Előadást letölteni
Az előadás letöltése folymat van. Kérjük, várjon

1
KROMOSZÓMA SZINTŰ HIBÁK KIMUTATÁSA, MÓDSZEREK
Előadó: Dr. Kocsis Zsuzsanna Országos Kémiai Biztonsági Intézet Molekuláris és Sejtbiológiai Osztály Budapest, Nagyvárad tér 2. Helyszín: ELTE,

2
Kromatinalapszerkezet: nukleoszóma
Kompartmentalizáció Prokarióta sejtek Eukarióta sejtek Kromatinalapszerkezet: nukleoszóma * DNS kettős spirál és a hisztonok alkotják * Hisztonok: bázikus fehérjék (argininben és lizinben gazdag) * 5 osztályuk van: H1H1, H2A, H2B, H3 és H4 nukleoszomális hisztonok Hisztonkorong (oktamer): 8 hisztonmolekulából álló (2*4) 2 csavarulatban 146 bázispárnyi DNS tekeredik rá 2 korong között kb. 60 bp linkerrégió + H1 molekula A DNS kondenzációját a kondenzin foszforilációja indítja el. kohezin ATP-áz
 * 5 osztályuk van: H1H1, H2A, H2B, H3 és H4. nukleoszomális hisztonok. Hisztonkorong (oktamer): 8 hisztonmolekulából álló (2*4) 2 csavarulatban. 146 bázispárnyi DNS tekeredik rá. 2 korong között kb. 60 bp linkerrégió + H1 molekula. A DNS kondenzációját. a kondenzin foszforilációja. indítja el. kohezin ATP-áz.")
3
Kromoszóma morfológia
Eukromatikus régió: aktív géneket tartalmaz (génexpresszió, RNS szintézis) Heterokromatikus régió: inaktív DNS szakaszok konstitutív heterokromatin (pl. centromer) fakultatív heteroktromatikus régió rövid kar centroméra hosszú kar testvérkromatidák
 Heterokromatikus régió: inaktív DNS szakaszok. konstitutív heterokromatin (pl. centromer) fakultatív heteroktromatikus régió. rövid kar. centroméra. hosszú kar. testvérkromatidák.")
4
Kromoszóma territórium
Egy adott kromoszóma a sejtmag egy adott régiójában található. Szigorú sejtmagi rend.

5
G-sávos normál női karyotípus
Fajok ecetmuslica rozs galamb egér patkány ember mezei nyúl kutya ponty páfrány Kromoszómaszám 8 14 16 40 42 46 48 78 104 1200 Humán lymphocyta kromoszóma preparátum Genom: a sejtmagban található összes genetikai információ. Kromoszóma: A genetikai információt tároló strukturális egység. Kromoszóma száma, morfológiája szervezettsége fajra jellemző és állandó. Humán kromoszóma Gének 1 2 968 13 748 18 766 21 303 X 1184 (Barr test) Y 231 (mikrodeléció meddőség) Nemhez kötött öröklődés G-sávos normál női karyotípus
 Y. 231 (mikrodeléció meddőség) Nemhez kötött öröklődés. G-sávos normál női karyotípus.")
6
Öregedés, rák, stabil kromoszóma
Telomer: - kromoszóma vége - rövid TTAGGG szakasz több ezerszeres mennyiségben (20-25 ezer bázispár) genetikai óra, telomer rövidülés öregedés 2009 orvosi Nobel Díj - minden osztódáskor 100 bp rövidülés a DNS polimeráz enzim működéséből adódóan - egészséges sejtekben a telomeraz inaktív - Az emberi sejt 50 osztódásra képes. 50 osztódás után a sejt apoptózissal meghal. Tumoros sejtekben a telomer szerepe: a telomeráz enzim aktivitása magasabb Azt gondolják, hogy a telomeráz enzim retrovírus eredetű. Ivarsejtekben is!
 genetikai óra, telomer rövidülés öregedés orvosi Nobel Díj. - minden osztódáskor 100 bp rövidülés a DNS polimeráz enzim. működéséből adódóan. - egészséges sejtekben a telomeraz inaktív. - Az emberi sejt 50 osztódásra képes. 50 osztódás után a sejt apoptózissal meghal. Tumoros sejtekben a telomer szerepe: a telomeráz enzim aktivitása magasabb. Azt gondolják, hogy a telomeráz enzim retrovírus eredetű. Ivarsejtekben is!")
7
MITOZIS * * * A mitózis gondoskodik a szülői és az utódsejt azonos kromoszómakészletéről. A sejtosztódás S fázisában a kromoszómák anyaga megkettőződik. A mitózis kromoszóma számtartó osztódás.

