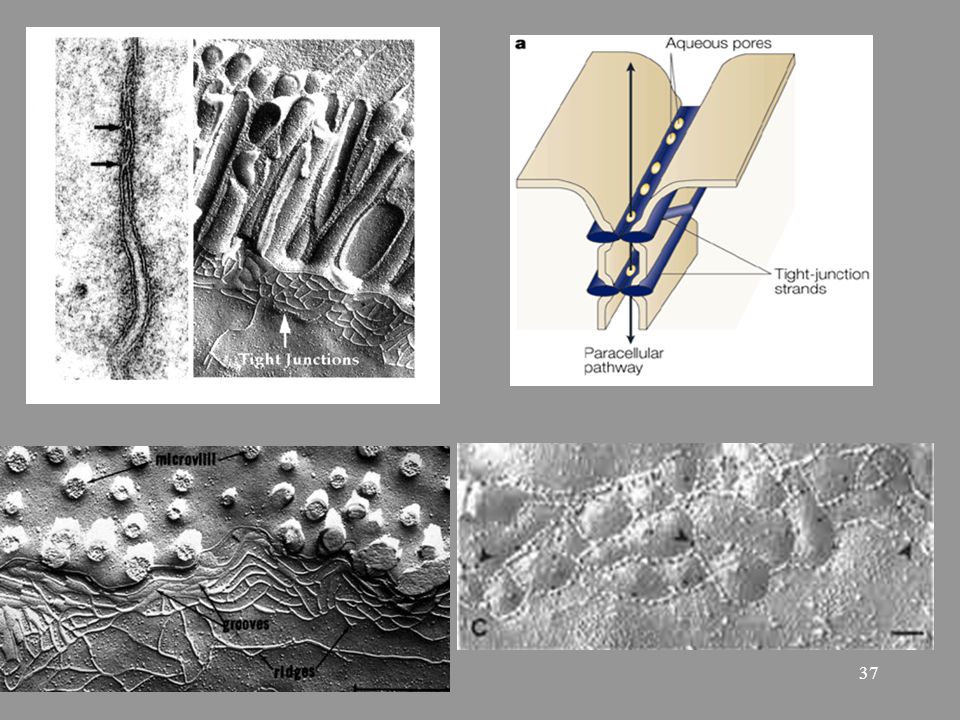
38
Szoros illeszkedés szerepe
-mechanikai kapcsolat - paracelluláris transzport szabályozása -apikális és laterális membrán domének megtartása

39
Zonula adherens:Mechanikai kapcsolat
Fő adhéziós molekula: E cadherin Kapcsoló fehérjék. Kateninek Aktin filamentumhoz kapcsolódik

