Előadást letölteni
Az előadás letöltése folymat van. Kérjük, várjon

1
Anyagtranszport és mechanizmusai
Dr. Horváth Ferenc SZTE, Növénybiológiai Tanszék

2
A membrántranszport alapjai 2. A membrántranszport szerveződése
Az anyagtranszportot négyféle hajtóerő mozgatja: koncentráció, hidrosztatikus nyomás, (gravitáció) és elektromos mező μj - j anyag (elektro)kémiai potenciálja μj0 – j anyag standard kémiai potenciálja R – egyetemes gázállandó (8,314 J mol-1 K-1) T – abszolút hőmérséklet (K) a – aktivitás, töltés nélküli oldatoknál: Cj moláris koncentráció Vj – j anyag parciális moláris térfogata P – hidrosztatikai nyomás z – az anyag töltése F- Faraday állandó (96500 coulomb / mol proton) E – elektromos potenciál A kémiai potenciál mértékegysége a J mol-1.
 és elektromos mező. μj - j anyag (elektro)kémiai potenciálja. μj0 – j anyag standard kémiai potenciálja. R – egyetemes gázállandó (8,314 J mol-1 K-1) T – abszolút hőmérséklet (K) a – aktivitás, töltés nélküli oldatoknál: Cj moláris koncentráció. Vj – j anyag parciális moláris térfogata. P – hidrosztatikai nyomás. z – az anyag töltése. F- Faraday állandó (96500 coulomb / mol proton) E – elektromos potenciál. A kémiai potenciál mértékegysége a J mol-1.")
3
A membrántranszport alapjai
2. A membrántranszport szerveződése Passzív transzport: A sejt nem fektet be metabolikus energiát az anyag felvételére. 1. A transzport hajtóereje: két pont között fennálló kémiai potenciál különbség →DIFFÚZIÓS MOZGÁS Passzív transzporttal mozog: a víz gázok: oxigén, széndioxid, N oxidok, kéndioxid stb. lipidoldékony, apoláros anyagok (benzin, szénhidrogének, aromás szennyeződések), ionok (bizonyos esetekben)
, ionok (bizonyos esetekben)")
4
A membrántranszport alapjai 2. A membrántranszport szerveződése
Elektromosan töltetlen anyagok kémiai potenciálja S anyag kémiai potenciálja: Két vizes teret határoló membrán esetén: A kémiai potenciál különbsége: Ez a koncentráció-különbségben raktározott energia. Mindig a citoplazmához viszonyítunk.

5
A membrántranszport alapjai
2. A membrántranszport szerveződése A transzport iránya DmS előjele fontos, ha pozitív – a citoplazmába az S anyag energiaigényes, aktív módon jut negatív – a citoplazmába az S anyag passzív módon jut 0 – nincs koncentráció-különbség

6
A membrántranszport alapjai
2. A membrántranszport szerveződése Vegyünk egy z töltéssel rendelkező iont (I): Elektrokémiai potenciál membránpotenciál
: Elektrokémiai potenciál. membránpotenciál.")
7
A membrántranszport alapjai
2. A membrántranszport szerveződése Aktív transzport A sejt metabolikus energiát fektet be az anyag felvételéhez. Elsődleges aktív transzport: ionpumpák Növényekben a proton-transzlokáló ATP-ázok (H+-ATP-áz) - lokalizáció: plazmamembrán, vakuólum membrán (tonoplaszt) - az ATP hidrolízisével felszabaduló energiát a proton sejtfalba ill vakuólumba irányuló transzportjára használja; Eredmény: proton elektrokémiai potenciál gradiens proton mozgató erő (pmf) Másodlagos aktív transzport: A pmf felhasználása más anyagok transzportjában - ezek a transzporterek lehetnek hordozók és csatornák
 - lokalizáció: plazmamembrán, vakuólum membrán (tonoplaszt) - az ATP hidrolízisével felszabaduló energiát a proton sejtfalba ill. vakuólumba irányuló transzportjára használja; Eredmény: proton elektrokémiai potenciál gradiens proton mozgató erő (pmf) Másodlagos aktív transzport: A pmf felhasználása más anyagok transzportjában. - ezek a transzporterek lehetnek hordozók és csatornák.")
8
A membrántranszport alapjai 2. A membrántranszport szerveződése
Proton mozgató erő (pmf): a protonra vonatkozó transzmembrán elektrokémiai potenciálkülönbség, voltokban kifejezve z = +1 A pmf-et voltokban fejezzük ki és nem J/mol egységekben, így Tehát 1 pH egység különbség 59 mV elektrokémiai potenciálváltozást jelent a protonmozgató erőben!
