Előadást letölteni
Az előadás letöltése folymat van. Kérjük, várjon

1
Fejezetek a sejtbiológiából Teloméra és telomeráz. Sejtmagvacska
Dr. Tóth Sára SE Genetikai, Sejt- és Immunbiológiai Intézet

3
Telomérák kimutatása in situ hibridizációval

4
A teloméra ismétlődő (repetitív) szekvenciákban gazdag
Teloméra ~ 30 repeat Szubtelomerikus régió ~ 2000 repeat

5
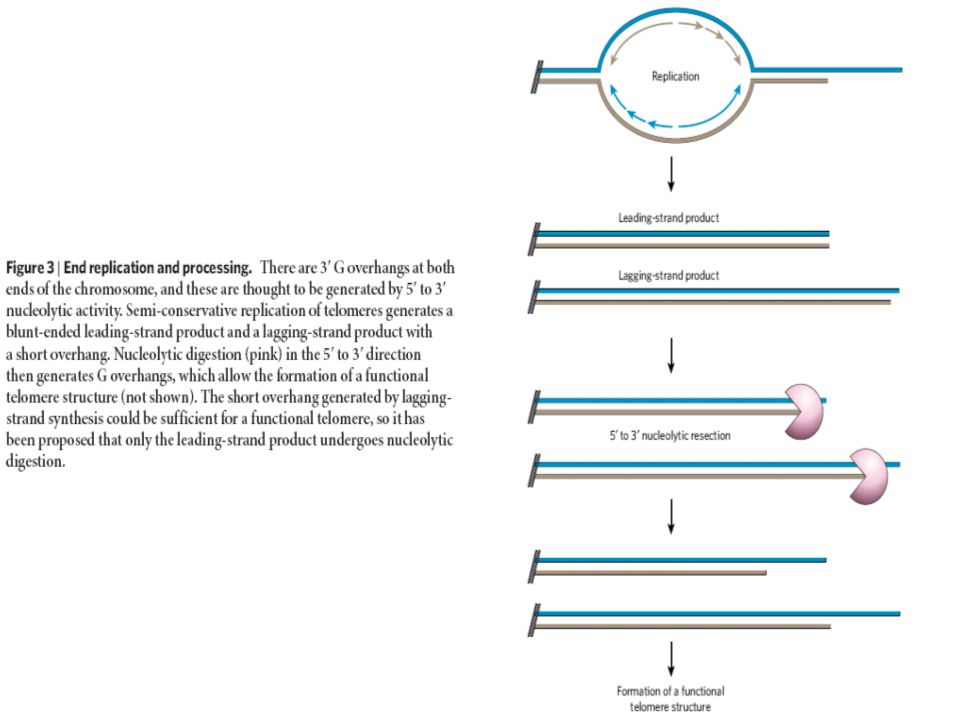
Egészséges szomatikus sejtekben a telomérák
osztódásról osztódásra rövidülnek

