Előadást letölteni
Az előadás letöltése folymat van. Kérjük, várjon

1
Az eukarióta sejtváz II.
Az aktin filamentumok és az intermedier filamentumok Darvas Zsuzsanna Ph.D. egyetemi docens GSI SE

2
Aktin filamentumok=mikrofilamentumok
mikrotubulusok: zöld aktin: piros migration.wordpress.com/.../

3
Baktérium: aktin homológ fehérje
www2.mrc-lmb.cam.ac.uk/SS/Lowe_J/ www2.mrc-lmb.cam.ac.uk/groups/jyl/frame_MreB.html

4
Aktin a baktériumsejtben
A baktérium sejt membránja alatt feladat: sejt alakja, sejt osztódása

5
aktin filamentumok szerveződése
Polymerizáció /depolimerizáció: polarizált struktúra

6
Aktin típusok: G aktin és F aktin
F aktin: 2 csavart szál poláris - és + vég polimerizáció depolimerizáció + - G aktin homodimerek ATP/ADP kötés Aktin típusok: G aktin és F aktin

7
F aktin és G aktin ciklusos kapcsolata és a taposómalom jelenség
Filamentous (F)-actin is, however, asymmetric and the two extremities retain different kinetic characteristics. Actin monomers assemble much more rapidly at the 'barbed end' compared to the 'pointed end' (these names correspond to the arrowhead appearance of myosin heads bound to actin filaments). The critical concentration of the pointed end is higher than that of the barbed end. When F-actin and G-actin are at equilibrium, the global critical concentration is intermediate between those of the two ends separately. So, at this stage, there is a net loss of molecules at the pointed end and a net addition at the barbed end. The two rates balance, which leads to treadmilling — a net flow of actin subunits through the filament.
-actin is, however, asymmetric and the two extremities retain different kinetic characteristics. Actin monomers assemble much more rapidly at the barbed end compared to the pointed end (these names correspond to the arrowhead appearance of myosin heads bound to actin filaments). The critical concentration of the pointed end is higher than that of the barbed end. When F-actin and G-actin are at equilibrium, the global critical concentration is intermediate between those of the two ends separately. So, at this stage, there is a net loss of molecules at the pointed end and a net addition at the barbed end. The two rates balance, which leads to treadmilling — a net flow of actin subunits through the filament.")
8
Növekvő aktin filamentumok
Aktin filamentum keletkezése in vitro Növekvő aktin filamentumok

9
Actin binding proteins (ABPs) – aktin kötő fehérjék
3 csoport polymerizáció/depolymerizáció, nukleáció - kötegelő és keresztkötő fehérjék - motor fehérjék: miozin

10
ABPs Aktin monomer: ATP és ADP kötött formája ATP kötött: beépül -profilin ADP kötött: leválik-cofilin ADP/ATP csere-CAP Nukleáció (aktin polimerizáció) fehérjéi: ARP komplex: polimerizáció és elágazás Formin: aktin polimerizáció és kontollált növekedés Spire: 4 aktin monomer összekapcsolása-új aktin filamentum indítása Keresztkötő és kötegelő fehérjék Laza és szoros kötegek –-aktinin, fimbrin Két és háromdimenziós hálózatok – filamin, spektrin Sapkaképző fehérjék + végen sapka, amely gátolja a polimerizációt: capZ Gelsolin: sapka és rövid filamentum képzés Reguláció: Foszforiláció/defoszforiláció RHo, Rac. Cdc42 kis G fehérjék
 fehérjéi: ARP komplex: polimerizáció és elágazás. Formin: aktin polimerizáció és kontollált növekedés. Spire: 4 aktin monomer összekapcsolása-új aktin filamentum indítása. Keresztkötő és kötegelő fehérjék. Laza és szoros kötegek –-aktinin, fimbrin. Két és háromdimenziós hálózatok – filamin, spektrin. Sapkaképző fehérjék. + végen sapka, amely gátolja a polimerizációt: capZ. Gelsolin: sapka és rövid filamentum képzés. Reguláció: Foszforiláció/defoszforiláció. RHo, Rac. Cdc42 kis G fehérjék.")
11
phalloidin Cytochalasin B gomba metabolitok
Gomba: Amanita phalloides gomba metabolitok cytochalasin – gátolja az aktin polymerizációt phalloidin – stabilizálja az F aktin filamentumokat commons.wikimedia.org/wiki/Image:Cytochalasin... Cytochalasin B Gomba: Helminthosporium dermatioideum