40
Adherens sejtkapcsolat és egy szignalizációs útvonal
Howard et al. BMC Musculoskeletal Disorders :16 doi: / Canonical Wnt/β-catenin pathway. β-catenin is a component of cell-cell adhesion structures (adherens junctions) [20-22] and a key signaling factor in the Wnt pathway [10]. As shown here canonical Wnt signalling (Wnt/β-catenin) is defined by its inhibition of glycogen synthase kinase-3β (GSK-3β) catalyzed phosphorylation of β-catenin. Several factors, including Cer (cerebrus), WIF-1 (wnt-interacting protein), and sFRPs (secreted frizzled related proteins) and dickkopf-1 (Dkk) are known to antagonize Wnt signalling [63-69]. However, upon Wnt stimulation several Fz-LRP downstream signalling components, including the phosphoprotein dishevelled (Dvl) [70-72], GBP/Frat1 (GSK-3β Binding Protein) [73] and casein kinase I (CKIε) [74,75], somehow co-operate to inhibit GSK-3β. This ultimately this leads to an increase in the cytosolic 'free' levels of β-catenin (uncomplexed to cadherin), and its accumulation within the nucleus where it binds to members of the Tcf/Lef (T-cell factor-lymphoid enhancer factor) transcription factor family to activate gene transcription in a cell-context dependent manner [28-34]. In the absence of Wnt signalling, Axin/conductin [76,77] in co-operation with the product of the tumour suppressor gene adenomatous polyposis coli (APC) [78,79] facilitate GSK-3β mediated phosphorylation of β-catenin on N-terminal serine and threonine residues [80]. This hyper-phosphorylated form of β-catenin then binds to the F-box protein β TrCP, which targets β-catenin for degradation via the ubiquitin-proteosome pathway [81-84].
 [20-22] and a key signaling factor in the Wnt pathway [10]. As shown here canonical Wnt signalling (Wnt/β-catenin) is defined by its inhibition of glycogen synthase kinase-3β (GSK-3β) catalyzed phosphorylation of β-catenin. Several factors, including Cer (cerebrus), WIF-1 (wnt-interacting protein), and sFRPs (secreted frizzled related proteins) and dickkopf-1 (Dkk) are known to antagonize Wnt signalling [63-69]. However, upon Wnt stimulation several Fz-LRP downstream signalling components, including the phosphoprotein dishevelled (Dvl) [70-72], GBP/Frat1 (GSK-3β Binding Protein) [73] and casein kinase I (CKIε) [74,75], somehow co-operate to inhibit GSK-3β. This ultimately this leads to an increase in the cytosolic free levels of β-catenin (uncomplexed to cadherin), and its accumulation within the nucleus where it binds to members of the Tcf/Lef (T-cell factor-lymphoid enhancer factor) transcription factor family to activate gene transcription in a cell-context dependent manner [28-34]. In the absence of Wnt signalling, Axin/conductin [76,77] in co-operation with the product of the tumour suppressor gene adenomatous polyposis coli (APC) [78,79] facilitate GSK-3β mediated phosphorylation of β-catenin on N-terminal serine and threonine residues [80]. This hyper-phosphorylated form of β-catenin then binds to the F-box protein β TrCP, which targets β-catenin for degradation via the ubiquitin-proteosome pathway [81-84].")
41
Dezmoszóma:Erős mechanikai kapcsolat
Kapcsoló fehérjék: desmoglein és desmokollin desmoszómális kadherinek Kapcsoló fehérjék : plakoglobin, desmoplakin Sejtváz: intermedier filamentum

42
Stratum spinosum dezmoszóma

43
Réskapcsolat (gap junction)
Kommunikációs sejt-sejt kapcsolat konnexon Fehérje: konnexinek Connexin molekula elhelyezkedése a membránban

44
A réskapcsolat felépülése konnexin fehérjékből
Példa kombinációkra

45
Homo- és heterotpikus réskapcsolatok a retina sejtjei között
The presence of gap junction coupling as a result of connexin 36 (CX36) expression has been shown between cones81, and are also assumed to contain CX36, on the cone side, between cones and rods81. CX36-containing gap junction channels also occur beween the dendrites of OFF cone bipolar cells (OFF CB)81. Horizontal cells (H) are coupled by CX57-containing gap junctions29. During scotopic light conditions, the network of AII amacrine cells (AII), which are connected to each other by CX36-containing gap junctions, mediates signalling from the rod to the cone signal transmission pathway by heterotypic electrical synapses between AII amacrine cells (CX36) and ON cone bipolar cells (CX45)28. Homotypic CX36 gap junction channels have been reported to be present between alpha ganglion cells ( GC)82, 84. Note that the schematic topology of all the electrical synapses shown needs to be verified by further research. C, cone photoreceptor cell; CB, cone bipolar cell; GC, ganglion cell; GCL, ganglion cell layer; INL, inner nuclear layer; IPL, inner plexiform layer; IS, inner segment; NFL, optic nerve fibre layer; ONL, outer nuclear layer; OPL, outer plexiform layer; OS, outer segment; R, rod photoreceptor cell; RB, rod bipolar cell.
 expression has been shown between cones81, and are also assumed to contain CX36, on the cone side, between cones and rods81. CX36-containing gap junction channels also occur beween the dendrites of OFF cone bipolar cells (OFF CB)81. Horizontal cells (H) are coupled by CX57-containing gap junctions29. During scotopic light conditions, the network of AII amacrine cells (AII), which are connected to each other by CX36-containing gap junctions, mediates signalling from the rod to the cone signal transmission pathway by heterotypic electrical synapses between AII amacrine cells (CX36) and ON cone bipolar cells (CX45)28. Homotypic CX36 gap junction channels have been reported to be present between alpha ganglion cells ( GC)82, 84. Note that the schematic topology of all the electrical synapses shown needs to be verified by further research. C, cone photoreceptor cell; CB, cone bipolar cell; GC, ganglion cell; GCL, ganglion cell layer; INL, inner nuclear layer; IPL, inner plexiform layer; IS, inner segment; NFL, optic nerve fibre layer; ONL, outer nuclear layer; OPL, outer plexiform layer; OS, outer segment; R, rod photoreceptor cell; RB, rod bipolar cell.")
46
Kis metabolitok (pl Glukóz-P) megoszlása
Réskapcsolat: metabolikus kapcsolás Kizárási határ: M<1000 Parciális szincícium Kis metabolitok (pl Glukóz-P) megoszlása Jeltovábbítás Másodlagos hírvivők, pl. cAMP Ionok
 megoszlása. Jeltovábbítás. Másodlagos hírvivők, pl. cAMP. Ionok.")
47
Réskapcsolat működése
Konformáció változás regulálja hogy a csatorna nyitott vagy csukott Ca2+ ionok szerepe

