Előadást letölteni
Az előadás letöltése folymat van. Kérjük, várjon

1
A működés szabályozása
A tápcsatorna működésének szabályozási mechanizmusai KIR reflexes szabályozás az enterális idegrendszer – a motoros és szekréciós funkció szabályozása a gasztrointesztinális jelzőmolekulák A szövetek tápanyagellátásának hormonális szabályozása a hasnyálmirigy belső szekréciós funkciója az inzulin a glukagon és a szomatosztatin a tápanyag-raktározás és mobilizálás szabályozása A táplálékfelvétel és a test energiaraktárainak szabályozása

2
A tápcsatorna működésének szabályozási mechanizmusai
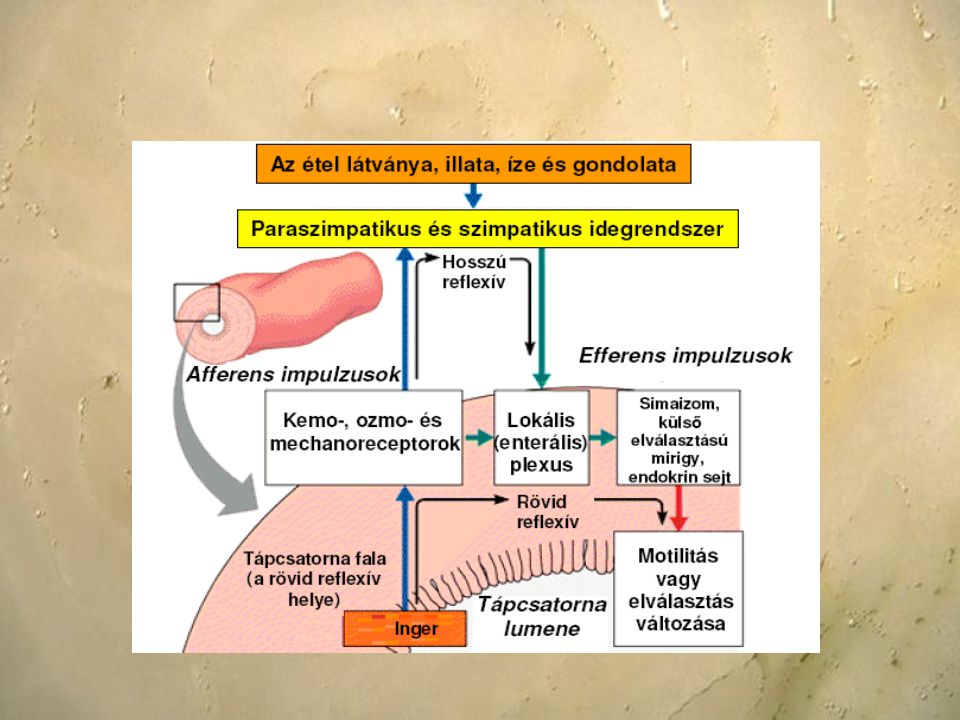
Az optimális működéshez a táplálékfelvételt követő motoros, szekréciós és felszívó folyamatokat össze kell hangolni, valamint az elfogyasztott táplálék mennyiségéhez és minőségéhez kell igazítani. A gasztrointesztinális rendszer működésének szabályozását hormonális és idegi mechanizmusok biztosítják: központi idegrendszeri enterális idegrendszer gasztrointesztinális hormonok, parakrin szekrétumok A szabályozó mechanizmusok a tápcsatorna adott szakaszai szerint változnak. A rágást, nyelést, nyálszekréciót és székletürítést a központi idegrendszer szabályozza, míg a gyomornedv elválasztását, a gyomor mozgását, a hasnyálmirigynedv elválasztását, az epe ürítését, a vékony- és vastagbél mozgását és a felszívódást az enterális idegrendszer szabályozza, amit a paraszimpatikus beidegzés nagyobb mértékben,a szimpatikus beidegzés kisebb mértékben módosít.

3
Központi idegrendszeri reflexes szabályozás
Egyes mirigyek elválasztását, továbbá a nyelőcső és a gyomor motoros működését a központi idegrendszer közvetlenül irányíthatja Pavlov: feltételes (kondícionált) reflex kialakítása – a nyálelválasztást vizsgálta
 reflex kialakítása – a nyálelválasztást vizsgálta.")
4
Pavlov kísérletei Pavlov az emésztés élettanát kutatta, munkásságáért Nobel díját is kapott. E kutatások részeként sajátos módszert dolgozott ki a kutyák evés közbeni nyálelválasztásának mérésére. A kutyát kantárokkal egy állványhoz kötötték, hogy ne tudjon mozogni, meghatározott mennyiségű ételt adtak neki, a képződött nyálat pedig, a pofájára varrt sipolyon keresztül elvezették. Gondosan ellenőrzött szituációt alakított ki, amelyben röviddel azelőtt, mielőtt a kutya ételt kapott volna, egy csengő szólalt meg, a csengő hangját és az ételt többször párosítva a csengő hangja önmagában is nyáladzást váltott ki. Pavlov az ételt feltétlen ingernek nevezte, a nyálelválasztást pedig feltétlen válasznak, minthogy ez a reakció minden körülmények között megvalósul.