: a protonra vonatkozó transzmembrán elektrokémiai potenciálkülönbség, voltokban kifejezve. z = +1. A pmf-et voltokban fejezzük ki és nem J/mol egységekben, így. Tehát 1 pH egység különbség 59 mV elektrokémiai potenciálváltozást jelent a protonmozgató erőben!")
9
A membrántranszport alapjai 2. A membrántranszport szerveződése
pH: 7,5 Vm = -150 mV DpH = -2 pmf = -268 mV pH: 5,5 A protonpumpák működése elektrogén: így nem csak a pH különbség előállításán keresztül növelik a pmf abszolút értékét, hanem a Vm értékét is negatívabbá teszik. A proton elektrokémiai potenciál-grádiensében rejlő energia a másodlagos aktív transzportban hasznosul

10
Kemiozmotikus hipotézis A membrántranszport alapjai
2. A membrántranszport szerveződése Kemiozmotikus hipotézis Peter Mitchell 1960-as évek (Nobel díj, 1978). A sejt energiatermelése és a H+ elektrokémiai potenciál-gradiense közötti összefüggés. ATP <-> ADP+Pi+pmf
. A sejt energiatermelése és a H+ elektrokémiai potenciál-gradiense közötti összefüggés. ATP <-> ADP+Pi+pmf.")
11
A membrántranszport alapjai 2. A membrántranszport szerveződése
Az ionpumpák átviteli száma kicsi, és pmf-et generál a hordozók számára is, ezért jóval nagyobb a koncentrációja a membránban, mint a csatornáké. Ionpumpák Hordozók Ioncsatornák Átviteli szám (db molkekula/másodperc) 102 103 106 – 108 Membránon vett sűrűség (db/mm2) 10-100 1-10
 – 108. Membránon vett sűrűség (db/mm2)")
12
Pumpák A membrántranszport alapjai 3. Pumpák F-típusú H+-ATPázok
belső mitokondriális és tilakoid membrán P-típusú H+-ATPázok gomba PM H+-ATPáz növény PM H+-ATPáz Na+ / K+ ATPáz (állati sejtek) Ca2+-ATPázok (növény és állat PM és endomembránok) H+ / K+ cserélő ATPáz (emlős gyomor mucosa réteg) V-típusú H+-ATPázok Vakuoláris proton pirofoszfatáz (H+-PPáz) ABC-típusú pumpák
 Ca2+-ATPázok (növény és állat PM és endomembránok) H+ / K+ cserélő ATPáz (emlős gyomor mucosa réteg) V-típusú H+-ATPázok. Vakuoláris proton pirofoszfatáz (H+-PPáz) ABC-típusú pumpák.")
13
A membrántranszport alapjai 3. Pumpák
F-típusú H+-ATPázok a belső mitokondrális membránon és a tilakoid membránon találhatók a redoxpotenciál ill. a fény energiájából pmf keletkezik a pmf hajtja a H+ áramlást az F-típusú ATPázon keresztül ATP szintetizálódik F1 (mitokondrium) CF1 (kloroplasztisz) alegység a3b3gde 3 db b alegység: 3 nukleotidkötő domén F0 (mitokondrium) CF0 (kloroplasztisz) alegység ab2c9-12 A c alegységek forognak a proton transzlokáció során, ami a g alegységet pörgeti, így a b nukleotidkötő helyek konformációváltozást szenvednek..
 CF1 (kloroplasztisz) alegység. a3b3gde. 3 db b alegység: 3 nukleotidkötő domén. F0 (mitokondrium) CF0 (kloroplasztisz) alegység. ab2c9-12. A c alegységek forognak a proton transzlokáció során, ami a g alegységet pörgeti, így a b nukleotidkötő helyek konformációváltozást szenvednek..")
14
A membrántranszport alapjai
3. Pumpák John Walker és Paul Boyer (Nobel díj, 1997) Kötésváltoztató mechanizmus Gyenge kötődés: az aktív centrum gyengén köti az ADP-t és Pi-t Erős kötődés: az ATP molekula kialakul Nyitott konformáció: ATP leválás 3 db H+ átjutása alatt szintetizálódik 1 ATP molekula.
 Kötésváltoztató mechanizmus. Gyenge kötődés: az aktív centrum gyengén köti az ADP-t és Pi-t. Erős kötődés: az ATP molekula kialakul. Nyitott konformáció: ATP leválás. 3 db H+ átjutása alatt szintetizálódik 1 ATP molekula.")