12
Motor fehérjék: miozinok
Szerkezet: nehéz lánc és könnyű láncok globuláris fej és spirális farok fej: motor domén ATP-áz aktivitás Mozás iránya: általában - vég motor

13
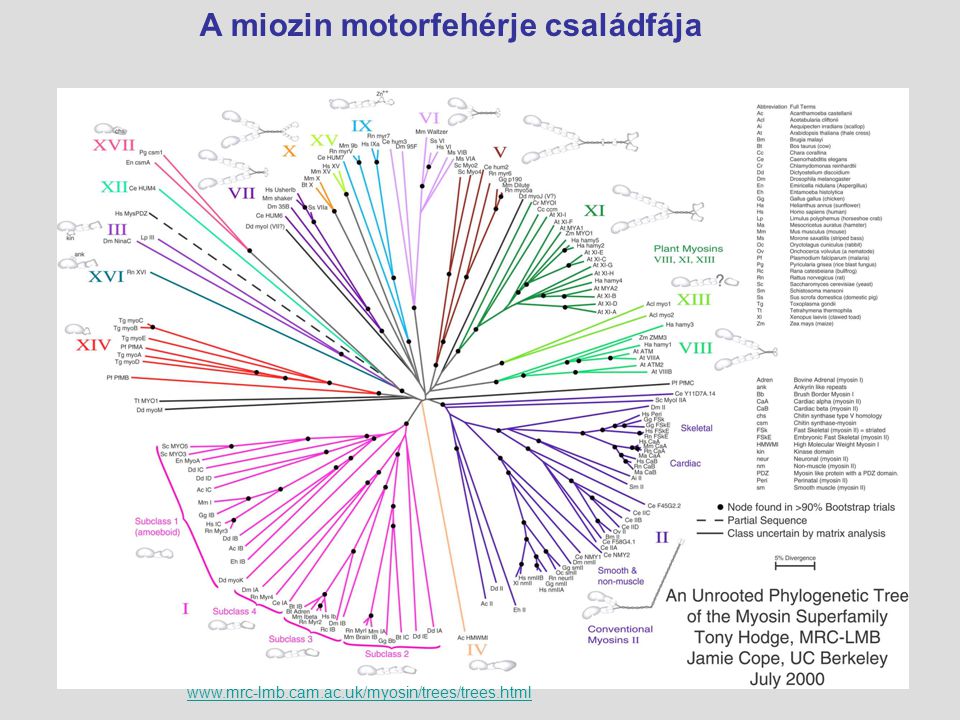
A miozin motorfehérje családfája
14
Aktin filamentumok szerveződése I.
ABPs Aktin ( piros): kérgi aktin réteg a plazmamembrán alatt
: kérgi aktin réteg a plazmamembrán alatt.")
15
kirschner.med.harvard.edu/.../andres.shtml
Molekulák a kérgi aktin hálózat létrehozásában Regulation of actin assembly by the WAVE complex Background Regulation of the actin cytoskeleton is crucial for cell morphology and motility during development, health and disease. In motile cells, actin assembly occurs largely by nucleation of new actin filaments on the sides of existing filaments to generate branched filament networks that push and deform the plasma membrane, causing protrusion of the leading edge. The Arp2/3 complex is a key factor mediating nucleation and branching of filaments, and it is regulated by nucleation promoting factors such as WASP, N-WASP and WAVE proteins. Upstream signals, initiated by ligands binding to growth factor or chemoattractant receptors, recruit nucleation promoting factors to the plasma membrane, where they stimulate actin nucleation by the Arp2/3 compex. kis G fehérjék (Rho, Rac, Cdc42) szerepe szignál továbbítás az aktin filamentumok nukleációjához) kirschner.med.harvard.edu/.../andres.shtml
 szerepe szignál továbbítás az aktin filamentumok nukleációjához) kirschner.med.harvard.edu/.../andres.shtml.")
16
1 2 Aktin filamentumok szerveződése II. lamellipódium:
2 ill. 3 dimenziós aktin hálózat a membrán alatt keresztkötő aktin fehérjék pl. filamin 2. filopódium: párhuzamos aktin kötegek - kötegelő fehérjék pl. fimbrin