5
Pavlov kísérletei Az új inger (a csengőhang) a feltételes inger nevet kapta, mivel ez korábban semleges inger volt, és nem váltott ki nyálelválasztási reakciót. A csengőhangra adott válasznak Pavlov a feltételes válasz nevet adta. A lejátszódó események sorrendje így ábrázolható: Pavlov szerint a feltételes inger és a feltétlen inger között képződött kapcsolat annak a következménye, hogy a csengőszó elhangzása után röviddel kapott étel megerősítette a nyálelválasztási reakciót. Ha az étel rendszeresen elmarad, a feltételes reakció is kezd elmaradozni, illetve kioltódik. Pavlov kimutatta, hogy az a kutya, amelyik megtanult csengő szóra nyáladzani, más - hasonló - ingerekre, így például berregésre, sípszóra is nyálelválasztással válaszol.
 a feltételes inger nevet kapta, mivel ez korábban semleges inger volt, és nem váltott ki nyálelválasztási reakciót. A csengőhangra adott válasznak Pavlov a feltételes válasz nevet adta. A lejátszódó események sorrendje így ábrázolható: Pavlov szerint a feltételes inger és a feltétlen inger között képződött kapcsolat annak a következménye, hogy a csengőszó elhangzása után röviddel kapott étel megerősítette a nyálelválasztási reakciót. Ha az étel rendszeresen elmarad, a feltételes reakció is kezd elmaradozni, illetve kioltódik. Pavlov kimutatta, hogy az a kutya, amelyik megtanult csengő szóra nyáladzani, más - hasonló - ingerekre, így például berregésre, sípszóra is nyálelválasztással válaszol.")
6
Központi idegrendszeri reflexes szabályozás
A táplálékfelvétel alatt, továbbá a táplálékfelvételt követően az emésztőcsatorna különböző részein (szájüreg, gyomor, vékonybél) elhelyezkedő receptorok ingerületbe kerülnek és a központi idegrendszerben (KIR) átkapcsolódva (elsősorban nyúltvelőben) kiváltják a mirigyek szekretomotoros idegeinek ingerületét. >>> a KIR ebben az esetben közvetlenül hat a mirigyek acinussejtjeire. A látószerv és a hallószerv ingerülete is szekréciót okoz.
 elhelyezkedő receptorok ingerületbe kerülnek és a központi idegrendszerben (KIR) átkapcsolódva (elsősorban nyúltvelőben) kiváltják a mirigyek szekretomotoros idegeinek ingerületét. >>> a KIR ebben az esetben közvetlenül hat a mirigyek acinussejtjeire. A látószerv és a hallószerv ingerülete is szekréciót okoz.")
7
Az enterális idegrendszer
A vegetatív idegrendszer harmadik része a szimpatikus és paraszimpatikus mellett az enterális idegrendszer. A nyelőcső alsó kétharmadától egészen a rectum végéig húzódik, magában foglalja a hasnyálmirigy idegelemeit is. Két egymástól elkülönült, de egymással szoros kapcsolatban álló idegfonatból áll, ezek a hosszanti és körkörös simaizom-rétegek közt elhelyezkedő plexux myentericus Auerbachi és a nyálkahártya alatt húzódó plexux submucosus Meissneri. Az enterális idegrendszert felépítő neuronok - a központi idegrendszerhez hasonlóan - reflexíveket alkotnak. Az érző (szenzoros) neuronok végződései részben mechanoreceptorok, amelyek a gyomor- bélrendszer falának feszülését érzékelik, részben pedig a gyomor- béltartalom összetételét detektáló kemoreceptorok.
 neuronok végződései részben mechanoreceptorok, amelyek a gyomor- bélrendszer falának feszülését érzékelik, részben pedig a gyomor- béltartalom összetételét detektáló kemoreceptorok.")
9
Az enterális idegrendszer
A szenzoros neuronok transzmittere mai ismereteink szerint a P-anyag (substance-P, SP). A szenzoros és effektorneuronok között a kapcsolatot igen sok interneuron biztosítja, amelyek között egyaránt vannak serkentők és gátlók. A serkentő interneuronok kolinergek, a gátló interneuronok többféle transzmiterrel működnek, ezek közül különösen fontosak az opoid peptidek (endorphinok, encephalinok). Az effektorneuronok simaizmokat, mirigyeket, endokrin, és parakrin sejteket valamint ereket idegeznek be. A submucosus plexus effektorneuronjai - többek között - kolinerg úton irányítják az emésztőmirigyek enzimszekrécióját, a myentericus effektorok pedig szerotoninerg transzmisszióval a motilitást. Az enterális idegrendszer helyi reflexeit a vegetatív idegrendszer másik két ága, a szimpatikus és a paraszimpatikus modulálja.
. A szenzoros és effektorneuronok között a kapcsolatot igen sok interneuron biztosítja, amelyek között egyaránt vannak serkentők és gátlók. A serkentő interneuronok kolinergek, a gátló interneuronok többféle transzmiterrel működnek, ezek közül különösen fontosak az opoid peptidek (endorphinok, encephalinok). Az effektorneuronok simaizmokat, mirigyeket, endokrin, és parakrin sejteket valamint ereket idegeznek be. A submucosus plexus effektorneuronjai - többek között - kolinerg úton irányítják az emésztőmirigyek enzimszekrécióját, a myentericus effektorok pedig szerotoninerg transzmisszióval a motilitást. Az enterális idegrendszer helyi reflexeit a vegetatív idegrendszer másik két ága, a szimpatikus és a paraszimpatikus modulálja.")
10
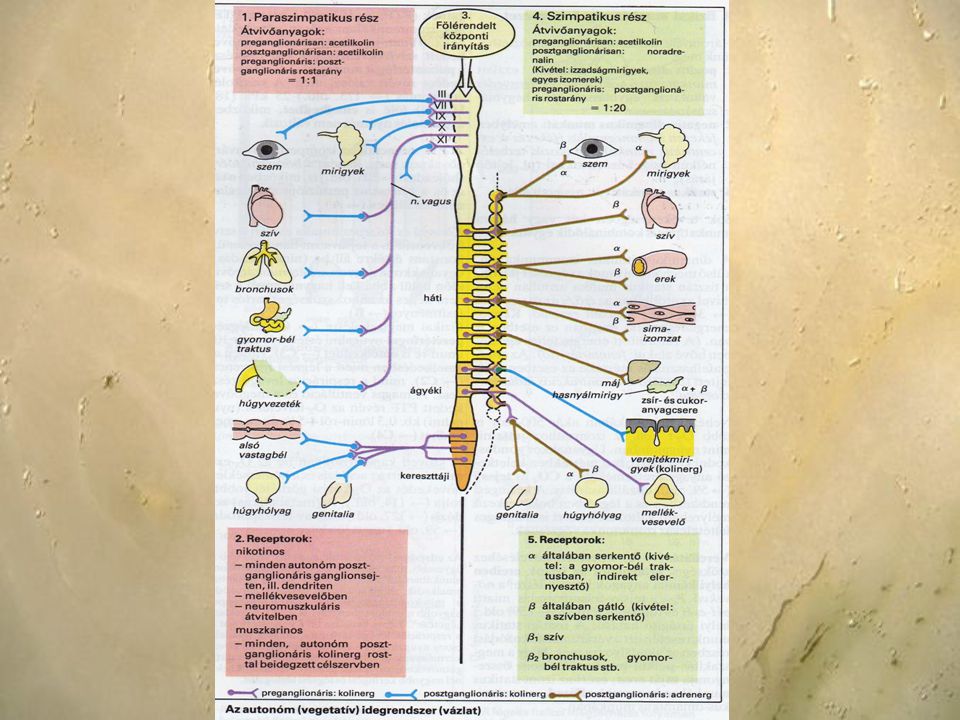
A vegetatív idegrendszer két - egymástól anatómiailag és funkcionálisan elkülönülő részből áll. A két rendszer közös vonása, hogy az efferens szár két neuronos: a központi idegrendszerben kezdődő preganglionáris, és valamelyik vegetatív ganglionban elhelyezkedő posztganglionáris neuronból áll. A ganglionokban az ingerület áttevődés folyamata is azonos: a neurotranszmitter az acetilkolin. Különbözik viszont a preganglionáris sejt és a ganglion elhelyezkedése. A szimpatikus idegrendszer preganglionáris rostjai a gerincvelő thoracolumális szakaszából, a szürkeállomány mediolaterális részéről indulnak ki. A ganglionok paravertebrálisan (dúclánc) vagy prevertebrálisan (ggl. stellatum, ggl.coeliaca, ggl. mesenterica sup. et inf.) helyezkednek el. A preganglionáris rostok tehát viszonylag rövidek, és a posztganglionáris rostok hosszabbak. A neurotranszmitter a ganglionokban az acetilkolin, a posztganglionáris rostok túlnyomó részében (90%) pedig a noradrenalin.
 vagy prevertebrálisan (ggl. stellatum, ggl.coeliaca, ggl. mesenterica sup. et inf.) helyezkednek el. A preganglionáris rostok tehát viszonylag rövidek, és a posztganglionáris rostok hosszabbak. A neurotranszmitter a ganglionokban az acetilkolin, a posztganglionáris rostok túlnyomó részében (90%) pedig a noradrenalin.")
11
A paraszimpatikus idegrendszer preganglionáris rostjai craniosacralis kiindulásúak. A craniális részt az agyidegek (III, VII, LX, X) vegetatív magvai képezik, ezek közül legfontosabb a vagus, ami a mellkasi és a felső hasi szervek paraszimpatikus beidegzését adja. A gerincvelő sacralis szegmentumából induló rostok csak a has alsó harmadában elhelyezkedő szervek vegetatív beidegzésében játszanak szerepet. A ganglionok a szervek falában foglalnak helyet, tehát a preganglionáris rostok hosszúak, míg a posztganglionárisak igen rövidek. Mind a pre- mind a posztganglionáris rost neurotranszmittere az acetilkolin.
 vegetatív magvai képezik, ezek közül legfontosabb a vagus, ami a mellkasi és a felső hasi szervek paraszimpatikus beidegzését adja. A gerincvelő sacralis szegmentumából induló rostok csak a has alsó harmadában elhelyezkedő szervek vegetatív beidegzésében játszanak szerepet. A ganglionok a szervek falában foglalnak helyet, tehát a preganglionáris rostok hosszúak, míg a posztganglionárisak igen rövidek. Mind a pre- mind a posztganglionáris rost neurotranszmittere az acetilkolin..")
12
Az emésztőrendszer károsodásai
Lonovics János id. Dubecz Sándor Erdős László Juhász Ferenc Misz Irén Írisz