6
De vannak kivételek: csírasejtek és tumorsejtek

8
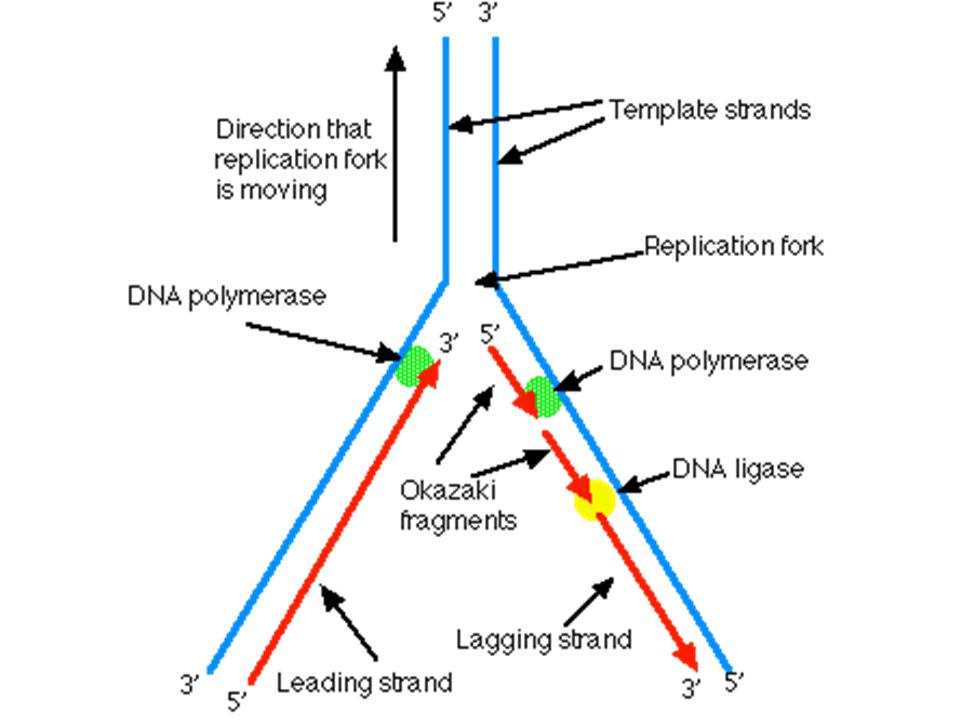
S fázis (DNS szintézis)
2 replikációs villa primáz RNS primer DNA polimeráz 5’ 3’ monomerek: dezoxinukleozid trifoszfátok leading (vezető) és lagging (elmaradó) szál primer kivágása ligáz szemi-konzervatív Azonos DNS molekulák
 és. lagging (elmaradó) szál. primer kivágása. ligáz. szemi-konzervatív. Azonos DNS molekulák.")
10
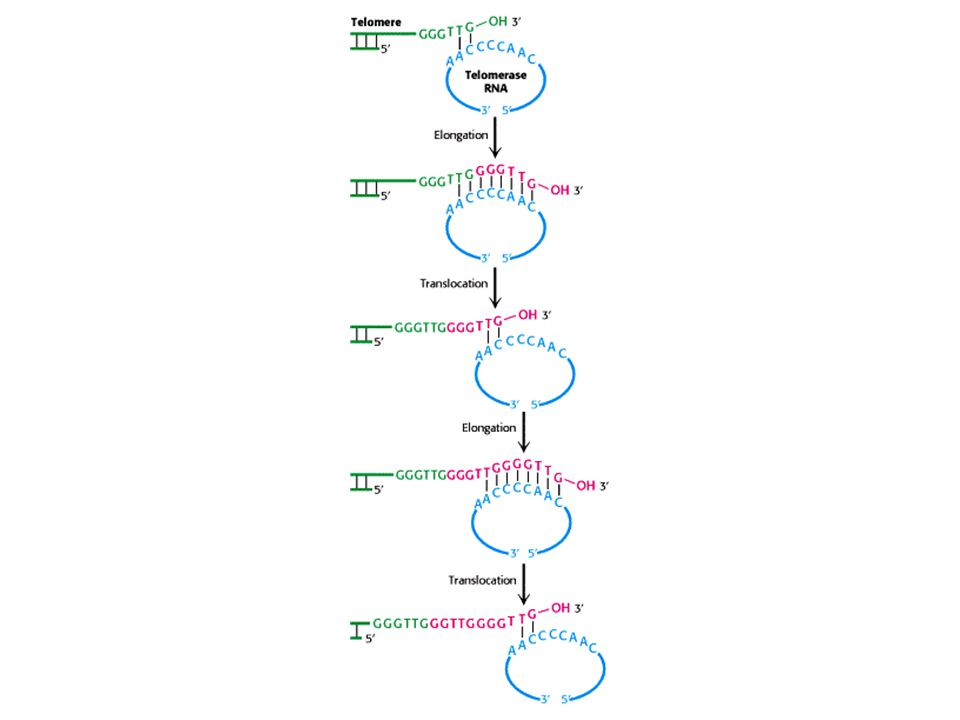
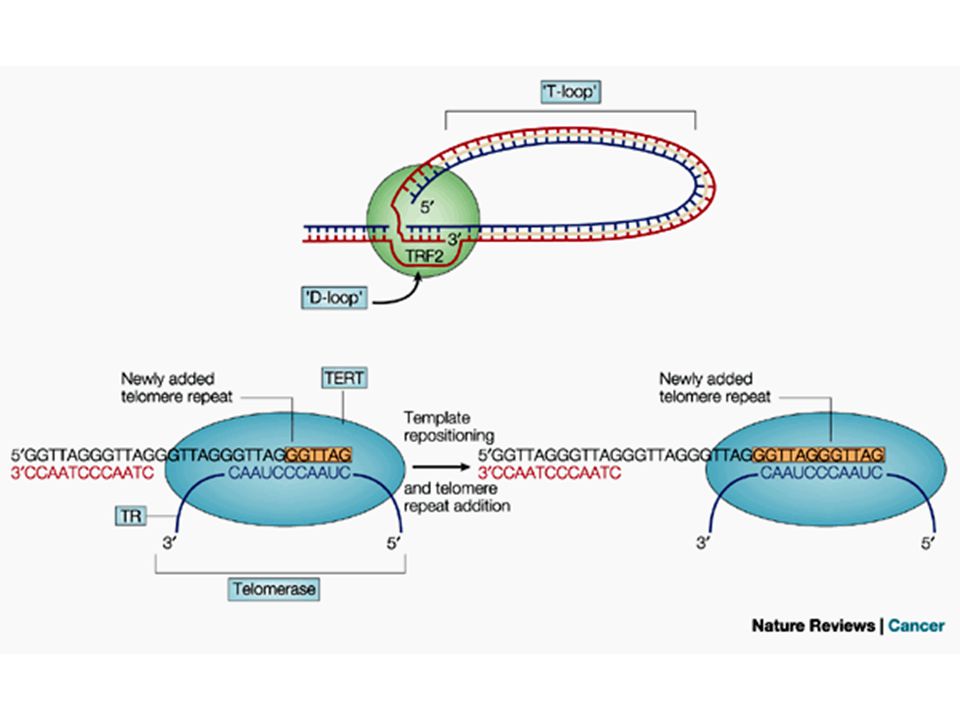
A telomeráz RNS dependens DNS szintézist végez;
reverz transzkriptáz