8
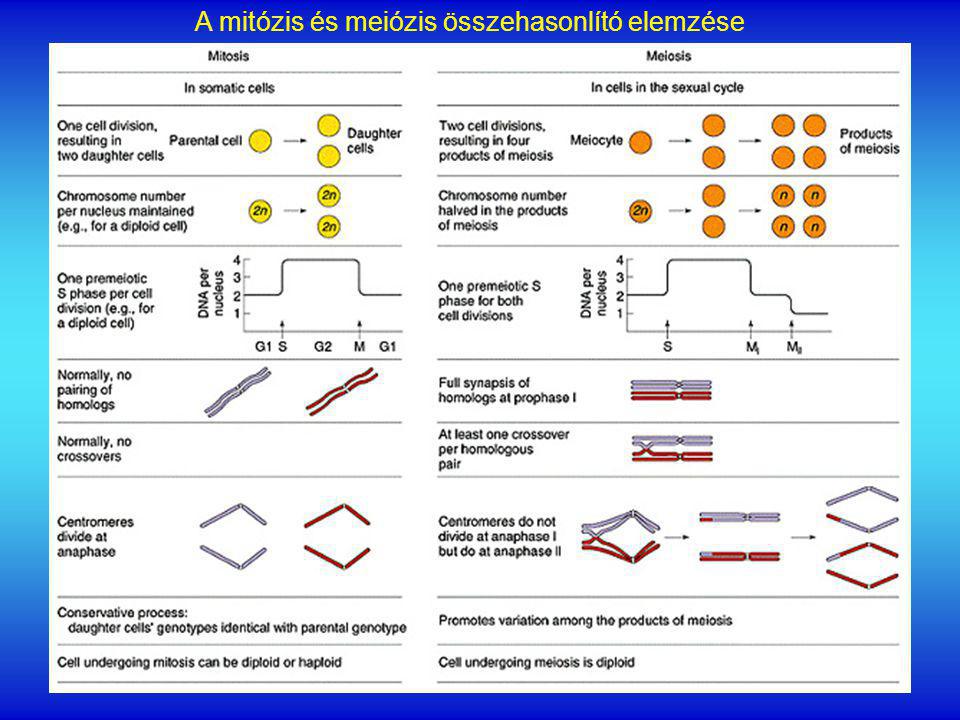
A mitózis és meiózis összehasonlító elemzése
9
X-kromoszóma inaktiváció A nőstény emlősök sejtjeiben
Az egyik X-kromoszóma inaktiválódik (random módon), XfXv . Az inaktivált X kondenzálódik - Barr-testecske. Bizonyos sejtek utódai együtt maradnak (sejtklónok). A nőstény emlősök teste mozaikos, vagyis olyan sejtek klónjainak a keveréke, amelyekben hol egyik, hol a másik X aktív. Heterozigótákon detektálhatók lehetnek a foltok. Vörös/fekete tarka macskák esete
, XfXv . Az inaktivált X kondenzálódik - Barr-testecske. Bizonyos sejtek utódai együtt maradnak (sejtklónok). A nőstény emlősök teste mozaikos, vagyis olyan sejtek. klónjainak a keveréke, amelyekben hol egyik, hol a másik X aktív. Heterozigótákon. detektálhatók lehetnek a foltok. Vörös/fekete tarka macskák esete.")
10
Hemofilia A B Nemhez kötött recesszív megbetegedés VIII. faktor
IX. faktor A VIII. faktor Hemofilia Nemhez kötött recesszív megbetegedés