15
A membrántranszport alapjai 3. Pumpák
P-típusú ATPázok családja 1 db 100 kDa-os protein, ATP-t köt és H+ transzportot katalizál Funkciói: - pmf (Vm) generálása (hordozók és ioncsatornák) - sejtfal savanyítás (auxin hozzáadása után 2 perccel!) expanzin enzimek aktiválódnak, amelyek a H-kötéseket lazítják – sejtfal növekedés - H+ eltávolítása a sejtből (anyagcsere folytonosan termeli) - a citoszol pH-jának szabályozása (7,3-7,5): a H+-ATPáz pH-optimuma 6,6, tehát ha savas a közeg, akkor jobban működik Hatékonysága: 1 proton / 1 MgATP hidrolízise Reakcióciklus: E1 konformáció – H+ kötődik hozzá Az ATP hidrolízise, és a foszfát-csoport enzimhez való kapcsolódása az E2 konformáció kialakulását eredményezi. E2 alacsony affinitású a H+-ra nézve, így az a túloldalon leválik. Az enzim-foszfát kötés hidrolízise során Pi disszociálódik, és az enzim E1 konformációba tér vissza.
 generálása (hordozók és ioncsatornák) - sejtfal savanyítás (auxin hozzáadása után 2 perccel!) expanzin enzimek aktiválódnak, amelyek a H-kötéseket lazítják – sejtfal növekedés. - H+ eltávolítása a sejtből (anyagcsere folytonosan termeli) - a citoszol pH-jának szabályozása (7,3-7,5): a H+-ATPáz pH-optimuma 6,6, tehát ha savas a közeg, akkor jobban működik. Hatékonysága: 1 proton / 1 MgATP hidrolízise. Reakcióciklus: E1 konformáció – H+ kötődik hozzá. Az ATP hidrolízise, és a foszfát-csoport enzimhez való kapcsolódása az E2 konformáció kialakulását eredményezi. E2 alacsony affinitású a H+-ra nézve, így az a túloldalon leválik. Az enzim-foszfát kötés hidrolízise során Pi disszociálódik, és az enzim E1 konformációba tér vissza.")
16
A membrántranszport alapjai 3. Pumpák
A foszfátcsoport kovalens kötődése (E-P) megkülönbözteti az F-típusú ATPázoktól. Az enzimcsalád tagjai: gomba PM H+-ATPáz növény PM H+-ATPáz Na+ / K+ ATPáz (állati sejtek) (Skou, Nobel díj, 1997) Ca2+-ATPázok (növény és állat PM és endomembránok) H+ / K+ cserélő ATPáz (emlős gyomor mucosa réteg) Közös tulajdonságok: ortovanadáttal (H2VO4-) való gátolhatóság domén struktúra azonossága (főleg az ATP-kötő domén konzervált a különböző pumpák között) ATP-kötő domén A defoszforilációban résztvevő hurok D – aszparaginsav (foszforilálódik) Autoinhibíciós domén
 megkülönbözteti az F-típusú ATPázoktól. Az enzimcsalád tagjai: gomba PM H+-ATPáz. növény PM H+-ATPáz. Na+ / K+ ATPáz (állati sejtek) (Skou, Nobel díj, 1997) Ca2+-ATPázok (növény és állat PM és endomembránok) H+ / K+ cserélő ATPáz (emlős gyomor mucosa réteg) Közös tulajdonságok: ortovanadáttal (H2VO4-) való gátolhatóság. domén struktúra azonossága (főleg az ATP-kötő domén konzervált a különböző pumpák között) ATP-kötő domén. A defoszforilációban résztvevő hurok. D – aszparaginsav (foszforilálódik) Autoinhibíciós domén.")