11
A telomeráz fő részei a TERT = az enzim,
és a TERC = az RNS templátot tartalmazza a TERT felépítése

14
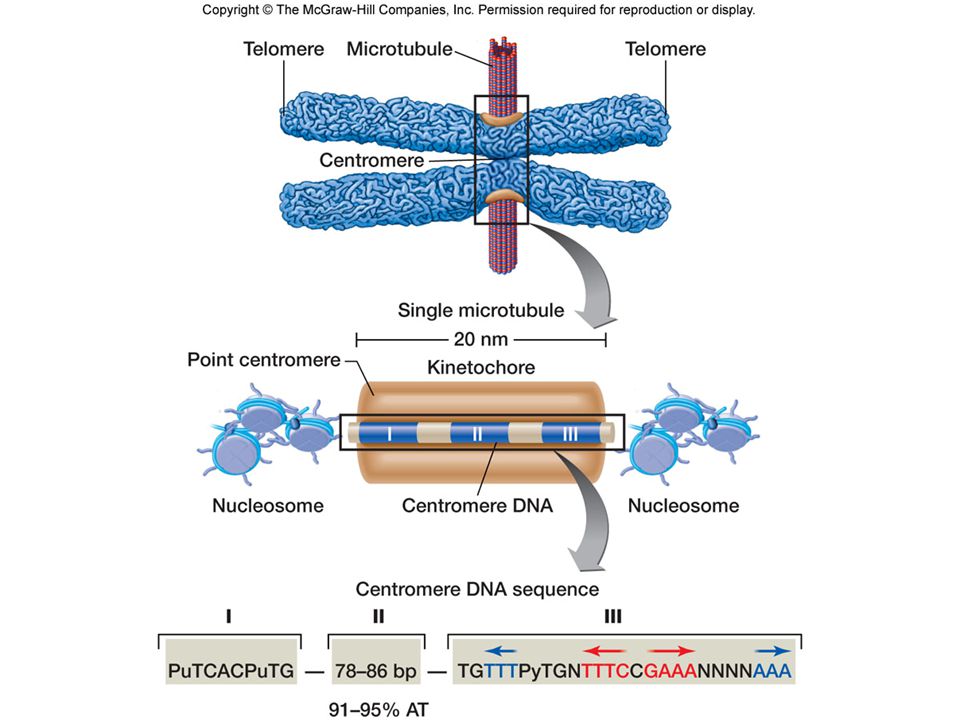
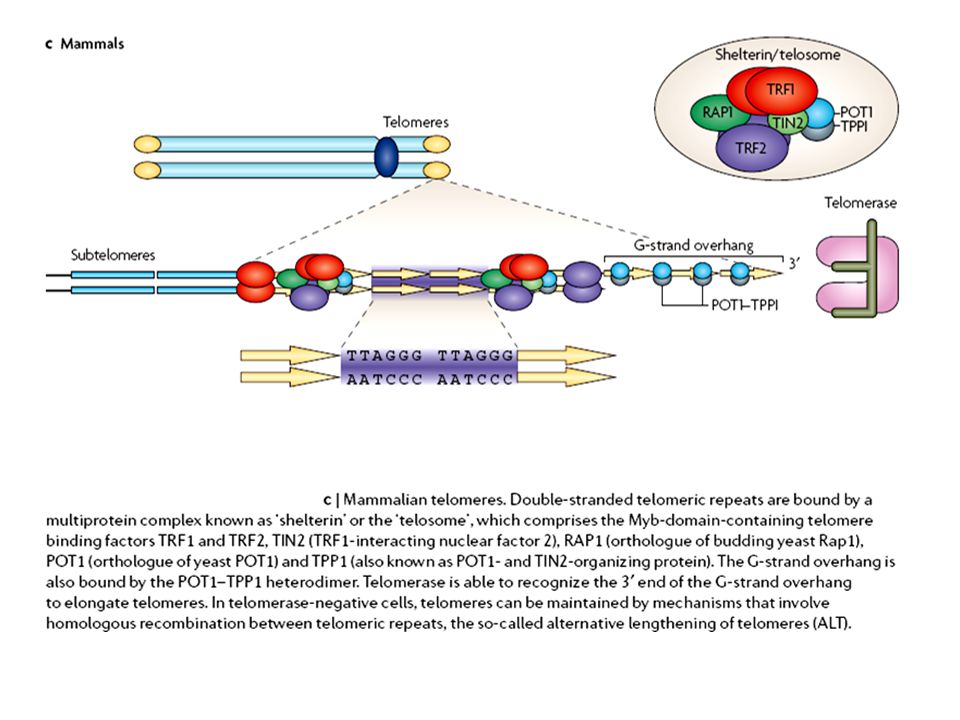
A telomérák sajátos szerveződésű és szerkezetű DNS-t
tartalmaznak G-gazdag Visszahurkolódó Shelterin komplex jellemzi („sapkázás)
")
15
A teloméra térszerkezete
G-kvadruplex