17
Actin filaments polymerize at their fast-growing 'barbed' ends near the plasma membrane. They stop growing when they are capped by capping protein. If barbed-end capping activity is high, filaments get capped, stop growing and remain short. This favours an actin filament network that is most efficient for lamellipodium expansion. Filopodia result when filaments of the lamellipodial actin network are recruited by a filopodial tip complex that includes Ena/VASP proteins; filaments grow long because Ena/VASP proteins also inhibit capping locally.

18
Aktin filamentumok szerveződése III.
Mikrobolyhok (villin, fimbrin- kötegelő fehérjék miozin I. – motor fehérje) anatomy.iupui.edu/.../cell.f04/cellf04.html Tövénél: (spektrin, miozin, intermedier filamentumok) terminal web (végháló)
 anatomy.iupui.edu/.../cell.f04/cellf04.html. Tövénél: (spektrin, miozin, intermedier filamentumok) terminal web (végháló)")
19
Aktin filamentumok szerveződése IV.
scienceblogs.com Aktin filamentum kötegek a citoplazmában: stressz rostok szerep az amőboid típusú mozgásban

20
Mozgás: sejtváz komponensek és motor
fehérjék segítségével Két alaptípus: Tubulin alapú mozgás: csilló, ostor Aktin alapú mozgás : amőboid, izom

21
Amőboid mozgás cellix.imba.oeaw.ac.at/.../

22
1. Álláb (vagy filopódium, lamellipódium) képződés a vezető végen : aktin filamentumok képződése – aktin polimerizáció, elágazás, amelyet kis G fehérjék szabályoznak (Rac, Rho, cdc42) 2. Sejt-mátrix kölcsönhatás: fokális kontaktusok képződése ebben:sejt-mátrix kapcsoló molekulák integrinek és ECM molekulák szerepe 3. proteolízis: fokális kontaktus területén ECM molekulák bontása MMP (matrix metallo proteináz) enzimek szerepe 4. Aktomiozin kontrakció (transzlokáció) : a fokális kontaktushoz kihorgonyzott aktin filamentumok (stressz rostok) közé miozin filametumok szerveződnek és az aktin filamentumok elcsúszva húzzák a sejtet a vezető vég irányába (miozin foszforiláció: ATP-áz aktivitás) 5.Leválás: a sejt hátsó végén a fokális kontaktusok megszüntetése, és az aktin filamentumok átrendeződése
 enzimek szerepe. 4. Aktomiozin kontrakció (transzlokáció) : a fokális kontaktushoz kihorgonyzott aktin filamentumok (stressz rostok) közé miozin filametumok szerveződnek és az aktin filamentumok elcsúszva húzzák a sejtet a vezető vég irányába (miozin foszforiláció: ATP-áz aktivitás) 5.Leválás: a sejt hátsó végén a fokális kontaktusok megszüntetése, és az aktin filamentumok átrendeződése.")
23
Fokális kontaktus szerkezete
Aktin filamentumok: stress rostok Kötegelő fehérje + miozin Kapcsoló fehérjék integrinek ECM

24
Striamoeba

25
Aktin filamentumok szerveződése: izomsejt
Izomrostok Miofibrillumok Miofilamentumok szarkomer Aktin - vékony filamentum Miozin – vastag filamentum