17
A membrántranszport alapjai 3. Pumpák
A P-típusú ATPázokat egy multigén-család kódolja, mely szövetspecifikus expressziót mutat Arabidopsis (lúdfű) esetén AHA géncsalád (10 tag, 10 izoformát kódol) pl. AHA3 – floém AHA10 fejlődő magvak Magyarázat: más és más az ATP-re vonatkozó KM érték, más az ortovanadátra való érzékenység Szövetspecifikus P-típusú ATPáz génexpresszió Arabidopsisban. szárkeresztmetszet, az AHA3-c-Myc fúziós protein immunofluoreszcens kimutatása P – floém, C – kortikális sejtek, X – xilém AHA10 gén promóterének expressziója fejlődő magvakban, b-glükuronidáz (GUS) festéssel. A nyilak két magot mutatnak a becőtermésben. A kék szín az AHA10-GUS fúziós protein termelődését mutatja.
 esetén AHA géncsalád (10 tag, 10 izoformát kódol) pl. AHA3 – floém. AHA10 fejlődő magvak. Magyarázat: más és más az ATP-re vonatkozó KM érték, más az ortovanadátra való érzékenység. Szövetspecifikus P-típusú ATPáz génexpresszió Arabidopsisban. szárkeresztmetszet, az AHA3-c-Myc fúziós protein immunofluoreszcens kimutatása P – floém, C – kortikális sejtek, X – xilém. AHA10 gén promóterének expressziója fejlődő magvakban, b-glükuronidáz (GUS) festéssel. A nyilak két magot mutatnak a becőtermésben. A kék szín az AHA10-GUS fúziós protein termelődését mutatja.")
18
A membrántranszport alapjai 3. Pumpák
A P-típusú ATPázok szabályozása - pH-tól való függés - a C-terminális autoinhibíciós domén (ennek eltávolítása vagy pontmutáció benne módosítja az enzim működését) - az auxinnak hatása a pumpa expressziójának növelésében van A fuzikokcin (Fusicoccum amygdali gomba toxinja) – növeli a zárósejtek turgorát sztómanyitódás, levélszáradás 1994 – a fuzikokcin receptora a szignál-transzdukciós proteinek családjából való protein proteinek – dimerek, melyek a célproteinre (foszforilált szerin reziduálist tartalmaz) vonatkozó konszenzus szekvenciával rendelkeznek
 - az auxinnak hatása a pumpa expressziójának növelésében van. A fuzikokcin (Fusicoccum amygdali gomba toxinja) – növeli a zárósejtek turgorát sztómanyitódás, levélszáradás – a fuzikokcin receptora a szignál-transzdukciós proteinek családjából való protein proteinek – dimerek, melyek a célproteinre (foszforilált szerin reziduálist tartalmaz) vonatkozó konszenzus szekvenciával rendelkeznek.")
19
A membrántranszport alapjai 3. Pumpák
A C terminus autoinhibíciós doménként viselkedik. A szerin foszforilációja és a kötődő 2 db protein megszünteti a gátlást és aktiválja az enzimet (balra) Az enzimet foszforiláció híján a fuzikokcin proteinek is aktiválhatják
 Az enzimet foszforiláció híján a fuzikokcin proteinek is aktiválhatják.")
20
A membrántranszport alapjai
3. Pumpák Na+ / K+ ATPáz (állati sejtek)
")
21
A membrántranszport alapjai 3. Pumpák
A Ca2+-ATPáz Megtalálható: PM, ER, kloroplasztisz membrán, vakuoláris membránok Ca2+-ot pumpálnak ki a citoszolból, [Ca2+]cyt = 0,2 mM Állati sejtekben részletesen tanulmányozott pumpák: PM-típusú ER-típusú Aktiváció a kalmodulin a C-terminálishoz kötődik nincs kalmodulin kötőhely Helye növényekben PM, tonoplaszt (itt kivételesen a kalmodulin kötőhely az N-terminálison van), kloroplasztisz belső membrán ER A Ca2+ átjuttatásához sok energia kell, mert: - kint több Ca2+ van mint a citoszolban - a citoszol elektromosan negatívabb, mint a külső oldal A Ca2+ eletrokémiai potenciálkülönbségéből származó szabadenergia: -60 kJ/mol Az ATP hidrolíziséből származó szabadenergia: -50 kJ/mol, ami nem elég a pumpa működtetéséhez. Gyakran Ca2+ / H+ cserélőként működik! Vakuoláris PM-típusú Ca2+-ATPáz
, kloroplasztisz belső membrán. ER. A Ca2+ átjuttatásához sok energia kell, mert: - kint több Ca2+ van mint a citoszolban. - a citoszol elektromosan negatívabb, mint a külső oldal. A Ca2+ eletrokémiai potenciálkülönbségéből származó szabadenergia: -60 kJ/mol. Az ATP hidrolíziséből származó szabadenergia: -50 kJ/mol, ami nem elég a pumpa működtetéséhez. Gyakran Ca2+ / H+ cserélőként működik! Vakuoláris PM-típusú Ca2+-ATPáz.")