48
Réskapcsolat és szignál útvonalak kapcsolata

49
Sejt-mátrix kapcsolatok
Hemidezmoszóma Fokális kontaktus

50
erős mechanikai kapcsolat
Hemidezmoszómák: erős mechanikai kapcsolat Adhéziós molekula:integrin Sejtváz elem: intermedier filamentum Getting under the skin of epidermal morphogenesis Elaine Fuchs & Srikala Raghavan Nature Reviews Genetics 3, (March 2002) bazális lamina komponenseivel kapcsolódnak az integrinek
 bazális lamina komponenseivel kapcsolódnak az integrinek.")
51
Fokális kontaktus FAK: fokális adhéziós kináz
Aktin filamentumok (stress rostok) FAK: fokális adhéziós kináz Adhéziós molekulák: integrinek
 FAK: fokális adhéziós kináz. Adhéziós molekulák: integrinek.")
52
Jelátviteli utakon át számos folyamat kapcsolódik össze
Jelátviteli utakon át számos folyamat kapcsolódik össze

53
Dezmoszómális betegségek, kórképek
3 típusú betegség: 1. Autoimmun eredetű : pl. pemphigus vulgaris 2. öröklött ioncsatorna betegségek: Hailey-Hailey disease and Darier disease 3. Öröklött dezmoszóma fehérje mutációk okozta betegségek Speciális eset: baktérium toxin kiváltotta kórkép : Staphylococcus scalded-skin syndrome

54
Table 1. Mutations and target antigens in desmosomal diseases
Protein Perturbation Diseases Dsg1 Haploinsufficiency SPPK Autoantibodies Pemphigus foliaceus Exfoliative toxin Staphylococcus scalded-skin syndrome Dsg3 Pemphigus vulgaris Dsg4 Hair follicle abnormalities, hair loss Autosomal recessive hypotrichosis Plakoglobin C-terminal truncation Naxos disease (PPK, woolly hair, ARVC) Plakophilin-1 Compound heterozygous mutations (premature stop codons and absence of protein) Ectodermal dysplasia and skin fragility syndrome Plakophilin-2 Various heterozygous mutations and haploinsufficiency ARVC Desmoplakin Compound heterozygosity with missense (N-terminal) and nonsense (C-terminal) mutations Keratoderma, keratin retraction, skin fragility and woolly hair/alopecia N-terminal missense mutation Autosomal-dominant ARVC C-terminal missense Dilated left ventricular cardiomyopathy, SPPK, woolly hair (Carvajal syndrome) Compound heterozygous mutations with C-terminal truncation Lethal acantholytic epidermolysis bullosa Abbreviations: ARVC, arrhythmogenic right-ventricular cardiomyopathy; PPK, palmoplantar keratoderma; SPPK, striate palmoplantar keratoderma
 Plakophilin-1. Compound heterozygous mutations (premature stop codons and absence of protein) Ectodermal dysplasia and skin fragility syndrome. Plakophilin-2. Various heterozygous mutations and haploinsufficiency. ARVC. Desmoplakin. Compound heterozygosity with missense (N-terminal) and nonsense (C-terminal) mutations. Keratoderma, keratin retraction, skin fragility and woolly hair/alopecia. N-terminal missense mutation. Autosomal-dominant ARVC. C-terminal missense. Dilated left ventricular cardiomyopathy, SPPK, woolly hair (Carvajal syndrome) Compound heterozygous mutations with C-terminal truncation. Lethal acantholytic epidermolysis bullosa. Abbreviations: ARVC, arrhythmogenic right-ventricular cardiomyopathy; PPK, palmoplantar keratoderma; SPPK, striate palmoplantar keratoderma.")
55
Erythema (bőrpír), peeling (hámlás), and necrosis (szövetelhalás)
Gyógyítható, 2% alatti a mortalitás Staphylococcus aureus: exotoxinok okozzák Desmoglein 1 károsodása