11
KROMOSZÓMA ABERRÁCIÓ TÍPUSAI:
Strukturális kromoszóma aberráció: deléció inszerció transzlokáció Numerikus kromoszóma aberráció: poliploidia aneuploidia

12
kromoszóma szegmentek elvesztése
Deléció kromoszóma szegmentek elvesztése Különböző humán betegségek delécióra vezethetőek vissza. A kis deléciók tolerálhatóak. A nagy deléciók nem tolerálhatóak, letalitáshoz vezetnek.

13
Cri du chat – macskasírás szindróma
Az ember esetében a genom kiegyensúlyozatlanság miatt a legkisebb deléciók is komoly abnormalitást okoznak. A macskasírás szindróma estében az 5. kromoszóma rövid (p) karjának vége hiányzik. Mikroencefáliával, holdszerű arccal és szellemi elmaradottsággal jár. 4 év alatti halál. születéskori gyakoriság: 1/50,000
 karjának vége hiányzik. Mikroencefáliával, holdszerű arccal és szellemi elmaradottsággal jár. 4 év alatti halál. születéskori gyakoriság: 1/50,000.")
14
Rákos sejtek gyakran mutatnak deléciókat

15
DUPLIKÁCIÓ: Kromoszóma szegmensek megkettőződése.
Jó példa a duplikációra a Drosophila Bar mutációja.

16
Az X kromoszóma 16A régiójának kópia száma különböző

17
A gén duplikáció evolúciós szerepe
Ha a gén fontos a szervezet számára nem változhat. De ha a génből több kópia van a képződő proteinek módosulhatnak és új funkciókat láthatnak el. Duplikációval keletkezett gén családok hasonló proteineket készítenek. Jó példa erre a globin gének, amelyekről α és β globin láncok szintetizálódnak, a hemoglobin szerkezeti alegységei.

18
A gén duplikáció lehetőséget ad, a mutációs változásoknak, a funkciók divergálásának
Az emberi hemoglobin gén duplikációs változások eredménye. Különböző életkorokban különböző alegységek alakítják ki a működő hemoglobin molekulát. 3 hónapos korig az embrionális Hemg. Szülésig a magzati Hemg % Szülés után 2α 2ß.

19
Inverzió: Egy kromoszóma szakasz 180 fokos megfordulásának
az eredménye. pericentrikus inverzió magába foglalja centromert paracentrikus inverzió a centromert nem érinti, csak a kromoszóma egyik v. másik karját Gén környezet, gén kapcsoltság, génátírás módosul.

20
intrakromoszómális transzlokáció: Egy kromoszóma szakasz áthelyeződése
ugyanabba a kromoszómába. interkromoszómális transzlokáció: Nem homológ kromoszómák közötti transzlokáció. Transzlokáció során nincs genetikai anyag vesztés. A gének pozíciója azonban megváltozik.

21
14 és 21-es kromoszóma közötti Robertson transzlokáció.
Down szindróma: 14 és 21-es kromoszóma közötti Robertson transzlokáció. A 21-es krom. transzlokációja a 14-es krom.-ra eredményezi a familiáris Down syndromát.Örökölhető. 21-es kromoszóma triszómia (Osztódási hiba) Az anya életkorának előrehaladtával (40-45 év felett) az előfordulási gyakoriság exp. növekszik. Prenatális szűrés. Születéskori gyakorisága nagy: 1/500-1/700
 Az anya életkorának előrehaladtával (40-45 év felett) az előfordulási gyakoriság exp. növekszik. Prenatális szűrés. Születéskori gyakorisága nagy: 1/500-1/700.")
22
Myeloid leukémia A 9-es és a 22-es kromoszómák közötti transzlokáció eredménye a “Philadelphia kromoszóma” Az esetek 90%-ban krónikus myeloid leukemiát okoz.