22
A membrántranszport alapjai 3. Pumpák
A vakuoláris (V-típusú) H+-ATPázok csoportja Funkciója: a vakuoláris tér savanyítása kb. pH 5,5-ös értékre (citrusfélék gyümölcsének esetén ez akár pH 3 alatt is lehet) nemcsak a hordozók számára energizálja a membránt, de sok vakuoláris enzim (proteázok, glükozidázok, foszfatázok, nukleotidázok) pH optimuma savas Az F-típusú ATPázok rokonai, de fordított irányban működnek. Sokkal összetettebb alegység szerkezet, mint az F-típusnál. Sztöchiometria: 2 db H+ transzlokáció / 1 ATP hidrolízis Gátolhatóság: bafilomicin A1, a V0 szektorral lép kölcsönhatásba Előfordulása: tonoplaszt, ER, Golgi, burkolt vezikulumok membránja (a mitokondriumon és a kloroplasztiszon kívül minden organellum képes fenntartani a savas belső terét. V1 V0
 H+-ATPázok csoportja. Funkciója: a vakuoláris tér savanyítása kb. pH 5,5-ös értékre. (citrusfélék gyümölcsének esetén ez akár pH 3 alatt is lehet) nemcsak a hordozók számára energizálja a membránt, de. sok vakuoláris enzim (proteázok, glükozidázok, foszfatázok, nukleotidázok) pH optimuma savas. Az F-típusú ATPázok rokonai, de fordított irányban működnek. Sokkal összetettebb alegység szerkezet, mint az F-típusnál. Sztöchiometria: 2 db H+ transzlokáció / 1 ATP hidrolízis. Gátolhatóság: bafilomicin A1, a V0 szektorral lép kölcsönhatásba. Előfordulása: tonoplaszt, ER, Golgi, burkolt vezikulumok membránja (a mitokondriumon és a kloroplasztiszon kívül minden organellum képes fenntartani a savas belső terét. V1. V0.")
23
A membrántranszport alapjai 3. Pumpák
Vakuoláris proton pirofoszfatáz (H+-PPáz) A szervetlen pirofoszfát (PPi) hidrolíziséből felszabaduló energiát hasznosítja Szerkezete: egyszerű, kicsi (80 kDa)-os fehérje, 16 transzmembrán szegmenssel Szubsztrátja a dimagnézium-pirifoszfát (citoplazmában mM-os nagyságrendben) Gátolhatósága: Ca2+, aminometilén-difoszfonát, a működéséhez a citoplazmatikus felszínnél K+ ionoknak kell jelen lenni Miért van szükség kétféle protonpumpára a tonoplaszton? Fiatal sejtekben sok PPi termelődik a hidrolízise során felszabaduló hő helyett az energia inkább a pmf létrehozásában hasznosul
 A szervetlen pirofoszfát (PPi) hidrolíziséből felszabaduló energiát hasznosítja. Szerkezete: egyszerű, kicsi (80 kDa)-os fehérje, 16 transzmembrán szegmenssel. Szubsztrátja a dimagnézium-pirifoszfát (citoplazmában mM-os nagyságrendben) Gátolhatósága: Ca2+, aminometilén-difoszfonát, a működéséhez a citoplazmatikus felszínnél K+ ionoknak kell jelen lenni. Miért van szükség kétféle protonpumpára a tonoplaszton Fiatal sejtekben sok PPi termelődik a hidrolízise során felszabaduló hő helyett az energia inkább a pmf létrehozásában hasznosul.")
24
A membrántranszport alapjai 3. Pumpák
ABC-típusú pumpák Az amfipatikus molekulák vakuoláris membránon való átjutását katalizálják pl. flavonoidok, antocianinok, a klorofill lebontási melléktermékei, xenobiotikumok (herbicidek). Az átjutáshoz ATP szükséges, a transzport viszont nem csökkenti a pmf-t. ABC = ATP binding casette – elterjedt az enzimek között, melyek ATP-t kötnek. Walker A és B motívum NBF = nucleotide-binding fold (2 db van belőle) Sok transzmembrán szegmens
. Az átjutáshoz ATP szükséges, a transzport viszont nem csökkenti a pmf-t. ABC = ATP binding casette – elterjedt az enzimek között, melyek ATP-t kötnek. Walker A és B motívum. NBF = nucleotide-binding fold (2 db van belőle) Sok transzmembrán szegmens.")