13
Az emésztőrendszer károsodásai
Lonovics János id. Dubecz Sándor Erdős László Juhász Ferenc Misz Irén Írisz

16
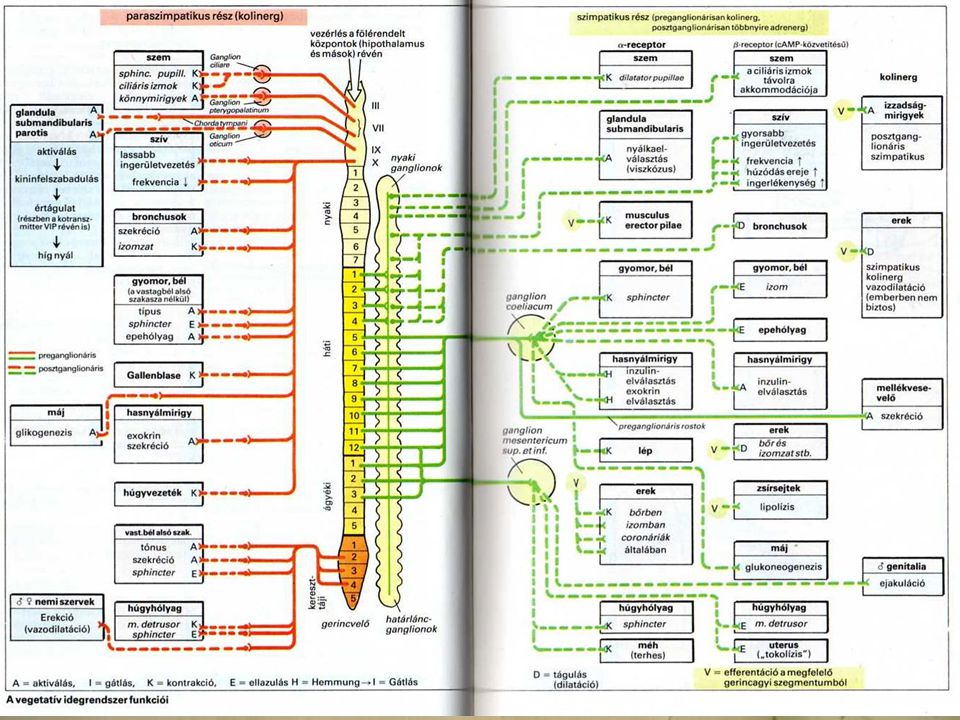
Paraszimpatikus posztganglionáris hatások
simaizom: gastrointestinális rendszer kontrakció mirigyek: nyálmirigyek szekréció gyomor fedősejtek HCl szekréció gyomor fősejtek pepszinogén szekréció pancreas acinussejtek enzim szekréció Szimpatikus hatások és receptorok Szerv - beidegzett struktúra Receptor Hatás Emésztőrendszer mirigyek simaizom sphincterek b2 a1 szekréció csökken motilitás csökken kontrakció - zárás Májsejtek glikogenolízis Zsírsejtek b3 lipolízis

17
A vagovagalis reflex Az emésztőcsatorna működését szabályozó feltétlen reflexek afferens neuronjainak nagy része az agyidegeken keresztül éri el a nyúltvelőt. Legfontosabb. N. vagus (X. agyideg) A nyelőcsőből, a gyomorból és a vékonybélből származó információt szállítja. (a szenzoros receptorok mechano, ozmo- és kemoreceptorok). Az efferens rostok szintén a n. vagusban futnak. = vagovagalis reflex
 A nyelőcsőből, a gyomorból és a vékonybélből származó információt szállítja. (a szenzoros receptorok mechano, ozmo- és kemoreceptorok). Az efferens rostok szintén a n. vagusban futnak. = vagovagalis reflex.")
19
Gasztrointesztinális jelzőmolekulák
A táplálékfelvételt követő mechanikai vagy kémiai változás a tápcsatornának valamely szakaszában egy specifikusan differenciált sejtcsoportból hormonszekréciót vált ki, amely a a véráramba kerülve távoli sejtek működését szabályozza.