56
Autoantitest termelődés dsg3 ellen
Pemphigus vulgaris Expression pattern of desmosomal components in the epidermis Desmoglein 3 (dsg3) a bazális és suprabazális rétegre jellemző Autoantitest termelődés dsg3 ellen
 a bazális. és suprabazális rétegre jellemző. Autoantitest termelődés dsg3 ellen.")
57
Clinical appearance and pathohistology of various human desmosomal disorders. Pemphigus vulgaris (A and B) is characterized by the loss of intercellular adhesion between basal and suprabasal keratinocytes (A) and by skin blistering and erosions (B). The hallmark of palmoplantar keratoderma (C and D) is massive thickening of the stratum corneum (C), resulting in dramatically thickened skin on palms and soles (D). Arrhythmogenic right-ventricular cardiomyopathy (ARVC) is characterized by fibrofatty replacement (*) of the myocardium (E). Panel D is reprinted from (Rickman et al., 1999) with permission from Oxford University Press. Panel E is reprinted from (Protonotarios and Tsatsopoulou, 2004) with permission from Elsevier. Plakofillin mutáció Journal of Cell Science 119, (2006) The desmosome: cell science lessons from human diseases Margaret D. Kottke1, Emmanuella Delva2 and Andrew P. Kowalczyk1,2
 is massive thickening of the stratum corneum (C), resulting in dramatically thickened skin on palms and soles (D). Arrhythmogenic right-ventricular cardiomyopathy (ARVC) is characterized by fibrofatty replacement (*) of the myocardium (E). Panel D is reprinted from (Rickman et al., 1999) with permission from Oxford University Press. Panel E is reprinted from (Protonotarios and Tsatsopoulou, 2004) with permission from Elsevier. Plakofillin mutáció. Journal of Cell Science 119, (2006) The desmosome: cell science lessons from human diseases. Margaret D. Kottke1, Emmanuella Delva2 and Andrew P. Kowalczyk1,2.")
58
woolly hair (gyapjas haj)
Naxos betegség: woolly hair (gyapjas haj) Palmoplantar keratoma (tenyér-talp szarusodás) ARV jobb kamrát érintő kardiomyopáthia Plakoglobin mutáció commons.wikimedia.org/wiki/Image:Naxos_diseas...
 Palmoplantar keratoma (tenyér-talp szarusodás) ARV jobb kamrát érintő kardiomyopáthia. Plakoglobin mutáció. commons.wikimedia.org/wiki/Image:Naxos_diseas...")
59
Colon tumor és katenin Gyomor tumor és E kadherin Ovárium tumor és E kadherin

60
vinkulin talin citokeratin E kadherin Ez is N CAM:
lpmt090.biomed.uni-erlangen.de/~gdiez/index.html vinkulin talin Ez is N CAM: North Cadbury Academy of Music citokeratin E kadherin

Hasonló előadás













 7) 8) 9) 10) Mennyi az x, y és z értéke? 11) 12) 13) 14) 15)>")