23
Burkitt’s lymphoma Az esetek 90% esetén a 8-as és 14-es kromoszómák közötti transzlokáció következménye. Transzlokáció következtében kialakuló pozíció effektus hatására kialakuló onkogének, sejtosztódást, rák keletkezését okozhatják.

24
Alternatív toxikológiai módszerek
in vitro vizsgálatok Ajánlott teszt rendszerek: Egysejtűeken végzett tesztek Rovartesztek in vitro sejtkultúrákon végzett tesztek in vivo mutagenitási tesztek long term karcinogenitási állatkísérletek humán epidemiológiai vizsgálatok Genotoxikus és daganatkeltő hatások vizsgálata előnyei: in vitro körülmények között sejteken (élő állat alkalmazása nélkül ) rövid idejű olcsóbb reprodukálható nem használ élő állatot Az esetleges hatás megállapításához több, különböző módszerrel végzett vizsgálat egybehangzó eredményére van szükség.
 rövid idejű. olcsóbb. reprodukálható. nem használ élő állatot. Az esetleges hatás megállapításához több, különböző módszerrel végzett vizsgálat. egybehangzó eredményére van szükség.")
25
Bevezetés A BIZOTTSÁG 440/2008/EK RENDELETE a vegyi anyagok regisztrálásáról,értékeléséről,engedélyezéséről és korlátozásáról (REACH) szóló a 1907/2006/EK európai parlamenti és tanácsi rendelet értelmében alkalmazandó vizsgálati módszerek megállapításáról. B.rész.:Módszerek a toxicitás és egyéb egészségügyi hatások meghatározására B.10. Mutagenitás-Kromoszóma-Rendellenesség in vitro vizsgálata emlősökön OECD 473 TG OECD 487 TG in vitro mikronukleusz vizsgálat emlős sejteken B.13/14. Mutagenitás: Reverz mutagenitási vizsgálat baktériumokkal OECD 471 TG B.18. DNS károsodás és –reparáció-nem tervezett DNS-szintészis (unscheduled DNA Synthesis, UDS) –emlős sejteken in vitro OECD 476 TG B.19. In vitro emlőssejt testvér-kromatid kicserélődés (sister chromatid exchange , SCE) vizsgálat OECD 479 TG B. 20. Nemhez kötött recesszív letális vizsgálat Drosophila melanogasteren OECD 477 TG
 –emlős sejteken in vitro. OECD 476 TG. B.19. In vitro emlőssejt testvér-kromatid kicserélődés. (sister chromatid exchange , SCE) vizsgálat. OECD 479 TG. B. 20. Nemhez kötött recesszív letális vizsgálat Drosophila melanogasteren. OECD 477 TG.")
26
Azonos szakmai szabályok
GLP= Good Laboratory Practice (Helyes Laboratóriumi Gyakorlat) ISO= International Organization for Standardization (Nemzetközi Szabványügyi Szervezet) Vizsgálati irányelvek OECD= Organisation for Economical Cooperation and Development (Gazdasági es Fejlesztési Együttműködési Szervezet) Kromoszóma aberráció in vitro vizsgálata emlős sejteken OECD TG 473 vizsgálati irányelv szerint Célja: azon kémiai anyagok meghatározása, amelyek kromoszóma károsodást okoznak emlős sejtekben.
 ISO= International Organization for Standardization. (Nemzetközi Szabványügyi Szervezet) Vizsgálati irányelvek. OECD= Organisation for Economical Cooperation and Development. (Gazdasági es Fejlesztési Együttműködési Szervezet) Kromoszóma aberráció in vitro vizsgálata emlős sejteken. OECD TG 473 vizsgálati irányelv szerint. Célja: azon kémiai anyagok meghatározása, amelyek kromoszóma károsodást. okoznak emlős sejtekben.")
27
A kromoszóma aberráció vizsgálat ismertetése Sejtek
Alkalmazható sejtvonalak jellemzői: stabil permanens v. primer sejtkultúrák (pl. CHO, humán lymfocita) jó növekedési képesség, rövid generációs idő, kariotípus stabilitás, állandó krom. szám. Kromoszómák alaki változatossága és stabilitása. kínai hörcsög ovárium fibroblaszt sejt (CHO, Puck1957) Tápfolyadékok, tenyésztési körülmények Mesterséges tápfolyadékok elterjedése (Ham’s F12). Sejtvonalak ellenőrzése: Kariogram Kromoszóma szám Mikoplazma Spontán kromoszóma aberráció gyakoriság Történeti kontroll