20
A tápcsatorna falában speciális hormonokat elválasztó sejtek is találhatók
• Az ilyen endokrin sejtek működését a béllumen felől érkező hatások, más hormonok és az enterális idegrendszer befolyásolja. • A hormonok parakrin módon, és a vérrel a célsejthez szállítódva befolyásolják a tápcsatorna szekretoros és motoros működését • A hormonok a tápcsatorna mozgását, a külső- és belső elválasztású mirigyek aktivitását és a mirigyszekrétumok ürülését befolyásolják • A gasztrointesztinális hormonok kivétel nélkül peptidek, szerkezetük alapján három csoportot különböztetünk meg: - gasztrin-család: gasztrin és kolecisztokinin (CCK) - szekretin-család: szekretin, glükagon, vazoaktív intesztinális peptid (VIP) - motilin-család: motilin
 - szekretin-család: szekretin, glükagon, vazoaktív intesztinális peptid (VIP) - motilin-család: motilin.")
21
A gasztrointesztinális rendszer endokrin működése
• A endokrin sejteket az általuk termelt hormonról nevezték el: G-sejt (gasztrin), CCK-sejt (CCK), S-sejt (szekretin), M-sejt (motilin), stb. • A hormonszekréciót a vegetatív idegi hatások mellett a lumenben található speciális anyagok is elindíthatják: - gasztrin: gyomorban fehérje-bomlástermékek, PSY hatás, gyomorfal feszülése - szekretin: duodenum nyálkahártyáját ér savi hatás - CCK: duodenumban lév zsírok és aminosavak
, CCK-sejt (CCK), S-sejt (szekretin), M-sejt (motilin), stb. • A hormonszekréciót a vegetatív idegi hatások mellett a lumenben található speciális anyagok is elindíthatják: - gasztrin: gyomorban fehérje-bomlástermékek, PSY hatás, gyomorfal feszülése. - szekretin: duodenum nyálkahártyáját ér savi hatás. - CCK: duodenumban lév zsírok és aminosavak.")
22
G-sejtek : gasztrin Parietalis sejt: HCl, intrinsic factor Trofikus hatás (gyomor, vékony- és vastagbél) Gyomor motilitása Exokrin hasnáylmirigy Inzulin Pajzsmirigy: Kalcitonin Bél motilitása gyomor antrumának és a duodenum nyálkahártyájában elszórtan - apicalis rész a lumen kémiai összetételét érzékeli: aminosavak, aminok Helicobacter pylori ammóniát termel - basalis rész: szekréció a kapilláris felé G-sejten 2 idegi végződés konvergál: (növelik a gasztrinleadást) bombezinerg neuron transzmittere: GRP (gastrin releasing peptid) (vagus postgang. neuronjaként felfogható, és ezen végződnek az enterális IR rostjai is) - sy postgang neuronok : NAβ2 –rec vér adrenalin szint emelkedése aktiválja stressz okozta hypergastrinaemiasósav túlprodukció
 bombezinerg neuron transzmittere: GRP (gastrin releasing peptid) (vagus postgang. neuronjaként felfogható, és ezen végződnek az enterális IR rostjai is) - sy postgang neuronok : NAβ2 –rec vér adrenalin szint emelkedése aktiválja stressz okozta hypergastrinaemiasósav túlprodukció.")
23
Gátló szabályozás: negatív visszacsatolás a gyomor lumene felől: gasztrin fedősejtek sósavat termelneklumenbe került H+-ok gátolják a gasztinszekréciót. (pH 2,5-3,0-nál) gyomornyh-ban levő D-sejtek: egy részük a G-sejt mellett szomatosztatint termelnek G-sejthez diffundálköt a G-sejthez gátolja a gasztrin szekréciót (parakrin) (D-sejt idegi szab. alatt is áll: psy postgang kolinerg aktivitás szomatosztatin szekréciója nő (atropin: szomatosztatinszekr-t gátol gasztrin szint nő) kolinerg ingerület : közvetlenül hatva a fedősejtensósav szekr. nő) gasztrin rövid és hosszútávú hatásai sósavszekréció biztosítása táplálékfelvételkor közvetett hatás: gyomor nyh-ban levő enterokromaffin sejtek gasztrinreceptorára hat hisztamin szabadul fel parakrin módon növeli a fedősejtek sósavszekrécióját közvetlen hatás (kisebb jelentőség) fedősejtek, enterokromaffin sejtek növekedési faktora A gyomornyálkahártya sejtjeinek proliferációja
 gyomornyh-ban levő D-sejtek: egy részük a G-sejt mellett szomatosztatint termelnek G-sejthez diffundálköt a G-sejthez gátolja a gasztrin szekréciót (parakrin) (D-sejt idegi szab. alatt is áll: psy postgang kolinerg aktivitás szomatosztatin szekréciója nő (atropin: szomatosztatinszekr-t gátol gasztrin szint nő) kolinerg ingerület : közvetlenül hatva a fedősejtensósav szekr. nő) gasztrin rövid és hosszútávú hatásai. sósavszekréció biztosítása táplálékfelvételkor. közvetett hatás: gyomor nyh-ban levő enterokromaffin sejtek gasztrinreceptorára hat hisztamin szabadul fel parakrin módon növeli a fedősejtek sósavszekrécióját. közvetlen hatás (kisebb jelentőség) fedősejtek, enterokromaffin sejtek növekedési faktora. A gyomornyálkahártya sejtjeinek proliferációja.")
24
CCK - epehólyag izomzat összehúzódása - Oddi-sphincter ellazulása - Hasnyálmirigy acinussejtek enzimszekréciójának fokozása - Hasnyálmirigy kivezető csősejt: elektrolitszekréció (szekretin hatását fokozza) - gyomorürülést gátolja - inzulinszekréciót fokozza - táplálékfelvételt gátolja (jóllakottság, vagus afferens) trofikus hatás a hasnyálmirigyben CCK-termelő sejtek: dudodenumban, jejunum felső szakaszán Legfontosabb szabályozója a lumen tartalmának összetétele: aminosavak (főleg fenilalanin, zsírsavak C>10) Vagovagalis választ is kivált tápcsatorna adott szakaszán megfelelő mennyiségű táplálék találkozzon a megfelelő mennyiségű emésztőnedvvel lebontási, felszívási viszonyok optimalizálása, közben a vér glükóz koncentrációja csak keveset ingadozzon pleiotróp hatás
 - gyomorürülést gátolja. - inzulinszekréciót fokozza. - táplálékfelvételt gátolja (jóllakottság, vagus afferens) trofikus hatás a hasnyálmirigyben. CCK-termelő sejtek: dudodenumban, jejunum felső szakaszán. Legfontosabb szabályozója a lumen tartalmának összetétele: aminosavak (főleg fenilalanin, zsírsavak C>10) Vagovagalis választ is kivált. tápcsatorna adott szakaszán megfelelő mennyiségű táplálék találkozzon a megfelelő mennyiségű emésztőnedvvel lebontási, felszívási viszonyok optimalizálása, közben a vér glükóz koncentrációja csak keveset ingadozzon pleiotróp hatás.")
25
Szekretin - Hasnyálmirigy ductus-sejtek: HCO3-szekréció - Epeutak sejtjei: HCO3--szekréció - Gyomor HCl-elválasztás gátlása - Gyomor pepszinszekréció - CCK-hatás potencírozása Gyomorürülés gátlása Ingere: a duodenumot érő savas vegyhatás Gátlása: savi vegyhatás megszűnése GIP – glükóz dependens inzulinotrop peptid Szekrécióját abélbekerülő szénhidrát is kiváltja inzulin-szekréció serkentése (fő hatás) Gátló hatás a gyomorban: motilitás, HCl-elválasztás
 Gátló hatás a gyomorban: motilitás, HCl-elválasztás.")
26
Motilin Perisztaltikus aktivitás serkentése: Az étkezések közötti motilitás feltételezett regulátora. (Hormonjelöltek PPP (pancreas F sejt)gátolja az exocrin szekréciót, abszorpció. Ingere: peptidek, vagus GLP-1 Serkenti bélnedv szekrécióját, insulin szekrécióját Végig a GI traktusban. Inger: glc neurotensin (ileum) GI-motilitás gátlása ilealis véráramlás serkentése inger: zsírsavak guanylin (vékony és vastagbél, parakrin) cGMP intestinalis szekréció serkentése peptid YY (vékonybél) HCl szekréció gátlása, inger: zsír (villikinin)-nem izolált (vékonybél) bolyhok mozgása)
gátolja az exocrin szekréciót, abszorpció. Ingere: peptidek, vagus. GLP-1 Serkenti bélnedv szekrécióját, insulin szekrécióját Végig a GI traktusban. Inger: glc. neurotensin (ileum) GI-motilitás gátlása ilealis véráramlás serkentése inger: zsírsavak. guanylin (vékony és vastagbél, parakrin) cGMP intestinalis szekréció serkentése. peptid YY (vékonybél) HCl szekréció gátlása, inger: zsír. (villikinin)-nem izolált (vékonybél) bolyhok mozgása)")
27
Parakrin anyagok Szomatosztatin D-sejtek termelik, olyan sejtek közelében helyzezkednek el, melyek működését gátolják - gátolja a G-sejt gasztrinszekrécióját és a parietális sejt HCl szekrécióját ill. a gyomor mozgásait - végig a belekben: gátolja : szekretin, GIP, VIP szekréciót, pankreas szekréciót, epeutak kontrakcióját, belek szekrécióját és motilitását, abszorpciót hisztamin: H2 receptoron hat, szekretoros hatás Neurocrinok - GRP: G-sejt gasztrinszekréció - P-anyag: motilitás serkentése - enkephalinópiátreceptorVIP neuronok gátlása tónusos simaizom kontrakció motilitás gátlása - VIP: exocrin szekréció fokozása (erős inger nyál, pancreas, epe, bél), HCl szekréció gátlása, vasodilatatio, izomrelaxáció (sphincterek)
, HCl szekréció gátlása, vasodilatatio, izomrelaxáció (sphincterek)")
28
Összefoglalás A táplálék feldarabolása és nyállal történő keverése részben akaratlagos, részben reflexes folyamat. A nyálelválasztás idegi szabályozás alatt áll A paraszimpatikus idegrendszer ingerlése erőteljes nyálelválasztást okoz( híg és kevés szerves anyagot tartalmaz) A szimpatikus idegek ingerlése hatására a gl. submandibularisból kis mennyiségű, de szerves anyagokban gazdag nyál elválasztása indul meg A nyelés a tápláléknak a szájból a gyomorba történő juttatása. Kezdeti fázisa akaratlagos, ezt követően azonban reflexes szabályozás alatt áll. A faringeális fázisban, amikor a falat (bolus) a garatba jut, a gégefedő reflexes mechanizmus révén lezárja a tranchea bemenetét, s a légzés leáll.
 A szimpatikus idegek ingerlése hatására a gl. submandibularisból kis mennyiségű, de szerves anyagokban gazdag nyál elválasztása indul meg. A nyelés a tápláléknak a szájból a gyomorba történő juttatása. Kezdeti fázisa akaratlagos, ezt követően azonban reflexes szabályozás alatt áll. A faringeális fázisban, amikor a falat (bolus) a garatba jut, a gégefedő reflexes mechanizmus révén lezárja a tranchea bemenetét, s a légzés leáll.")
29
Összefoglalás

30
Összefoglalás

31
Összefoglalás A sósav szekrécióját PSY beidegzés, gasztrin és a hisztamin fokozza, míg a szekretin gátolja • A sósav szekréciója ízek és illatok (kefalikus fázis), a gyomorbennék (gasztrikus fázis) és a vékonybélbe került fehérjék (intesztinális fázis) is kiválthatják A hasnyálmirigynedv szekréciót a CCK, szekretin, VIP, PSY hatás fokozza • Kefalikus- (látás, szaglás, rágás), gasztrikus- (gyomrfal feszülése) és intesztinális (vékonybélbe került peptidek, aminosavak, zsírsavak) fázis Az epe termelését a szekretin, ürülését a CCK szabályozza, valamint mind a termelés, mind az ürítés fokozódik PSY hatásra.
, a gyomorbennék (gasztrikus fázis) és a vékonybélbe került fehérjék (intesztinális fázis) is kiválthatják. A hasnyálmirigynedv szekréciót a CCK, szekretin, VIP, PSY hatás fokozza. • Kefalikus- (látás, szaglás, rágás), gasztrikus- (gyomrfal feszülése) és intesztinális (vékonybélbe került peptidek, aminosavak, zsírsavak) fázis. Az epe termelését a szekretin, ürülését a CCK szabályozza, valamint mind a termelés, mind az ürítés fokozódik PSY hatásra.")
32
Összefoglalás