25
A membrántranszport alapjai 3. Pumpák
ABC-típusú pumpák A flavonoidok és xenobiotikumok glutation-konjugátumként (GS-konjugátumok) transzlokálódnak. GS=tripeptid (glutaminsav+cisztein+glicin) Glutation S-transzferáz (GST) enzim végzi a konjugátum-képzést. Az ABC-transzporterhez való kötődés után 1, pumpaszerű működés, vagy 2, flippázszerű működés. Nem tisztázott. glutation (GS) AtMRP2 – Arabidopsis ABC-transzporter DNP – dinitrofenol (xenobiotikum) NCC – lineáris tetrapirrol Bn – Brassica napus
 transzlokálódnak. GS=tripeptid (glutaminsav+cisztein+glicin) Glutation S-transzferáz (GST) enzim végzi a konjugátum-képzést. Az ABC-transzporterhez való kötődés után. 1, pumpaszerű működés, vagy. 2, flippázszerű működés. Nem tisztázott. glutation (GS) AtMRP2 – Arabidopsis ABC-transzporter. DNP – dinitrofenol (xenobiotikum) NCC – lineáris tetrapirrol. Bn – Brassica napus.")
26
Hordozók A membrántranszport alapjai 4. Hordozók
pmf anyag H+ Hordozók uniport szimport v. antiport uniport – elektrokémiai gradiensen lefelé (facilitált diffúzió) kotranszport – elektrokémiai gradiensen fölfelé szimport antiport Másodlagos aktív transzport: az elsődleges aktív transzportban generált pmf használódik fel az anyag energiaigényes transzportjára Energiát szolgáltatja: pmf (H+), Na+ és több töltés nélküli anyag is
 kotranszport – elektrokémiai gradiensen fölfelé. szimport. antiport. Másodlagos aktív transzport: az elsődleges aktív transzportban generált pmf használódik fel az anyag energiaigényes transzportjára. Energiát szolgáltatja: pmf (H+), Na+ és több töltés nélküli anyag is.")
27
A membrántranszport alapjai 4. Hordozók
A hordozók működése Michaelis-Menten kinetikát mutat, mely konformációs változásra utal - a transzport alatt nem történik kémiai módosulás, - a transzport kinetikája szubsztráttal való telítődést mutat - a Michaelis-Menten kinetikával kezelhető A maximális sebesség felénél mérhető szubsztrát koncentráció a Michaelis-Menten állandó (KM) szubsztrát Lineáris szakasz: Alacsony S koncentráción az S hordozóhoz való kötődése alakítja a kinetikát Magasabb S koncentráción a konformációs változás, a kötőhelyhez való hozzáférhetőség a limitáló tényező
 szubsztrát. Lineáris szakasz: Alacsony S koncentráción az S hordozóhoz való kötődése alakítja a kinetikát. Magasabb S koncentráción a konformációs változás, a kötőhelyhez való hozzáférhetőség a limitáló tényező.")
28
A membrántranszport alapjai
4. Hordozók A másodlagos aktív transzport molekuláris mechanizmusa (itt szimport esetén)
")
29
Másodlagos aktív transzport: szimport és antiport
A membrántranszport alapjai 4. Hordozók Másodlagos aktív transzport: szimport és antiport A kotranszportált anyag lehet semleges és töltéssel rendelkező is

30
A membrántranszport alapjai 4. Hordozók - A kálium felvétele
nagy affinitású rendszer: K+-H+ szimport (1:1) HKT1 (534 aminosav protein) μM [K+]ext tartományban gyökér kortex sejtekben alacsony affinitású rendszer: befelé egyenirányító ioncsatornák 1 mM [K+]ext fölött Vm-hajtotta K+ influx (H+-ATPáz!) Al3+ ionok gátolják
 HKT1 (534 aminosav protein) μM [K+]ext tartományban. gyökér kortex sejtekben. alacsony affinitású rendszer: befelé egyenirányító ioncsatornák. 1 mM [K+]ext fölött. Vm-hajtotta K+ influx (H+-ATPáz!) Al3+ ionok gátolják.")
31
A membrántranszport alapjai 4. Hordozók
Más, nem ionhoz kapcsolt kotranszport pl. vakuoláris glükóz és aminosav hordozók Kloroplasztisz: 2-oxoglutársav-almasav, ATP-ADP, hexóz-foszfát-ortofoszfát, DHAP-ortofoszfát Ekkor a transzportált anyag koncentráció-gradiense szolgáltatja az energiát.