17
A visszahurkolódás korlátozza a telomeráz telomérához való
hozzáférését

18
és a komponensek hibái által okozott betegségek
A teloméra szerkezete és a komponensek hibái által okozott betegségek

19
A telomérahossz telomeráz általi fenntartása

22
Bizonyos tumorokat (telomeráz negatívak) alternatív teloméra
hosszabítás (ALT) jellemez, amely homológ rekombináción alapul
 jellemez, amely homológ rekombináción. alapul.")
23
az alternatív teloméra hosszabbítás
PML body A PML testecske egyik lehetséges funkciója az alternatív teloméra hosszabbítás

24
A telomérák és a hisztonmódosulások – kromatinszerkezet kapcsolata
AcH4 = acetilált H4 hiszton; AcH3 = acetilált H3 hiszton; 5metC = DNS metilált citozinja; H3K9me = H3 hiszton metilált 9. lizinnel; H4K20me = H4 hiszton metilált 20. lizinnel; HP1 = heterokromatin protein 1; TRF = TERF = telomeráz repeat-kötő faktor; RB = retinoblasztoma proteinek; DNMT = DNS metil-transzferáz; SUV = Hiszton metil-transzferáz

25
A telomérák és a hisztonmódosulások – kromatinszerkezet kapcsolata
AcH4 = acetilált H4 hiszton; AcH3 = acetilált H3 hiszton; 5metC = DNS metilált citozinja; H3K9me = H3 hiszton metilált 9. lizinnel; H4K20me = H4 hiszton metilált 20. lizinnel; HP1 = heterokromatin protein 1; TRF = TERF = telomeráz repeat-kötő faktor; HMT = hiszton metil-transzferáz; RB = retinoblasztoma proteinek; DNMT = DNS metil-transzferáz

26
A telomérák és a hisztonmódosulások – kromatinszerkezet kapcsolata
AcH4 = acetilált H4 hiszton; AcH3 = acetilált H3 hiszton; 5metC = DNS metilált citozinja; H3K9me = H3 hiszton metilált 9. lizinnel; H4K20me = H4 hiszton metilált 20. lizinnel; HP1 = heterokromatin protein 1; TRF = TERF = telomeráz repeat-kötő faktor

27
Az emlős telomérák és szubtelomerikus régiók epigenetikus módosulásai
Epigenetikus jelenségek: a DNS szekvenciát nem érintő, de expresszióját befolyásoló jelenségek pl. DNS metiláció, hiszton módosulások, kromatin remodellezés

28
A nem jól működő, illetve rövid telomérák feltételezett szerepe
az emberi betegségekben

29
A rövid telomérákkal kapcsolatos
- a csontvelő működés zavarával járó - kórképek

30
A teloméra rövidülés az öregedés és a rák kapcsolata

31
Az öregedés, a rák és a telomérák kapcsolata

32
Werner-szindróma

33
A Werner-szindrómára jellemző sejtfenotípus megfordítható telomeráz expresszáltatásával
A TERT-tel (telomeráz) transzformált dermális fibroblasztok folyamatosan osztódnak, míg a Werner szindrómás sejtek általában 20 osztódást követően leállnak az osztódással.
 transzformált dermális fibroblasztok folyamatosan osztódnak, míg a Werner szindrómás sejtek általában 20 osztódást követően leállnak az osztódással.")
34
Teloméra rövidülést gyorsító tényezők
Sterssz Dohányzás Kövérség Korai öregedéssel járó kórképek Ataxia telangiectasia (ATM) Werner szindróma (WR) Bloom szindróma (BLM) Dyskeratosis congenita (KC1, TERC) Aplasztikus anémia (TERC, TERT) Fanconi anémia (Fans gének) Nijmegen kromoszómatörési szindróma (NBN)
 Werner szindróma (WR) Bloom szindróma (BLM) Dyskeratosis congenita (KC1, TERC) Aplasztikus anémia (TERC, TERT) Fanconi anémia (Fans gének) Nijmegen kromoszómatörési szindróma (NBN)")
35
Daganat- és áttétképzés kapcsolata a teloméra rövidüléssel