 jó növekedési képesség, rövid generációs idő, kariotípus stabilitás, állandó krom. szám. Kromoszómák alaki változatossága és stabilitása. kínai hörcsög ovárium fibroblaszt sejt (CHO, Puck1957) Tápfolyadékok, tenyésztési körülmények. Mesterséges tápfolyadékok elterjedése (Ham’s F12). Sejtvonalak ellenőrzése: Kariogram. Kromoszóma szám. Mikoplazma. Spontán kromoszóma aberráció gyakoriság. Történeti kontroll.")
28
Vizsgálati anyag előkészítése:
kémiai összetétel, szennyeződés, stabilitás oldószer kiválasztása hígítási sor készítése stabilitás vizsgálat archiválás Elő-kísérlet: citotoxicitási vizsgálat MTT-assay (mitokondriális szukcinát-dehidrogenáz enzim) mitotikus index meghatározása MI: A metafázisban lévő sejtek és a sejtpopuláció összes sejtjének aránya. A sejtpopuláció proliferációjának a mértékét jellemzi. Koncentráció Legkevesebb három elemezhető koncentráció. Citotoxikus anyag esetén: va. az a koncentrációja amely a festék-redukciót 50%-kal csökkenti. Nem citotoxikus anyag esetén: va. 5 mg/ml v. 0,01M
 mitotikus index meghatározása. MI: A metafázisban lévő sejtek és a sejtpopuláció összes sejtjének aránya. A sejtpopuláció proliferációjának a mértékét jellemzi. Koncentráció. Legkevesebb három elemezhető koncentráció. Citotoxikus anyag esetén: va. az a koncentrációja amely a festék-redukciót 50%-kal csökkenti. Nem citotoxikus anyag esetén: va. 5 mg/ml v. 0,01M.")
29
Metabolikus aktiválás:
Direkt-és indirekt-mutagén anyagok Metabolikus aktiválás: S9 alkalmazása rágcsálók enziminducerrel (Arochlor 1254, v. fenobarbiturát és β-naftoflavon) kezelt májából előállított poszmitokondriális frakció (S9) kofaktorokkal kiegészítve (ADPH, glukóz-6-foszfát). S9 végkoncentráció 5-10% közötti. citokróm P450 enzim aktiválása. Indirekt mutagének kimutatására alkalmas
 kezelt májából előállított poszmitokondriális frakció (S9) kofaktorokkal kiegészítve. (ADPH, glukóz-6-foszfát). S9 végkoncentráció 5-10% közötti. citokróm P450 enzim aktiválása. Indirekt mutagének kimutatására alkalmas.")
30
Kontrollok: Kezelési idők:
Metabolikus aktiválás nélkül: MMS, EMS, Mitomycin-C Metabolikus aktiválással: CP, Benz(a)pirén Oldószer: tápfolyadék, DMSO Archiválás Kezelési idők: 4 óra +S9 4 óra -S9 ÉRTÉKELÉS pozitív Negatív ismétlés 24 óra -S9 48 óra -S9
pirén. Oldószer: tápfolyadék, DMSO. Archiválás. Kezelési idők: 4 óra +S9. 4 óra -S9 ÉRTÉKELÉS. pozitív Negatív. ismétlés 24 óra -S9. 48 óra -S9.")
31
Célja a metafázisos kromoszómák összegyűjtése
Kolhicinezés: Célja a metafázisos kromoszómák összegyűjtése Őszi kikerics (Colchicum autumnale) hagymájából készült kivonat. Köszvény gyógyítására alkalmazták. A kolhicin sejtosztódás gátló, a mitózis metafázisában. Normál esetben a sejtek 2-5%-a osztódik, néhány órás kolhicinezés összegyűjti a sejteket a metafázisban, akár százalékuk is blokkolt metafázisban található. A kolhicin a magorsó mikrotubulusaihoz kötődik.
 hagymájából készült kivonat. Köszvény gyógyítására alkalmazták. A kolhicin sejtosztódás gátló, a mitózis metafázisában. Normál esetben a sejtek 2-5%-a osztódik, néhány órás kolhicinezés összegyűjti a sejteket a metafázisban, akár százalékuk is blokkolt metafázisban található. A kolhicin a magorsó mikrotubulusaihoz kötődik.")
32
Kromoszóma preparátum
Fénymikroszkópos értékelés kromatid típusú aberrációk: kromoszóma típusú aberrációk: deléció deléció exchange exchange