26
Vázizom EM-os kép homepage.smc.edu/.../anatomy1/1musphys.html

27
Vékony filamentum szerkezete
Aktin filamentumok és ABP -ek Tropomyozin és troponin komplex Vékony filamentum szerkezete Tropomyozin: dupla alfa hélix fenntartja a filamentum helyzetét aktin más fehérje kötődést gátolja Troponin: Tn-T tropomyozin kötés Tn-C Ca2+(4 Ca2+/mol = calmodulin) kötés Tn-I inhibitor
 kötés. Tn-I inhibitor.")
28
Vastag filamentum szerkezete I.
Filamentum kialakításában: Myosin binding proteins pl. MyBP C, és más fehérjék pl. M protein, myomesin) fej Könnyű láncok MLC Nehéz láncok MHC „hinge” 134 nm miozin fejek Farki részek Bipoláris filamentum
 fej. Könnyű láncok MLC. Nehéz láncok MHC. „hinge 134 nm. miozin fejek. Farki részek. Bipoláris filamentum.")
29
Vastag filamentum térbeli szerkezete II.
gilead.org.il/hcm/ Miozin kötő fehérje: MyBPC elhelyezkedése

30
Aktin filamentumok szerveződése: szarkomer
Actin (green), myosin (red). Rod-like tropomyosin molecules (black lines). Thin filaments in muscle sarcomeres are anchored at the Z-disk by the cross-linking protein α -actinin (gold) and are capped by CapZ (pink squares). The thin-filament pointed ends terminate within the A band, are capped by tropomodulin (bright red). Myosin-binding-protein C (MyBP-C; yellow transverse lines), (Gregorio et al. 2000). A haráncsíkolt izom és a szívizom müködési egysége a szarkomer
, myosin (red). Rod-like tropomyosin molecules (black lines). Thin filaments in muscle sarcomeres are anchored at the Z-disk by the cross-linking protein α -actinin (gold) and are capped by CapZ (pink squares). The thin-filament pointed ends terminate within the A band, are capped by tropomodulin (bright red). Myosin-binding-protein C (MyBP-C; yellow transverse lines), (Gregorio et al. 2000). A haráncsíkolt izom és a szívizom müködési egysége a szarkomer.")
31
Az izommüködés mechanizmusa: „sliding”: elcsúszás
scienceblogs.com vékony filamentumok a vastag filamentumok közé csúsznak szarkomer rövidül (~ 7 nm)
")
32
Az elcsúszási mechanizmus

33
Miozin fej ATP hiányában aktinhoz kötődik
Az elcsúszás menete: Miozin fej ATP hiányában aktinhoz kötődik Miozin fej ATP-t köt és lazán vagy nem kötődik az aktinhoz Miozin ATP-t bont, és a fej konformációja megváltozik A foszfát csoport leválása közben a miozin fej erősen kötődik az aktinhoz ADP leválása közben a fej konformációja megváltozik és elhúzza az aktin filamentumot accessexcellence.org/RC/VL/GG/ecb/myosin_...

34
intro.bio.umb.edu/.../muscle/actomyosin.html

35
Szarkoplazmás retikulum szerepe

36
citokinézis

37
Kontraktilis gyűrű az osztódó sejtekben
straightlab.stanford.edu/cytokinesis.html /ywwy/zbsw(E)/edetail11.htm
/edetail11.htm.")
38
X-hez kötött recesszív öröklődés
Duschenne izomdisztrófia Duschenne izomdisztrófia X-hez kötött recesszív öröklődés Duschenne izomdisztrófia és Becker izomdisztrófia

39
Disztrofin fehérje kimutatása és a DMD/BDM öröklődése
X-hez kötött recesszív öröklődésmenet

40
Disztrofin: egyik kulcsmolekula az ECM és a sejtváz között
Aktin kötőhely-piros Β-dystroglycan kötőhely-zöld Syntrophin kötőhely - sárga jennyndesign.com/DMD/physiology5.html

41
in frame (Becker) - frame shift (Duschenne) mutáció
Disztrofin gén ghr.nlm.nih.gov/gene=dmd Allél heterogénia in frame (Becker) - frame shift (Duschenne) mutáció compbio.berkeley.edu/.../ed/rust/Dystrophin.html
 - frame shift (Duschenne) mutáció. compbio.berkeley.edu/.../ed/rust/Dystrophin.html.")
42
Duchenne és Becker izomdisztrófia
Duchenne muscular distrophy Becker muscular distrophy Disztrofin gén: 79 exonból áll – génen belül forró pontok – mutációs helyek többféle klinikai betegség – eltérő mutációk miatt