36
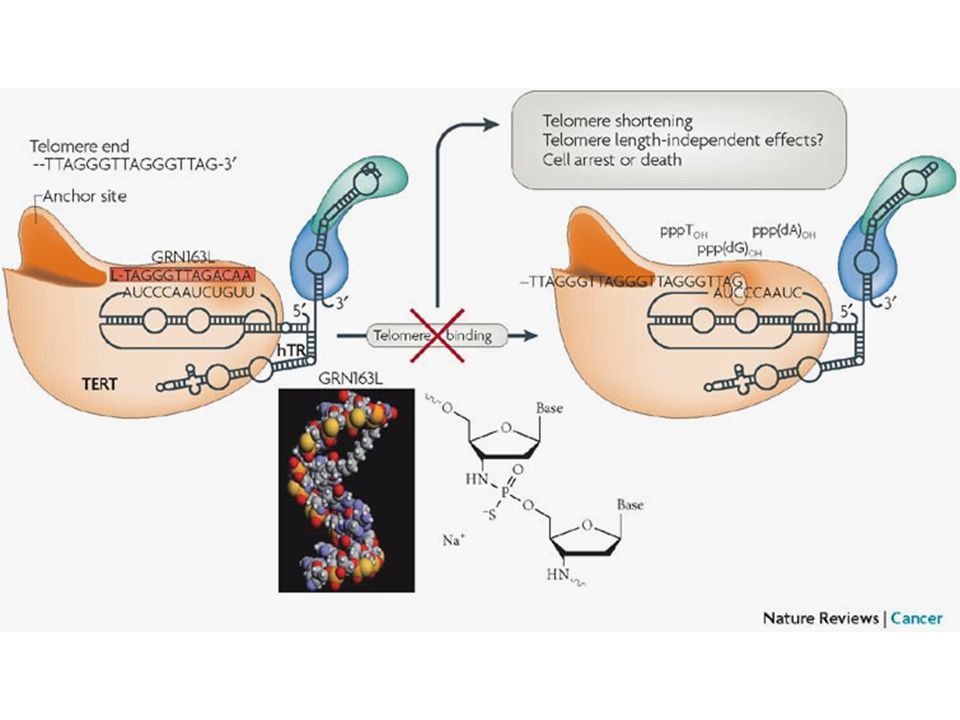
Terápiás lehetőségek Telomeráz gátlás Aktív immunterápia Telomérát roncsoló ágensek Szuicid génterápia Telomeráz expressziót vagy biogenezist gátló szerek

37
Terápiás lehetőségek I.

38
Terápiás lehetőségek II.

39
Egyéb sejtmag alkotók: sejtmagvacska

40
A sejtmagvacska részei
NO = nukleolusz organizátor = FC = fibrilláris centrum PG = pars granulosa = GK = granuláris kompartment PF = pars fibrosa = DFK= denz fibrilláris kompartment NAC = nukleolusz asszociált kromatin

42
Nucleolar organizer region = NOR = nukleolusz organizátor régió:
Ismétlődő rDNS szekvenciákból áll

43
Kétéltű oocyta lámpakefe kromoszómái
Génamplifikáció rDNS sokszorozódás sok rRNS

44
A sejtciklus során a sejtmagvacskát érintő változások
szétesés fúzió

45
Sejtmagvacska fúzió

46
A riboszóma keletkezés kinetikája és az rRNS splicingja

47
rRNS szintézis a sejtmagvacskában

48
A sejtmagvacskában zajló folyamatok
rRNS szintézis rRNS érés (splicing) riboszóma alegység összeszerelés snoRNA= small nucleolar RNA
 riboszóma alegység. összeszerelés. snoRNA= small nucleolar RNA.")
49
A riboszómák keletkezése és funkciója
= módosult DNS-bázis DNS-bázis

50
A prokaryota és eukaryota riboszómák összehasonlítása

51
A sejtmagvacska és a mitózis
NDF = nucleolar derived foci; PNB = prenucleolar body; PR= perichromosomal region A sejtmagvacska és a mitózis

52
A sejtmagvacska fehérjéinek funkcionális megoszlása
emberi növényi

53
Riboszóma alegységek összeszerelődése

54
Riboszóma alegységek összeszerelődése II.
Biogenesis of ribosomes represents a highly dynamic process which is precisely coordinated in a temporal and spatial fashion. More than 150 non-ribosomal factors and approximately 70 ribosomal proteins as well as several RNAs participate in this process. Formation of heterodimeric Noc-proteins is essential for assembly, maturation and transport of both ribosomal subunits. We investigate how generation and dynamics of pre-ribosomes are controlled through the formation of different Noc-complexes. Furthermore we are analysing how the communication between maturation of ribosomes, rDNA transcription and the cell cycle occurs.