33
CHO sejt, kezeletlen kontroll. Felvétel: OKBI. MSBO. 2010.

34
CHO sejt, Deléció. Felvétel OKBI. MSBO. 2000.

35
CHO sejt. Deléció. Felvétel: OKBI. MSBO 2000.

36
Humán limfocita. Kezeletlen kontroll. Felvétel: OKBI. MSBO. 2000.

37
A környezeti arzén-expozíció egészségkárosító hatásai
Arzénmérgezés” Teratogenitás; neurotoxikus hatás Citotoxikus hatás (immuntoxicitás?) Hormon diszruptor hatás Glukokortikoid receptor gátlás Ösztrogén receptor gátlás Daganatkeltő hatás Géntoxicitás (oxidatív stressz) Repair gátlás (ligáz gátlás) Epigenetikus hatás (metiláció) Növekvő dózis
 Hormon diszruptor hatás. Glukokortikoid receptor gátlás. Ösztrogén receptor gátlás. Daganatkeltő hatás. Géntoxicitás (oxidatív stressz) Repair gátlás (ligáz gátlás) Epigenetikus hatás (metiláció) Növekvő dózis.")
38
Kromoszóma aberráció vizsgálat in vitro emlős sejteken
OECD TG 473 Kromoszóma aberráció Normál metafázis Fragmentált kromoszómák Endomitózis CHO-K1 (ATCC-CCL-61; Chinese hamster) Kínai hörcsög ovárium fibroblaszt Felvételek készültek: OKBI 2011 Molekuláris és Sejtbiológiai Osztály
 Kínai hörcsög ovárium fibroblaszt. Felvételek készültek: OKBI Molekuláris és Sejtbiológiai Osztály.")
39
B.19. IN VITRO EMLŐSSEJT TESTVÉR – KROMATID KICSERÉLŐDÉS
(SISTER CHROMATID EXCHANGE , SCE) VIZSGÁLAT OECD TG 479 A testi sejtek kromoszómáiban a testvér-kromatidák közötti kicserélődés, reciprok DNS csere. Láthatóvá tehető, ha két sejtcikluson át bróm-dezoxi-uridinnel (timidin-analóg) jelöljük. Hoechst festés, majd UV kezelés, klasszikus Giemsa festés. Az SCE nem kromoszómaaberráció (Ctr. SCE 5). Az SCE már olyan kis vegyi mutagén-karcinogén expozíció kiváltja, amelyik még nem okoz krom. aberrációt. Alkalmazása primer prevencióban.
 VIZSGÁLAT. OECD TG 479. A testi sejtek kromoszómáiban a testvér-kromatidák közötti kicserélődés, reciprok DNS csere. Láthatóvá tehető, ha két sejtcikluson át bróm-dezoxi-uridinnel (timidin-analóg) jelöljük. Hoechst festés, majd UV kezelés, klasszikus Giemsa festés. Az SCE nem kromoszómaaberráció (Ctr. SCE 5). Az SCE már olyan kis vegyi mutagén-karcinogén expozíció kiváltja, amelyik még. nem okoz krom. aberrációt. Alkalmazása primer prevencióban.")
40
Az SCE indukcióban a repair folyamatok részvétele valószínű.
A DNS repair deficiens congenitális defektusok esetében fokozott SCE indukáltság figyelhető meg az egészséges kontrollokhoz képest. pl.: Bloom szindróma Fanconi-anémia Werner sindróma xeroderma pigmentosum ataxia telangiectasia Dow kór