33
Összefoglalás

34
Összefoglalás A végbél nyílását egy belső simaizom és egy külső, akaratlagosan szabályozható harántcsíkolt izom szabályozza • Míg a rectum telődése a belső sphincter reflexes ellazulását okozza, a külső sphincter tónusa fokozódik. A székelési ingert a rectum falának feszülése váltja ki. • A székletürítés komplex folyamat, amit kortikális és szubkortikális struktúrák (hypothalamus, nyúltvelő), a gerincvelő, és az enterális idegrendszer összehangolt működése hoz létre.
, a gerincvelő, és az enterális. idegrendszer összehangolt működése hoz létre.")
35
Szénhidrát anyagcsere
A szövetek tápanyagellátásának hormonális szabályozása Szénhidrát anyagcsere Anabolizmus Katabolizmus Hormonok Inzulin↑ Glukagon↓ Inzulin↓ Glukagon↑ Forrás táplálkozás raktárak Folyamatok Glikogén szintézis Triglicerid szintézis Protein szintézis Glikogenolízis Lipolízis Proteolízis Ketogenezis Forrás:

36
A glükóz homeosztázis szabályozása
Hormonális hatások: gyorsan ható hormonok: inzulin, glukagon, adrenalin lassan ható hormonok: kortizol, növekedési hormon I Idegi hatások : paraszimpatikus hatás (Ach) szimpatikus hatás (noradrenalin) Szubsztrátok: glükóz, aminosavak, szabad zsírsav
 szimpatikus hatás (noradrenalin) Szubsztrátok: glükóz, aminosavak, szabad zsírsav.")
37
Szénhidrát anyagcsere
Forrás:

38
Lipid anyagcsere Forrás:

39
Fehérje anyagcsere Forrás:

40
Intermedier anyagcsere
Forrás:

41
Hasnyálmirigy Langerhans-szigetek

42
Inzulin Forrás:

43
Inzulin Az inzulin szekréció 98 %-a szabályozott, 2 %-a konstitutív
• Felezési idő: 5-8 perc (C-peptid felezési ideje: 30 min) • Lebomlás: – vesében és májban: specifikus proteáz és glutation-dependens transzhidrogenáz – IR-hoz kötődve (internalizáció) • Kis mennyiségben a vizelettel ürül • Kezelt diabetesben: specifikus antitestek Forrás:
 • Lebomlás: – vesében és májban: specifikus proteáz és glutation-dependens transzhidrogenáz. – IR-hoz kötődve (internalizáció) • Kis mennyiségben a vizelettel ürül. • Kezelt diabetesben: specifikus antitestek. Forrás:")
44
Forrás:http://phys. dote

45
Inzulin elválasztás Forrás:

46
Ligand kötődés → dimerizáció - Aktiválódnak a tirozin-kinázok
- Az aktivált receptor relé proteineket aktivál (inzulin Receptor Substrate IRS) - Egy tirozin-kináz dimer akár tíz intracelluláris proteint is aktiválhat Forrás:
 - Egy tirozin-kináz dimer akár tíz intracelluláris. proteint is aktiválhat. Forrás:")
47
Glükóz felvétel Glükóz transzporterek • GLUT-1 – nagy glükóz affinitás
– inzulin independens – a bazális glükóz transzportért felelős • GLUT-2 – kis glükóz affinitás – mindkét irányú glükóz transzportot lehetővé tesz • GLUT-3 • GLUT-4 – inzulin dependens – a fokozott metabolikus aktivitás facilitálja a membránba történő kihelyeződését • Zsírszövet – GLUT-1, GLUT-4 • Vázizom – GLUT-1, GLUT-4 (inzulin independens aktiváció !!!) • Máj – GLUT-2 • Agy – idegsejt: GLUT-1, GLUT-3, erek: GLUT-2 • Hasnyálmirigy β sejtek • Vese – GLUT-2 (SGLT1 és GLUT-1) • Bélhám Forrás:
 • Máj. – GLUT-2. • Agy. – idegsejt: GLUT-1, GLUT-3, erek: GLUT-2. • Hasnyálmirigy β sejtek. • Vese. – GLUT-2 (SGLT1 és GLUT-1) • Bélhám. Forrás:")
48
Forrás:http://phys. dote

49
Inzulinszerű aktivitással rendelkező anyagok Inzulin Proinzulin
Nem-szupprimálható inzulinszerű aktivitás Kis molekulatömegű frakció IGF-I (somatomedin C) IGF-II Somatomedin A Foetalis somatomedin Nagy molekulatömegü frakció (nem-szupprimálható inzulinszerű proteinek) Ha az inzulinszerű aktivitást a zsírszövet glükózfelvételével és gázcsere meghatározásával mérik, akkor a teljes aktivitás 7%-a gátolható inzulin ellenes antitesttel. A fennmaradó 93%-ot nem-szupprimálható inzulinszerű aktivitásnak nevezzük. Forrás:
 IGF-II. Somatomedin A. Foetalis somatomedin. Nagy molekulatömegü frakció. (nem-szupprimálható inzulinszerű proteinek) Ha az inzulinszerű aktivitást a zsírszövet glükózfelvételével és gázcsere. meghatározásával mérik, akkor a teljes aktivitás 7%-a gátolható inzulin. ellenes antitesttel. A fennmaradó 93%-ot nem-szupprimálható inzulinszerű. aktivitásnak nevezzük. Forrás:")
50
Glukagon • hasnyálmirigy α-sejtekben termelődik
• Egyláncú peptid, MW: 3,500; aa:29 – preprohormon MW: 18,000 – prohormon (glycentin) MW: 12,000 • Szignalizáció: cAMP (Gs) A glucagon metabolizmusa • Felezési idő: 5-10 perc • Lebomlás: – minden szövetben, de elsősorban a májban Forrás:
 MW: 12,000. • Szignalizáció: cAMP (Gs) A glucagon metabolizmusa. • Felezési idő: 5-10 perc. • Lebomlás: – minden szövetben, de elsősorban a májban. Forrás:")
51
A glukagon termelés szabályozása
Forrás:

52
Glukagon elválasztás Forrás:

53
Forrás:http://phys. dote

54
Az inzulin és glukagon antiparallel elválasztása
• Anabolikus és katabolikus fázisok között I/G ∼ 2 • Fizikai munka: inzulin ⇓ - Glukagon ⇑ - I/G ⇓ (0.05) • Vészreakció: inzulin ⇑ - Glukagon ⇑ • Éhezés: inzulin ⇓ - Glukagon ⇑ - I/G ⇓ (<0.5) • Táplálékfelvételt követően: – Vegyes táplálék :inzulin ⇑ - Glukagon ⇓ - I/G ⇑ (15-25) – Szénhidrát: inzulin ⇑ - Glukagon ⇓ - I/G ⇑ (25-30) – Protein: inzulin ⇑ - Glukagon ⇑ - I/G ⇑ (3-4) !!!!!! – Zsír: inzulin ⇑ - Glukagon ⇓ - I/G ⇑ Forrás:
 • Vészreakció: inzulin ⇑ - Glukagon ⇑ • Éhezés: inzulin ⇓ - Glukagon ⇑ - I/G ⇓ (<0.5) • Táplálékfelvételt követően: – Vegyes táplálék :inzulin ⇑ - Glukagon ⇓ - I/G ⇑ (15-25) – Szénhidrát: inzulin ⇑ - Glukagon ⇓ - I/G ⇑ (25-30) – Protein: inzulin ⇑ - Glukagon ⇑ - I/G ⇑ (3-4) !!!!!! – Zsír: inzulin ⇑ - Glukagon ⇓ - I/G ⇑ Forrás:")
55
Somatostatin • Két forma: 28 and 14 aa; hasonló hatások
• D sejtek termelik • Felezési idő: 2 perc • Gátolja a – GI traktus minden funkcióját – A glucagon és inzulin elválasztás Forrás:

56
A somatostatin elválasztás
Forrás:

57
A szövetek glükóz felvétele nem elegendő a funkció fenntartásához.
Hypoglycaemia Vércukor szint < 3.0mM A szövetek glükóz felvétele nem elegendő a funkció fenntartásához. A központi idegrendszer nagyon érzékeny: látás, beszéd, járás zavarok aggresszivitás kóma, halál Stressz válasz (adrenalin felszabadulás): sápadtság verejtékezés Forrás:
: sápadtság. verejtékezés. Forrás:")
58
Csökkent glükóz tolerancia • A DM osztályozása – IDDM – NIDDM
– egyéb formák • Gesztációs diabetes • Indukált DM – metaszteroid • génmutáció Forrás:

59
A diabetes mellitus rövid távú szövődményei
Forrás:

60
Zsír mobilizáció és szintézis
A zsírsavak igen fontos energia források, trigliceridek (neutrális zsírok) formájában a zsírszövetekben raktározódnak. További fontos szerepük van a foszfolipidek felépítésében is. A trigliceridek mobilizációja lipáz enzimek segítségével történik. A zsírsavak oxidációjuk előtt acil-CoA-vá aktiválódnak, oxidációjuk a mitokondrium mátrixában történik, ahová az acil-karnitin transzferáz szállítja az aktivált zsírsavakat. Oxidációjuk két szénatomonként β oxidációval történik: acetil-CoA, FADH2 és NADH keletkezik. A zsírsavak szintézise teljesen eltérő módon történik A szintézist a citoszólban egy ACP körül szerveződött szintáz komplex végzi malonil-CoA prekurzorból NADPH oxidoredukciós partnerrel. A neutrális zsírok és foszfolipidek közös intermedierje a foszfatidsav. A foszfolipidek diészter kötéseinek kialakulásakor citidin nukleotidok vesznek részt az aktivált intermedier kialakításában. A koleszterin szintézis acetil-CoA-ból több lépésben mevalonsav és aktivált izoprén egység intermediereken keresztül történik. A koleszterin fontos összetevője a membránoknak és a szérum lipoprotein komplexeknek. A koleszterin szint szabályozása több szinten történik.
 formájában a zsírszövetekben raktározódnak. További fontos szerepük van a foszfolipidek felépítésében is. A trigliceridek mobilizációja lipáz enzimek segítségével történik. A zsírsavak oxidációjuk előtt acil-CoA-vá aktiválódnak, oxidációjuk a mitokondrium mátrixában történik, ahová az acil-karnitin transzferáz szállítja az aktivált zsírsavakat. Oxidációjuk két szénatomonként β oxidációval történik: acetil-CoA, FADH2 és NADH keletkezik. A zsírsavak szintézise teljesen eltérő módon történik A szintézist a citoszólban egy ACP körül szerveződött szintáz komplex végzi malonil-CoA prekurzorból NADPH oxidoredukciós partnerrel. A neutrális zsírok és foszfolipidek közös intermedierje a foszfatidsav. A foszfolipidek diészter kötéseinek kialakulásakor citidin nukleotidok vesznek részt az aktivált intermedier kialakításában. A koleszterin szintézis acetil-CoA-ból több lépésben mevalonsav és aktivált izoprén egység intermediereken keresztül történik. A koleszterin fontos összetevője a membránoknak és a szérum lipoprotein komplexeknek. A koleszterin szint szabályozása több szinten történik.")
61
A koleszterin szint szabályozása
A koleszterin szintézis út elején a mevalonsav szintézise hormonális szabályozás alatt áll, ezen kívül az intracelluláris koleszterin feedback inhibitora az enzimnek. Magas intracelluláris koleszterin koncentráció gátolja az LDL receptorok szintézisét ezáltal az LDL mennyiség a véráramban koncentrálódik. Örökletes hiperkoleszterinémia esetén az LDL receptorok hibája miatt nincs megfelelő LDL felvétel így az extracelluláris koleszterin nem jut be a sejtekbe hanem a véráramban felhalmozódik fel.

62
A vegetatív funkciók magasabb szintű szabályozása
A vegetatív funkciók felsőbb szintű szabályozása az agytörzsben és a hypothalamusban történik. Az agytörzsben találhatók az alapvető, vitális központok, amelyek nélkül az élet fenntartása elképzelhetetlen. Itt történik a vérnyomás, a vérkeringés és a légzés szabályozása, valamint itt található számos vegetatív ill. védő reflex - köhögés, tüsszentés, csuklás, nyelés, hányás - központja is.

63
A hypothalamus leginkább olyan vegetatív funkciók irányításáért felelős, amelyeknek magatartási összetevőjük is van. Ezek a homeostasist, az önfenntartást, és a fajfenntartást szolgálják. A vegetatív funkciók szervezésével ezen a szinten kapcsolódik össze az endokrin rendszer, a belső elválasztású mirigyek nagy részének irányítása is. A hypothalamusba befutó kémiai és idegi ingerek alapján itt integrálódik számos homeosztatikus funkció (táplálékfelvétel, vízforgalom, anyagcsere, hőszabályozás), a szervezet növekedése és a szaporodással kapcsolatos funkciók, és itt szerveződik a nagy megterhelések elhárítását szolgáló vészreakció is. A hypothalamusból kifutó parancsok részben idegi részben hormonális természetűek.
, a szervezet növekedése és a szaporodással kapcsolatos funkciók, és itt szerveződik a nagy megterhelések elhárítását szolgáló vészreakció is. A hypothalamusból kifutó parancsok részben idegi részben hormonális természetűek.")
64
A táplálékfelvétel és a test energiaraktárainak szabályozása
A napi táplálékfelvétel a raktárak feltöltöttségét biztosítja. A táplálék felvétele az energiaszükséglettől függő, szabályozott folyamat. A táplálékfelvételt szabályozó mechanizmusok elsősorban az összenergiabevitelt illesztik a szükséglethez, de a szervezet a felvett táplálék minőségét is szabályozza. A táplálék megszerzésére irányuló tevékenység a biológiai szükséglet által motivált magatartás, összetett végrehajtó funkciókkal. A táplálékfelvételt ill. a visszautasítást nem kizárólag a táplálékfelvétel szükségessége szabályozza, hanem sok magatartást befolyásoló tényező (napi ritmus, sötétésg/világosság, ált. aktivitási szint, stb.) pszichoszociális tényezők!
 pszichoszociális tényezők!")
65
A táplálékfelvétel központi szabályozása
A táplálékfelvétel a hipotalamikus és az agytörzsi magvak, valamint a limbikus rendszer ellenőrzése alatt áll. A szénhidrát, zsír és fehérje iránti vágy kialakításában és megszüntetésben specifikus pályák és transzmitterek szerepelnek. A táplálékfelvétel gátlásában szerotonerg és dopaminerg pályák szerepelnek. es évek kutatásai: hipotalamusz ventromediális része – „jóllakottság központ” hipotalamusz laterális része – „éhség központ” később kiderült, hogy az agytörzsi magvaknak is szerepe van, ill. más neuroncsoportok és pályák együttesen szabályozzák a táplálékfelvételt. Az agytörzsben (nucleus tractus solitarii) található kemoszenzitív receptorok fogadják a perifériáról jövő és a vér kémiai összetételét jelző információkat. A felsőbb agyrészek közül a limbikus rendszer játszik kiemelt szerepet. Más felső agyrészeknek módosító szerepük van: ezen utóbbiakon keresztül a táplálék minsőgének jelzései, az illat, íz pozitív vagy negatív hatást gyakorolnak a hipotalamikus és agytörzsi központokra. Az integráció a hipotalamuszban történik.
 található kemoszenzitív receptorok fogadják a perifériáról jövő és a vér kémiai összetételét jelző információkat. A felsőbb agyrészek közül a limbikus rendszer játszik kiemelt szerepet. Más felső agyrészeknek módosító szerepük van: ezen utóbbiakon keresztül a táplálék minsőgének jelzései, az illat, íz pozitív vagy negatív hatást gyakorolnak a hipotalamikus és agytörzsi központokra. Az integráció a hipotalamuszban történik.")
66
A hypothalamus