55
Az rRNS érésében szerepe van a snoRNS-ek által irányított
bázismódosításoknak (uridin–pszeudouridin csere és metiláció) is All ribosomal RNAs, with the exception of the 5S, are transcribed as a polycistronic transcript known as pre-ribosomal RNA (pre-rRNA or 45S) in the nucleolus. Transcription of rDNA is dependent on three basal transcription factors: the 'selectivity complex' (SL1 or TIF-IB), the HMG1 box architectural upstream binding factor (UBF) and the DNA-dependent RNA polymerase I (Pol I). Both UBF and SL1 make direct contacts with a 150-bp region of DNA in the rRNA gene promoter, which contains two sequence elements known as the upstream control element (UCE) and the core. In the case of UBF, it probably binds to these promoter elements as a dimer. UBF is thought to bind the promoter first, enabling subsequent recruitment of SL1 and Pol I. The initiation factor TIF-IA is also part of this complex. Concomitant with rRNA transcription, the rRNA sequences are extensively modified. Specifically, a large family of small nucleolar RNAs (snoRNAs) guides the site-specific conversion of uridine to pseudouridine ( ) in rRNA. This is accomplished through direct base-pairing of snoRNAs with specific rRNA sequences, leaving a single uridine exposed to the enzymatic activity of dyskerin, the pseudouridine synthase that mediates the modification of this residue. In addition, other snoRNAs also guide the formation of 2'-0-methylated nucleosides (-CH3) in rRNA. The pre-rRNA precursor is then cleaved at specific sites by RNases to produce a series of characteristic intermediates (41S), and finally mature rRNAs — 18S, 5.8S and 28S. During rRNA processing, the rRNA species must associate with more than 70 ribosomal proteins, as well as the 5S rRNA in the nucleolus, to form the small (S, 40S) and the large (L, 60S) ribosomal subunits, which are assembled and transported to the cytoplasm to initiate protein synthesis.
 is. All ribosomal RNAs, with the exception of the 5S, are transcribed as a polycistronic transcript known as pre-ribosomal RNA (pre-rRNA or 45S) in the nucleolus. Transcription of rDNA is dependent on three basal transcription factors: the selectivity complex (SL1 or TIF-IB), the HMG1 box architectural upstream binding factor (UBF) and the DNA-dependent RNA polymerase I (Pol I). Both UBF and SL1 make direct contacts with a 150-bp region of DNA in the rRNA gene promoter, which contains two sequence elements known as the upstream control element (UCE) and the core. In the case of UBF, it probably binds to these promoter elements as a dimer. UBF is thought to bind the promoter first, enabling subsequent recruitment of SL1 and Pol I. The initiation factor TIF-IA is also part of this complex. Concomitant with rRNA transcription, the rRNA sequences are extensively modified. Specifically, a large family of small nucleolar RNAs (snoRNAs) guides the site-specific conversion of uridine to pseudouridine ( ) in rRNA. This is accomplished through direct base-pairing of snoRNAs with specific rRNA sequences, leaving a single uridine exposed to the enzymatic activity of dyskerin, the pseudouridine synthase that mediates the modification of this residue. In addition, other snoRNAs also guide the formation of 2 -0-methylated nucleosides (-CH3) in rRNA. The pre-rRNA precursor is then cleaved at specific sites by RNases to produce a series of characteristic intermediates (41S), and finally mature rRNAs — 18S, 5.8S and 28S. During rRNA processing, the rRNA species must associate with more than 70 ribosomal proteins, as well as the 5S rRNA in the nucleolus, to form the small (S, 40S) and the large (L, 60S) ribosomal subunits, which are assembled and transported to the cytoplasm to initiate protein synthesis.")
56
A Myc protoonkogén által kódolt fehérje mindhárom RNS
polimeráz transzkripcióját serkenti, ezáltal a riboszóma bio- genezis mindhárom komponensének lértrejöttéhez hozzájárul UBF, upstream binding factor; SL-1, promoter selectivity factor; TIFIA, transcription intermediary factor IA; ERK, extracellular signal-regulated kinase; HAT, histone acetyltransferase.