44
A sejtváz komponensei: Intermedier filamentumok
Fő feladatok - sejt alakjának stabilizálása mechanikai támaszték szöveti integritás (sejtek sejtváza a sejtkapcsoló struktúrákkal az ECM-hoz kapcsolódik és így a sejtek egymással is egységet alkotnak) Aktin filamentum
 Aktin filamentum.")
45
Intermedier filamentumok
Xenopus oligodendrocytes, immunostained with antibodies to the intermediate filament protein GFAP (green). Nuclei labelled with DAPI (blue). GFAP (glial fibrillary acidic protein) is a commonly used marker protein for the identification of glial cells. web.uct.ac.za/.../research/neurosci2.htm migration.wordpress.com/.../
. Nuclei labelled with DAPI (blue). GFAP (glial fibrillary acidic protein) is a commonly used marker protein for the identification of glial cells. web.uct.ac.za/.../research/neurosci2.htm. migration.wordpress.com/.../")
46
Baktérium: intermedier filamentum homológ: Crescentin : creS
Citokeratin 19-el lamin A-val homológ Feladat: sejt alak Caulobacter crescentus thefutureofthings.com/articles.php?itemId=31/57/

47
Eukarióta sejtekben 2 nagy IF rendszer
sejtmagban laminokból amelyek létrehozzák a nukleáris laminát és az intranukleáris filamentumokat citoplazmában - eltérő fehérjékből amelyek kapcsolatot teremtenek az ECM és a külső magmembrán között IF –t 65 gén kódolja emberben nagy géncsalád

48
Intermedier Filamentum IF
TEM TEM MT és IF kapcsolat Dr. Alan R. Prescott University of Dundee CHIPs, School of Life Sciences Dundee, UK Cells showing small heat shock protein localized to intermediate filaments (100x)
")
49
Sejtvándorlás, sejtmozgás
a. Mechanikai támasz Sejtszerkezet stabilizálása Szignál továbbítás és modulálás