67
Hypothalamus eredetű elhízás

68
2. Szénhidrát iránti vágy: hipotalamusz dorzomediális részén (nucleus pataventricularis) konvegálnak azok apályák, amelyek a szénhidrát iránti étvágy kiváltásában szerepelnek; egy-egy étkezés hatására többet fogyasztanak Noradrenalin (α2-receptoron) GABA Y-neuropeptid (NPY)(nuclus arcuatus sejtjeiben termelődik): a NPY-neuronokon helyezkednek el a zsírraktárak állapotát jelző leptin receptorai. A zsírraktárak csökkenésekor a hypothalamushoz kevesebb leptinmolekula kerül, ezért fokozódik az NPYsejtek aktivitása. >>> megnövekdett táplálékfelvétel, éhezésre jellemző reakciók A szénhidrát iránti vágy reggel a legnagyobb. A rendszer hormonálisan is szabályozott: a mellékvesekéreg glukokortikoid hormonjai fokozzák a neurotranszmitterek szintézisét, valamint a receptorok számát.
 GABA. Y-neuropeptid (NPY)(nuclus arcuatus sejtjeiben termelődik): a NPY-neuronokon helyezkednek el a zsírraktárak állapotát jelző leptin receptorai. A zsírraktárak csökkenésekor a hypothalamushoz kevesebb leptinmolekula kerül, ezért fokozódik az NPYsejtek aktivitása. >>> megnövekdett táplálékfelvétel, éhezésre jellemző reakciók. A szénhidrát iránti vágy reggel a legnagyobb. A rendszer hormonálisan is szabályozott: a mellékvesekéreg glukokortikoid hormonjai fokozzák a neurotranszmitterek szintézisét, valamint a receptorok számát.")
69
2. Zsír iránti étvágy: a hipotalamusz mediális magcsoportjait elérő másik pályarendszer.
Galanin (állatban biztosan, emberben valószínű): hatását az ösztrogén is befolyásolja opioid pepdtidek: a fehérje iránti vágyat is fokozzák. Ez a rendszer a késői órákban aktiválódik. 3. A táplálékfelvétel gátlása szerotonerg pályák (a mediális hipotalamusz magjaink konvergálnak) gátolják a szénhidrát iránti étvágyat és valószínűleg ezek játszanak szerepet a szénhidrát iránti vágy nappali fokozatos csökkenésében - Dopaminerg pályák a zsírfogyasztást módosítják
: hatását az ösztrogén is befolyásolja. opioid pepdtidek: a fehérje iránti vágyat is fokozzák. Ez a rendszer a késői órákban aktiválódik. 3. A táplálékfelvétel gátlása. szerotonerg pályák (a mediális hipotalamusz magjaink konvergálnak) gátolják a szénhidrát iránti étvágyat és valószínűleg ezek játszanak szerepet. a szénhidrát iránti vágy nappali fokozatos csökkenésében. - Dopaminerg pályák a zsírfogyasztást módosítják.")
70
A táplálékfelvételt megindító és leállító perifériás jelzések
A táplálékfelvétel szabályozásában szereplő neuroncsoportokat kiterjedt glükózmonitorozó rendszer juttatja információhoz. A megkezdett táplálékfelvételt preabszortív jelzések állítják le. Az energiaraktárak állandóságának beállításához a zsírszövetből jövő jelzőmolekula, a leptin is hozzájárul A hideg környezetben, valamint fizikai aktivitás következtében megnövekszik a táplálékfelvétel A testtömeg hosszú távú szabályozásában mind a táplálékfelvétel, mind az energiafelhasználása megváltozása szerepel

71
Az eddigi kutatások sierint a noradrenerg pályák közül az alfa-2-recpetorn végződőd, a GBAerg, agalaninnal, az opioid peptidekkel és az NPY-nal műkődő pályák a táplálékfelvételt fokozzák A táplálékfelvételt gátolják: szerotonerg, noraedrenerg alfa-2-recperoon, deopaminerg gáolják.

72
A táplálékfelvételt megindító és leállító perifériás jelzések
A táplálékfelvétel ugyan araktárak szinten tratására szolgál, mégis alkalmazkoidk anapi szükségletekhez. A táplálékkeresést emberben illat. Látvány ismegindítja, de nem lesz hatásos, ha a központba olyan jelzések futnak be, amelyek a közelmúltban történt jelentős táplálékfelvételről tájékoztatnak. A táplálkozást azonban többnyire oylan jelzések indítják be,amelyek a táplálékfelvétel szükségességéről tájékoztták a hipotalamuszt. Ezek a jelzések a perifériáról származnak Éhség Jóllakottság, táplálkozás leállítása