57
A riboszóma biogenezis lépéseinek hibái tumorképződéshez vezethetnek
The regulation of ribosome biogenesis is a highly coordinated process that leads to accurate initiation and regulation of protein synthesis. There are three steps in this process, which, when deregulated, can contribute to increased tumorigenesis. The first step in ribosome production requires the synthesis of the 45S rRNA precursor. The transcription of this rRNA gene is negatively regulated by tumour suppressors such as p53 and retinoblastoma (RB) and augmented on mitogenic stimuli by several kinases that phosphorylate components of the transcription complex that are responsible for 45S synthesis. The accurate regulation of rRNA synthesis can be lost in cancer cells through inactivating mutations in tumour suppressors or upregulation of these kinases.Another step in ribosome biogenesis that maintains accurate cellular function involves the modification of rRNA. One type of modification, which is catalysed by an enzyme known as dyskerin, converts uridine into pseudouridine ( ). Mutations in the gene encoding dyskerin, DKC1, result in increased tumour susceptibility. Animal models that have lost DKC1 function show a marked increase in tumour incidence associated with a decrease in rRNA processing. Ribosome assembly, which involves the association of rRNA with more than 70 ribosomal proteins (made in the cytoplasm), is another important step. The rRNA and proteins are assembled into the large subunit (60S) and small subunit (40S) of the ribosome. An increase in ribosomal protein production and activity has been observed in many cancer types. Mutations in ribosomal proteins such as S19 have also been associated with a human syndrome that is characterized by increased tumour susceptibility. Each of the three highlighted steps (blue boxes) might have deleterious effects on the cell that could contribute to tumour initiation or cancer progression, or both, through aberrant regulation of protein synthesis. This can be manifested either by an increase in ribosome production, thereby leading to an upregulation in total translation, or by alterations in translation of specific mRNAs, which are involved in the regulation of cell proliferation. Furthermore, when key checkpoints in the cell that are important in coordinating ribosome production with accurate cell-cycle progression are lost, 'nucleolar stress' can result, and subsequently unrestrained cellular proliferation occurs. The advent of proteomics will aid in identifying protein targets that are deregulated as a result of perturbations in these pathways.
 and augmented on mitogenic stimuli by several kinases that phosphorylate components of the transcription complex that are responsible for 45S synthesis. The accurate regulation of rRNA synthesis can be lost in cancer cells through inactivating mutations in tumour suppressors or upregulation of these kinases.Another step in ribosome biogenesis that maintains accurate cellular function involves the modification of rRNA. One type of modification, which is catalysed by an enzyme known as dyskerin, converts uridine into pseudouridine ( ). Mutations in the gene encoding dyskerin, DKC1, result in increased tumour susceptibility. Animal models that have lost DKC1 function show a marked increase in tumour incidence associated with a decrease in rRNA processing. Ribosome assembly, which involves the association of rRNA with more than 70 ribosomal proteins (made in the cytoplasm), is another important step. The rRNA and proteins are assembled into the large subunit (60S) and small subunit (40S) of the ribosome. An increase in ribosomal protein production and activity has been observed in many cancer types. Mutations in ribosomal proteins such as S19 have also been associated with a human syndrome that is characterized by increased tumour susceptibility. Each of the three highlighted steps (blue boxes) might have deleterious effects on the cell that could contribute to tumour initiation or cancer progression, or both, through aberrant regulation of protein synthesis. This can be manifested either by an increase in ribosome production, thereby leading to an upregulation in total translation, or by alterations in translation of specific mRNAs, which are involved in the regulation of cell proliferation. Furthermore, when key checkpoints in the cell that are important in coordinating ribosome production with accurate cell-cycle progression are lost, nucleolar stress can result, and subsequently unrestrained cellular proliferation occurs. The advent of proteomics will aid in identifying protein targets that are deregulated as a result of perturbations in these pathways.")
58
A sejtciklus alatt számos fehérje akkumulálódik
a magvacskában PTHrP = parathyroid hormon related protein; PPIγ = protein phosphatase Iγ; CDC14A =

60
A p53 és a magvacska kapcsolata
HDM2(MDM2) = ubiquitin ligáz p53 = tumor szuppresszor p14ARF = tumor szuppresszor Normális sejt DNS károsodás és/vagy onkogén aktiválódás A p53 és a magvacska kapcsolata
 = ubiquitin ligáz. p53 = tumor szuppresszor. p14ARF = tumor szuppresszor. Normális sejt. DNS károsodás és/vagy. onkogén aktiválódás. A p53 és a magvacska kapcsolata.")
61
A TIF-IA jelenléte nélkülözhetetlen a sejtmagvacska
meglétéhez TIF-IA = transcription intermediary factor MDM2 = HDM2 = ubiquitin ligáz

62
A TIF IA génkiütés a magvacska szerkezetének megváltozásához vezet
vad génkiütött

63
A TIF IA génkiütött sejtek magvacskájából kiszabaduló
fehérjék a sejtmagban oszlanak el vad génkiütött

64
A RNS interferencia (RNAi) és a H3 hisztonmetiláció
befolyásolja sejtmagvacska számot

65
EPIGENETIKUS MECHANIZMUSOK
Transzkripciós Hiszton módosítások DNS metiláció Poszt-transzkripciós RNAi= RNS interferencia miR= mikro-RNS lincRNS = large intergenic non-coding RNS

66
RNS INTERFERENCIA (RNAi)
– GÉN INAKTIVÁCIÓ: SPECIFIKUS GÉNEK ÉS FELTEHETŐEN ENDOGÉN VÍRUSOK, TRANSZPOZONOK, RETROVÍRUSOK