50
Intermedier filamentumok csoportosítása
5 osztályuk van (SHC=sequence homology classes) aminósav szekvencia homológia alapján webpathology.com/image.asp?n=7&Case=38 cytokeratin bázikus keratin (II típus) (CK1, CK2, CK3, CK4, CK5, CK6, CK7, CK8 and CK9.) savas keratin (I típus) (CK10, CK12, CK 13, CK14, CK16, CK17, CK18, CK19,CK20) 3. vimentin, desmin, GFAP (glial fibrillary acid protein) 4. neurofilamentum proteinek (NF M, NF H) 5. laminok desmin
 aminósav szekvencia homológia alapján. webpathology.com/image.asp n=7&Case=38. cytokeratin. bázikus keratin (II típus) (CK1, CK2, CK3, CK4, CK5, CK6, CK7, CK8 and CK9.) savas keratin (I típus) (CK10, CK12, CK 13, CK14, CK16, CK17, CK18, CK19,CK20) 3. vimentin, desmin, GFAP. (glial fibrillary acid protein) 4. neurofilamentum proteinek (NF M, NF H) 5. laminok. desmin.")
51
IF csoportjai és lokalizációja
Intermediate-filament proteins (IF in the figure) have been classified into five distinct types on the basis of their sequence identity and tissue distribution (see figure)5. The type-I and -II sequence-homology groups are the keratins of epithelial cells. These form obligate heterodimers that consist of one of each type of keratin (one acidic and one basic) and are categorized under assembly group-1. Type-III intermediate filaments include vimentin, desmin, glial fibrillary acidic protein (GFAP), synemin and peripherin. The neurofilament (NF) triplet proteins — NF-L, NF-M and NF-H — -internexin, syncoilin and nestin comprise the type-IV intermediate filaments. The type-III and type-IV intermediate filaments can form heterodimers with each other and are classified as assembly group-2. Nuclear lamin A and its splice variant, lamin C, together with lamin B1 and B2 are type-V intermediate filaments. Because lamins do not seem to co-polymerize with other types of intermediate filament, they have been assigned to assembly group-3. All intermediate-filament proteins have a characteristic tripartite structure, consisting of a highly -helical central rod domain that is flanked by non- -helical head and tail domains. The rod domain consists of the heptad repeats that are the signature of -helical proteins. The heptads are interrupted by short linker sequences (L1, L12 and L2), which results in four -helical segments: 1A and 1B comprise 'coil 1', and 2A and 2B form 'coil 2'. The structure and length of the central rod domain is highly conserved in vertebrate intermediate-filament proteins, with the exception of the nuclear lamins, which contain six extra heptads in the 1B segment. The longer 1B segment is thought to be responsible for the inability of lamins to co-polymerize with other intermediate-filament proteins. A four-residue insertion in segment 2B that produces a discontinuity in the heptad-repeat pattern ('stutter') is a feature that is highly conserved in all intermediate-filament proteins. The variability of intermediate-filament proteins lies in the length and sequence of the head and tail domains (for example, the long nestin tail or the immunoglobulin fold in the lamin tail domain) that are thought to be involved in regulating the interactions between intermediate filaments and other proteins.
 have been classified into five distinct types on the basis of their sequence identity and tissue distribution (see figure)5. The type-I and -II sequence-homology groups are the keratins of epithelial cells. These form obligate heterodimers that consist of one of each type of keratin (one acidic and one basic) and are categorized under assembly group-1. Type-III intermediate filaments include vimentin, desmin, glial fibrillary acidic protein (GFAP), synemin and peripherin. The neurofilament (NF) triplet proteins — NF-L, NF-M and NF-H — -internexin, syncoilin and nestin comprise the type-IV intermediate filaments. The type-III and type-IV intermediate filaments can form heterodimers with each other and are classified as assembly group-2. Nuclear lamin A and its splice variant, lamin C, together with lamin B1 and B2 are type-V intermediate filaments. Because lamins do not seem