32
A hordozók szerkezete A membrántranszport alapjai 4. Hordozók
méretük kDa, többnyire hidrofób fehérjék általában 12 transzmembrán domén, a 6-7 között erősen hidrofób loop több belső repeat szekvencia a C és N-terminálisfélben (az idők során génduplikáció történt) MFS (main facilitator superfamily)-ba tartoznak, mely nagy és erősen diverz csoport
 MFS (main facilitator superfamily)-ba tartoznak, mely nagy és erősen diverz csoport.")
33
A membrántranszport alapjai 4. Hordozók
pmf H+ Sz Reakció: e c n: sztöchiometriai arány z: átvitt anyag töltése szimporter A reakció akkor játszódik le balról jobbra (akkor juttat a szimporter befelé protont és S anyagot), ha a két elektrokémiai potenciálgradiens vektori összege befelé mutat, azaz (a citoplazmához viszonyítunk) Kifejtve a két kémiai potenciált:
, ha a két elektrokémiai potenciálgradiens vektori összege befelé mutat, azaz. (a citoplazmához viszonyítunk) Kifejtve a két kémiai potenciált:")
34
A membrántranszport alapjai 4. Hordozók
Ha pl. pHcyt = 7,5 pHextracell = 5,5 akkor az anyag maximális felhalmozódási aránya: Ha pl. S töltés nélküli anyag, és 1 protonnal transzportálódik, akkor n = 1 esetén Vm= -150 mV mellett a max. felhalmozódási arány n = 2 esetén ez 1,34 x 109 ! Tehát a protonhoz kapcsolt hordozók nagy koncentrációkülönbséget képesek létrehozni, szabályozásuk nagyon fontos.

35
A membrántranszport alapjai
4. Hordozók Néhány esetben a hordozók működése nem H+-hoz hanem Na+-hoz kötött. Tengerben élő algafajok esetén az NO3- és néhány aminosav felvétele Na+-hoz kötött szimportot mutat. Tengervíz 480 mM Na! Édesvízi algáknál a Na/K szimport is lehetséges, a Na-ra vonatkozó nagy Vm komponens miatt.

36
Ioncsatorna A membrántranszport alapjai
5. Az ioncsatornák általános tulajdonságai Ioncsatorna Minden sejt minden membránján megtalálható. Két konformációs állapot: nyitott és zárt. Az állapotok közti átmenetet a membránpotenciál és a ligandum kötődése szabályozza (kapuzási faktorok). Teljes sejt méréseknél az teljes ionáramot fel lehet írni: I = N · i · Po , ahol N – a csatornák száma i – 1 db nyitott csatornán átfolyó áram Po – annak a valószínűsége, hogy a csatorna nyitott állapotban van.
. Teljes sejt méréseknél az teljes ionáramot fel lehet írni: I = N · i · Po , ahol. N – a csatornák száma. i – 1 db nyitott csatornán átfolyó áram. Po – annak a valószínűsége, hogy a csatorna nyitott állapotban van.")
37
A membrántranszport alapjai 5. Az ioncsatornák általános tulajdonságai
Az ioncsatornán való átjutás passzív: csak tól függ. Az egycsatorna-áram membránpotenciáltól való függése lineáris: Érvényes rá Ohm-törvénye: I = V / R Meredeksége = 1 / R = g (konduktancia) [pS] g értéke függ a permeáló ion koncentrációjától, tehát a csatorna konduktanciáját adott ionkoncentráció-viszony mellett adják meg. Az áram iránya: negatív áram – kation influx vagy anion efflux pozitív áram – kation efflux vagy anion influx
 [pS] g értéke függ a permeáló ion koncentrációjától, tehát a csatorna konduktanciáját adott ionkoncentráció-viszony mellett adják meg. Az áram iránya: negatív áram – kation influx vagy anion efflux. pozitív áram – kation efflux vagy anion influx.")
38
A membrántranszport alapjai
5. Az ioncsatornák általános tulajdonságai Equilibrium esetén nincs nettó ionáramlás: Eeq,ion az ion egyensúlyi potenciálja, csak az ion koncentráció-különbségétől függ. Ha csak egyféle ion van a rendszerben, ezt Nernst-potenciálnak nevezzük. Az egyensúly itt azt jelenti, hogy az iont adott irányba mozgató koncentráció-gradiens nagysága megegyezik az ellenkező irányban ható elektromos potenciál-gradiens nagyságával. Ha többféle permeáló ion (legfontosabb a Na+, K+, Cl-) van a membrán két oldalán, akkor a Goldman-Hodgkin-Katz egyenlet leírja a nyugalmi membránpotenciált: P – permeabilitás
 van a membrán két oldalán, akkor a Goldman-Hodgkin-Katz egyenlet leírja a nyugalmi membránpotenciált: P – permeabilitás.")