67
POSZT-TRANSZKRIPCIONÁLIS
Indukáló dsRNS Új dsRNS Cél RNS vagy Széttekerés? siRNS DICER = dsRNS-függő endonukleáz RdRP = RNS-függő RNS polimeráz DICER RdRP Primer kapcsolódás RNS INTERFERENCIA/ POSZT-TRANSZKRIPCIONÁLIS GÉNCSENDESÍTÉS 1

68
POSZT-TRANSZKRIPCIONÁLIS (small interfering RNS)
RNS INTERFERENCIA/ POSZT-TRANSZKRIPCIONÁLIS GÉNCSENDESÍTÉS 2 mRNS degradáció Nincs fehérje expresszió homológ siRNS (small interfering RNS) Kétszálú RNS oligonukleotid 21-23 nt TT 3’ túlnyúló szál RISC (RNS induced silencing complex)
 Kétszálú RNS oligonukleotid nt. TT 3’ túlnyúló szál. RISC (RNS induced silencing complex)")
69
RNAi vad The cells were fixed and stained for tubulin, actin and DNA

70
A RNS interferencia (RNAi) és a H3 hisztonmetiláció
befolyásolja sejtmagvacska számot

71
A RNS interferencia (RNAi) és a H3 hisztonmetiláció
befolyásolja sejtmagvacska számot

72
A normal Drosophila cell nucleus (left) contains just one well-formed nucleolus, shown in pink. The cell nuclei of Su(var)3-9 mutants (center and right) exhibit multiple irregular nucleoli. Karpen and Peng studied fruit flies with mutant Su(var)3-9 genes, flies that could not produce HP1 proteins or the proteins necessary to methylate H3K9. The results were dramatic. Because the ribosomal DNA genes could not be silenced, rDNA proliferated. These and other repeated DNAs became dispersed and disorganized throughout the cell nucleus. Instead of a single well-formed nucleolus where genes for ribosomal DNA were normally located in the heterochromatin, multiple nucleoli appeared in the nucleus.
3-9 mutants (center and right) exhibit multiple irregular nucleoli. Karpen and Peng studied fruit flies with mutant Su(var)3-9 genes, flies that could not produce HP1 proteins or the proteins necessary to methylate H3K9. The results were dramatic. Because the ribosomal DNA genes could not be silenced, rDNA proliferated. These and other repeated DNAs became dispersed and disorganized throughout the cell nucleus. Instead of a single well-formed nucleolus where genes for ribosomal DNA were normally located in the heterochromatin, multiple nucleoli appeared in the nucleus..")
73
A nukleoszóma kapcsolata más betegségekkel

74
45,X0 karyotípus

75
Turner- szindróma 139,5 cm

76
Turner szindróma Pterygium colli

77
Turner- szindróma Hydrops fetalis

78
Az alacsony termet magyarázatául szolgáló hipotézis
RPS4 = kis riboszóma alegység proteint kódoló gén Az alacsony termet magyarázatául szolgáló hipotézis Ideograms of the X and Y chromosomes with the position of the lymphoedema critical region for both chromosomes indicated. The position of the pseudoautosomal boundary (PAB), the STS markers sY20 and sY45, the PAC clone dJ109C12, the Y chromosome genes SRY, ZFY, RPS4Y, TGIFLY, PCDHY, DBY, DFFRY, and SMCY, and their X chromosome homologues SOX3, ZFX, RPS4X, TGIFLX, PCDHX, DBX, DFFRX, and SMCX, the X chromosome genes VEGF-D, OTC, TALLA-1, NYX, CASK, GPR34, and IPP2H, and the pseudogenes NAPΨ and CASKΨ are also indicated.
, the STS markers sY20 and sY45, the PAC clone dJ109C12, the Y chromosome genes SRY, ZFY, RPS4Y, TGIFLY, PCDHY, DBY, DFFRY, and SMCY, and their X chromosome homologues SOX3, ZFX, RPS4X, TGIFLX, PCDHX, DBX, DFFRX, and SMCX, the X chromosome genes VEGF-D, OTC, TALLA-1, NYX, CASK, GPR34, and IPP2H, and the pseudogenes NAPΨ and CASKΨ are also indicated.")
80
Nucleostemin egy őssejt specifikus nukleáris komponens

81
A sejtmagvacska nucleosteminjének kimutatása
Cells expressing a nucleostemin–GFP fusion (green) and labelled with anti-B23 antibody (red) show localization of nucleostemin in the nucleolus B23 = nucleophosmin = numatrin = nucleolar phosphoprotein
 and labelled with anti-B23 antibody (red) show localization of nucleostemin in the nucleolus. B23 = nucleophosmin = numatrin = nucleolar phosphoprotein.")
Hasonló előadás





 DNS-ből,>")

 daganatkeltő.>")
 megváltozHAT Ebből visszakövetkeztethetünk a mutációt szenvedett gén funkciójára.>")












 kialakulása Genetikai, Sejt- és Immunbiológiai Intézet Falus András.>")