41
Ataxia telangiectasia (ATM)
Rendkívül sokféle, sok szervet érintő tünet együttes sugár érzékenység és tumorok kialakulása

42
Bloom szindróma Kromoszóma instabilitás jellemzi
Örökletes betegség, családi halmozódás 15q26.1, a BLM gén mutációja. Fokozott sister chromatid exchange (SCE) és spontán kromoszóma törékenység. DNS repair enzimek károsodása Jelentős mutagén túlérzékenység. a magasabb daganatképződési kockázat normál sejt Bloom szindromás beteg sejt
 és spontán kromoszóma törékenység. DNS repair enzimek károsodása. Jelentős mutagén túlérzékenység. a magasabb daganatképződési kockázat. normál sejt. Bloom szindromás beteg sejt.")
43
In vitro mikronukleusz teszt
OECD TG 487 Kromoszóma aberráció kimutatására alkalmas Kb 80% egyezés a kromoszóma aberráció teszttel, olcsóbb, gyorsabb Mikronukleusz: a sejtmagnál kisebb méretű, membránhatárolt DNS darabok, amelyek a citoplazmában jelennek meg a sejtosztódás zavara esetén mikronukleusz apoptózis

44
patkány primer májsejt
A nemhez kötött recesszív mutagenitási vizsgálat (Müller-5 teszt) elvi vázlata OECD TG B. 18. Nem-tervezett DNS-szintézis (UDS) in vitro vizsgálata emlős májsejteken OECD TG 482 patkány primer májsejt Jelölések: * = sárga testszínű kerek, piros szemű hím kék = kezeletlen X kromoszóma piros = kezelt X kromoszóma nyíl = Y kromoszóma Molinát és Vernolát gyomírtók repair-szintézist idukáló hatása primer patkánymájsejtben Egészségtudomány: 36, (1992) Kinoxalin származékok vizsgálata Népegészségügy 90 (1) (2012) A vizsgálatokat az OKBI molekuláris és Sejtbiológiai Osztályán végezték.
 elvi vázlata. OECD TG. B. 18. Nem-tervezett DNS-szintézis (UDS) in vitro vizsgálata emlős májsejteken OECD TG 482. patkány primer májsejt. Jelölések: * = sárga testszínű kerek, piros szemű hím. kék = kezeletlen X kromoszóma. piros = kezelt X kromoszóma. nyíl = Y kromoszóma. Molinát és Vernolát gyomírtók repair-szintézist. idukáló hatása primer patkánymájsejtben. Egészségtudomány: 36, (1992) Kinoxalin származékok vizsgálata. Népegészségügy 90 (1) (2012) A vizsgálatokat az OKBI molekuláris és Sejtbiológiai Osztályán végezték.")
45
Köszönöm a figyelmet 2012 ELTE 11.27.

Hasonló előadás

 DNS-ből,>")


 daganatkeltő.>")








 Bihari Péter.>")



 kialakulása Genetikai, Sejt- és Immunbiológiai Intézet Falus András.>")