to co-polymerize with other types of intermediate filament, they have been assigned to assembly group-3. All intermediate-filament proteins have a characteristic tripartite structure, consisting of a highly -helical central rod domain that is flanked by non- -helical head and tail domains. The rod domain consists of the heptad repeats that are the signature of -helical proteins. The heptads are interrupted by short linker sequences (L1, L12 and L2), which results in four -helical segments: 1A and 1B comprise coil 1 , and 2A and 2B form coil 2 . The structure and length of the central rod domain is highly conserved in vertebrate intermediate-filament proteins, with the exception of the nuclear lamins, which contain six extra heptads in the 1B segment. The longer 1B segment is thought to be responsible for the inability of lamins to co-polymerize with other intermediate-filament proteins. A four-residue insertion in segment 2B that produces a discontinuity in the heptad-repeat pattern ( stutter ) is a feature that is highly conserved in all intermediate-filament proteins. The variability of intermediate-filament proteins lies in the length and sequence of the head and tail domains (for example, the long nestin tail or the immunoglobulin fold in the lamin tail domain) that are thought to be involved in regulating the interactions between intermediate filaments and other proteins.")
52
IF szerkezete Közös motívum: 45 nm-es hosszúságú -helikális un. „rod domain” + nem -helikális N terminális fej (head) – és C terminális farok (tail) domén
 – és C terminális farok (tail) domén.")
53
? Intermedier filamentum szerveződése
(Lodish, H. et al. Mol. Cell Biol. 2000, 767)
")
54
Mikrotubulusok szerepe az intermedier filamentumok kialakulásában
Particles move along microtubules (MT in the figure) bidirectionally in association with the molecular motors kinesin and dynein/dynactin. b | Particles, which are aggregates of intermediate-filament (IF in the figure) precursor structures/subunits, give rise to squiggles (short filaments). c | Squiggles, in turn, fuse with other squiggles, end-to-end, to form longer filaments. d | Squiggles and long intermediate filaments also move along microtubules in the same way as particles. e | Longer intermediate filaments are formed, which can remain connected to microtubules by motors such as dynein/dynactin.
 bidirectionally in association with the molecular motors kinesin and dynein/dynactin. b | Particles, which are aggregates of intermediate-filament (IF in the figure) precursor structures/subunits, give rise to squiggles (short filaments). c | Squiggles, in turn, fuse with other squiggles, end-to-end, to form longer filaments. d | Squiggles and long intermediate filaments also move along microtubules in the same way as particles. e | Longer intermediate filaments are formed, which can remain connected to microtubules by motors such as dynein/dynactin.")
55
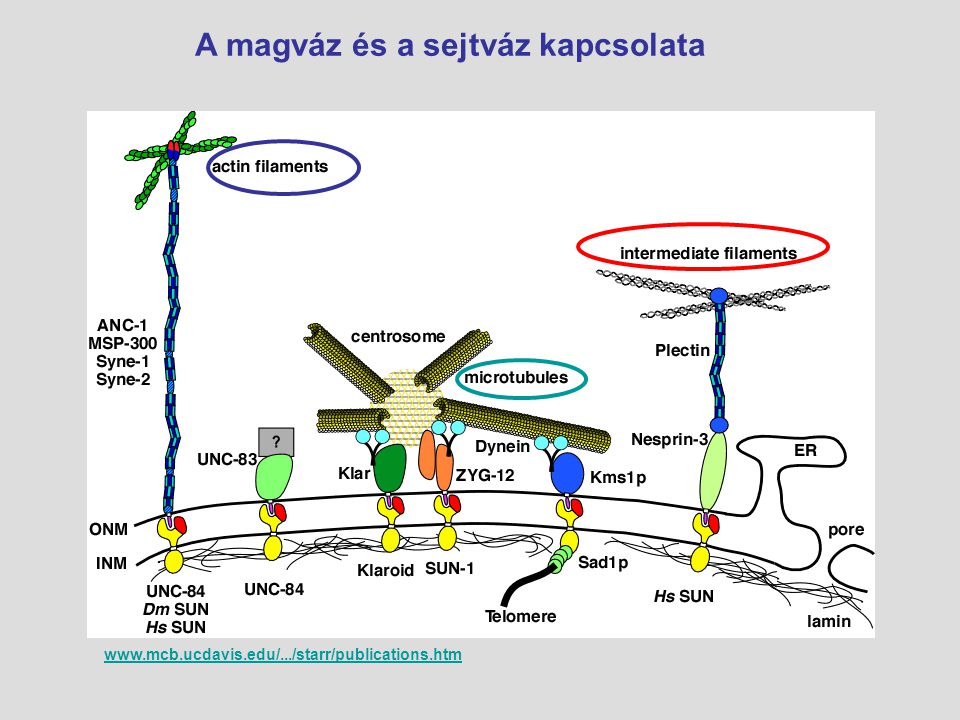
A magváz és a sejtváz kapcsolata
56
A magváz, a sejtváz és az ECM kapcsolata – a sejtek egységes hálózatot alkotnak

57
Intermedier filamentumok kimutatása tumorokban
Intermedier filamentumok mutációihoz kötődő betegségek

58
IF összetétel jellemző a tumorokra – IF festés a tumorazonosítás lehetősége biocare.net/clone/ks20-8/ Cytokeratin 20 is a unique type I keratin that is expressed in adenocarcinomas of the colon, stomach, pancreas and bile system. It is also expressed in mucinous ovarian tumors, transitional cell carcinomas of the urinary tract, and Merkel cell carcinomas. Tumor cells are positive for Vimentin.

59
Citokeratin mutáció okozta betegség
12 kromoszóma:KRT 5 gén kromoszóma:KRT 14 gén magas expresszió az epidermis alsóbb rétegeiben Mutációk: epidermolysis bullosa simplex (több mint 60 ismert mutáció)
")
60
Epidermolysis bullosa simplex
A sejtek közötti hálózat megszakad, s sejtek felső rétegei kis mechanikai stressz hatására vérzés kiséretében leválnak az alsóbb rétegekről

61
Epidermolysis Bullosa simplex AD
Epidermolysis Bullosa simplex AD *

Hasonló előadás
















>")