73
Glükózmonitorozó rendszer
Az agytörzs, hipotalamusz, limbikus rendszer: Perifériás glükózreceptorok és centrális neuronok glükózérzékenysége folytán értesülnek a glükózszintről. Perifériás glükózreceptorok: vena portae, hasnyálmirigy, vékonybél – n. vagus végződései érzékelik a helyi glükózkoncentrácót. >>> nyúltvelő (nucleus tractus solitarii) >>> csökken a táplálékfelvétel Központi idegrendszer (amygdala, hypothalamus, nucleus tractus solitarii): glükóz szenzitív és glükózreceptor sejtek Együtetsen egy összehangolt rendszert alkotnak, ami ezt az információt összeveti egyéb jelzésekkel, így atáplálékfelvételi motivációt ennek megfelleően állítaj be. Mindezek ellenére nem igazolt, hogy a fiziológiás éhségérzetben és táplálékfelvételi kséztetésében a glükóz szint csökkenése lenne a megindítő tényező.
 >>> csökken a táplálékfelvétel. Központi idegrendszer (amygdala, hypothalamus, nucleus tractus solitarii): glükóz szenzitív és glükózreceptor sejtek. Együtetsen egy összehangolt rendszert alkotnak, ami ezt az információt összeveti egyéb jelzésekkel, így atáplálékfelvételi motivációt ennek megfelleően állítaj be. Mindezek ellenére nem igazolt, hogy a fiziológiás éhségérzetben és táplálékfelvételi kséztetésében a glükóz szint csökkenése lenne a megindítő tényező.")
74
A megkezdett táplálékfelvétel leállítása
Gyomor teltsége – mechanoreceptorokon keresztül kémiai tényezők: CCK >>> anorexigén hatású (n. vaguson közvettíéésvel , ill. közvetlen a központon), lassítja a gyomorürülést bombezin inzulinszekréció: gátolja az NPY gén átíródását >>> NPY csökken Az inzulinhiányos dibateses hyperphagia keletkezésében jelentős szerepe van! + felszívott glükóz, triptofán Leptin A zsírraktár nagyságának optimallizálást szolgálja a leptin-leptin-receptor rendszer. A leptin a szírszövetből származó polipeptid. A zsírszövet tömegének csökkenésekor a vér leptin koncentrációja csökken; a csökkenésre a hipotalamuszban fokozódik az NPY leadása. Ha a zsírszövet tömege megnövekszik,akkor több leptin kerül a vérbe. A leptin a hipotalamikus sejtek leptinreceptorainkersztül csökeknti atáplálékfelvételt, növeli az energiafelhasználást.
, lassítja a gyomorürülést. bombezin. inzulinszekréció: gátolja az NPY gén átíródását >>> NPY csökken. Az inzulinhiányos dibateses hyperphagia keletkezésében jelentős szerepe van! + felszívott glükóz, triptofán. Leptin. A zsírraktár nagyságának optimallizálást szolgálja a leptin-leptin-receptor rendszer. A leptin a szírszövetből származó polipeptid. A zsírszövet tömegének csökkenésekor a vér leptin koncentrációja csökken; a csökkenésre a hipotalamuszban fokozódik az NPY leadása. Ha a zsírszövet tömege megnövekszik,akkor több leptin kerül a vérbe. A leptin a hipotalamikus sejtek leptinreceptorainkersztül csökeknti atáplálékfelvételt, növeli az energiafelhasználást.")
75
Éhezés Adaptáció: - glükoneogenezis fenntartása
- összes triglicerid mennyisége, amelyből a szervezet biztosítani tudja alapműködéseit. A túlélést az egyes hormonok összehangolt szekréciója teszi lehetővé. - az éhezés valamennyi szakaszában cskken avérben az inzulin/glukagon arány. Mellékvese glukokortikoid hormonjai (főleg kortizon) >>> génexpresszió szabláyozáával biztosítják a glukoneogenetikus és a lipolítikus enzimek szintjét (permisszív hatás), de szekréciójuk az éhezés során nem változik! ( aglujkokortikoidok hiányában az éhező szerezet enm képes a glukoneogenezisre) 3 szakasza: posztabszorptív (max. 24 óráig) rövid távú: óra krónikus éhezés > 72 óra
 >>> génexpresszió szabláyozáával biztosítják a glukoneogenetikus és a lipolítikus enzimek szintjét (permisszív hatás), de szekréciójuk az éhezés során nem változik! ( aglujkokortikoidok hiányában az éhező szerezet enm képes a glukoneogenezisre) 3 szakasza: posztabszorptív (max. 24 óráig) rövid távú: óra. krónikus éhezés > 72 óra.")
76
Éhezés Posztabszorptív állapot (két táplálkozási ciklus között is megtörténik): - az inzulinszekréció csökken, a glukagon szekréció nő A vér glükóz szintjét 75%-ban a máj glikogenolízise, 25%-ban a glukoneogenezis tartja állandaón. A gükoneogenezis szubsztrátjai: vázizomból, vérsejtekből, agyból leadott tejsav, lipolízisből származó glicerin, kis mértékben aminosavak. minden szövetben, amelynek nem egyedüli energiaforrása a glükóz, az alcsony inzulin szint következtében csökken a glükóz felvétele a zsírsejtkben csökken a lipolízis gátlása (inzulin hatás), ezért nő avérbe kerülő szabad zsírsav és glicerin. a szabad zsírsavak részben a váz- és szívizomztaban használódhatnak fel a májban a még nagyon kevés ketontest keletkezik
, ezért nő avérbe kerülő szabad zsírsav és glicerin. a szabad zsírsavak részben a váz- és szívizomztaban használódhatnak fel. a májban a még nagyon kevés ketontest keletkezik.")
77
Éhezés rövid távú: folytatódik az inzulin szekréció további csökkenése, a glukagon és a növekedési hormon szekréció fokozódik – ezt a vérplazma csökkent glükóz koncentrációja indítja be A máj glikogénraktárának nagy rész lebomlott, a glükóz szintet a glukoneogenezis biztosítja (izomzat proteolíziséből származó aminosavak, glicerin és laktát a szubsztrát) a nitrogén-ürítés nő a lipolízis tovább fokozódik a legtöbb szövet a szabad zsírsavakat használja energiaforrásként (kivéve agy, vérsejtek) a megnövekedett glukagonszint és a fokozott zsírsavszint miatt a májban fokozódik a ketogenezis (éhezési ketonaemia) a ketontesteket a vázizom és szívizom használja, az agy még nem!
 a nitrogén-ürítés nő. a lipolízis tovább fokozódik. a legtöbb szövet a szabad zsírsavakat használja energiaforrásként (kivéve agy, vérsejtek) a megnövekedett glukagonszint és a fokozott zsírsavszint miatt a májban fokozódik a ketogenezis (éhezési ketonaemia) a ketontesteket a vázizom és szívizom használja, az agy még nem!")
78
Éhezés Hosszú távú, krónikus:
A szervezet összenergia-igénye is csökkenni kezd – inaktivitás, hormonális tényezők (pl. csökkent pajzsmirigy működés), leptintermelés, a szervek kezdődő állománycsökkenése; a 20%-ot is elérheti az inzulinszekréció tovább csökken, anövekedési hormonszekráció tovább nő lipolízis, ketogenezis fokozódik; vér ketontest szint 2-3 mmol/l is lehet >>> ennél aszintnél már az idegsejtek is képesek energiaszükségletüket részben ketontestek oxidációjából fdezni. így a glükóz felhasználás 50%-kal csökken, a glukoneogenezis iránti igény is kisebb lesz - az izomzatban a proteolzís csökken (ismeretlen ok), a nitrogén-ürítés csökken - Eben az állapotban - a zsírraktárak mennyiségétől függően – a szervezet hetekig élhet. A későbbiekben azonban a proteolízis már érinti alétfontosságú szövteket (pl. légzőizmok). Ha az éhezés tovább folytatódik, akkor azutolsó szakaszban ismét nő a nitrogén-ürítés, és ez az állpaot már irrevrezibilis.
, leptintermelés, a szervek kezdődő állománycsökkenése; a 20%-ot is elérheti. az inzulinszekréció tovább csökken, anövekedési hormonszekráció tovább nő. lipolízis, ketogenezis fokozódik; vér ketontest szint 2-3 mmol/l is lehet >>> ennél aszintnél már az idegsejtek is képesek energiaszükségletüket részben ketontestek oxidációjából fdezni. így a glükóz felhasználás 50%-kal csökken, a glukoneogenezis iránti igény is kisebb lesz. - az izomzatban a proteolzís csökken (ismeretlen ok), a nitrogén-ürítés csökken. - Eben az állapotban - a zsírraktárak mennyiségétől függően – a szervezet hetekig élhet. A későbbiekben azonban a proteolízis már érinti alétfontosságú szövteket (pl. légzőizmok). Ha az éhezés tovább folytatódik, akkor azutolsó szakaszban ismét nő a nitrogén-ürítés, és ez az állpaot már irrevrezibilis.")
79
Szomjúság A szervezet vízállománynak gyarapítása: ivással bevitt folyadék, étel, oxidációs víz A szomjúságérzet kiváltó tényezői: Az extracelluláris ozmolalitás emelkedése: - egészsége semberben aküszöb a plazma ozmolalitáásnak 2-3%-os emelkedése érzékelése: elülső hipotalamusz – angiotenzin II közvetítő szerepe; 290 mosm/kg víz ozmolalitás felett (az arginin-vazopreszin (AVP) szekréciója már 285 mosm/kg víz ozmolalitásnál fokozódik, tehát a vízvisszatartó mechanizmus érzékenyebb mint avízpótló. - a szájnyálkahártyából származó afferentáció si smodulálja az extracelluláris térfogat csökkenése
 szekréciója már 285 mosm/kg víz ozmolalitásnál fokozódik, tehát a vízvisszatartó mechanizmus érzékenyebb mint avízpótló. - a szájnyálkahártyából származó afferentáció si smodulálja. az extracelluláris térfogat csökkenése.")
80
Szomjúság az extracelluláris térfogat csökkenése:
akkor is szomjúságérzetet vált ki, ha az ozmolalitás nem nő vagy éppen csökken (pl. vérzés esetén, izoozmotikus folyadékvesztésnél vagy ascitesnél) a szomjúságérzetet generál neuronokat a keringési rendszer teltségét jelző alacsony nyomású baroreceptrok gátolják; a volumen csökkenésénél csökken a receptorok ingerülete és a szomjúságot kiváltó neuronok aktiválódnak a magas nyomású baroreceptorok ingerületcsökkenését más mechanizmus közvetíti: szimpatikus ingerület>>> fokozódik a vesében a reninszekréció >>> angiotenzin II közvetlenül hat a szomjúságot közvetítő neuronokra A folyadékfelvétel megszakítása – kevésbé tisztázott emberben a hiperozmózisal kiváltott vízfelvétel akkor szűnik meg, ha a vízdeficit felér perc alatt elfogyasztotta az egyén; a deficit máik felét perc alatt fogyasztja el a vizsgálati alany
 a szomjúságérzetet generál neuronokat a keringési rendszer teltségét jelző alacsony nyomású baroreceptrok gátolják; a volumen csökkenésénél csökken a receptorok ingerülete és a szomjúságot kiváltó neuronok aktiválódnak. a magas nyomású baroreceptorok ingerületcsökkenését más mechanizmus közvetíti: szimpatikus ingerület>>> fokozódik a vesében a reninszekréció >>> angiotenzin II közvetlenül hat a szomjúságot közvetítő neuronokra. A folyadékfelvétel megszakítása – kevésbé tisztázott. emberben a hiperozmózisal kiváltott vízfelvétel akkor szűnik meg, ha a vízdeficit felér perc alatt elfogyasztotta az egyén; a deficit máik felét perc alatt fogyasztja el a vizsgálati alany.")
Hasonló előadás



















 VEGETATÍV IDEGRENDSZER>")