39
A Nernst-egyenletből számított érték
megfigyelt érték A K+ ionok a citoszolba és a vakuólumba passzív módon jutnak, kivéve, ha az extracelluláris K + koncentráció alacsony (aktív módon hordozóval) A Na+ aktív módon pumpálódik a citoszolból kifelé az extracelluláris térbe és a vakuólumba A köztes anyagcsere protonjai aktív módon pumpálódnak ki a citoszolból (megőrződik a citoszol semleges pH-ja, míg az extracelluláris tér és a vakuólum 1-2 pH egységgel savanyodik) Minden anion aktív módon jut a citoszolba A Ca2+ aktív módon transzportálódik ki a citoszolból
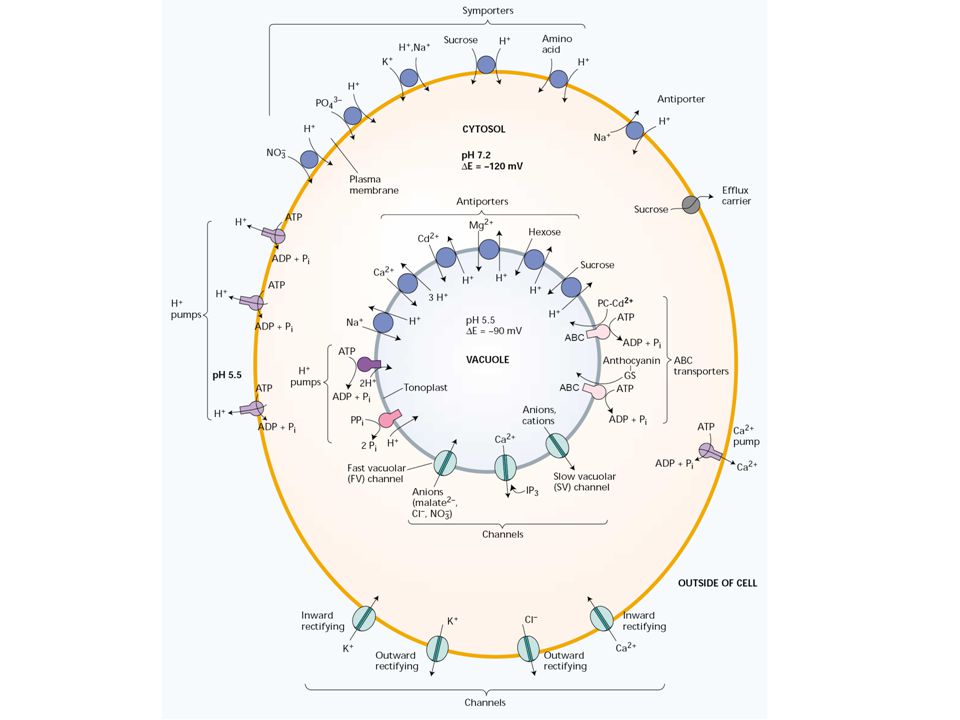
 A Na+ aktív módon pumpálódik a citoszolból kifelé az extracelluláris térbe és a vakuólumba. A köztes anyagcsere protonjai aktív módon pumpálódnak ki a citoszolból (megőrződik a citoszol semleges pH-ja, míg az extracelluláris tér és a vakuólum 1-2 pH egységgel savanyodik) Minden anion aktív módon jut a citoszolba. A Ca2+ aktív módon transzportálódik ki a citoszolból.")
41
Kapuzási kölcsönhatás
A membrántranszport alapjai 5. Az ioncsatornák általános tulajdonságai Kapuzási faktor Csatorna Membrán Kapuzási kölcsönhatás Csatorna funkció Fizikai: Feszültség Ca2+, K+, Cl-, M+, X-, N PM, T, C, M de-/aktiváció időfüggő inaktiváció Feszültség stabilitás, ingerelhetőség, ozmotikus anyagmozgás Mechanikai stressz Ca2+, K+, Cl-, M+, X- PM, T Aktiváció Turgor/ozmoreguláció Kémiai (ligandum): Ca2+ K+, Cl- PM In-/aktiváció ingerelhetőség, ozmotikus anyagmozgás T ozmotikus anyagmozgás, szignál-transzdukció K+ Feszültségfüggő aktiváció Elektrokémiai potenciál integráció, ozmotikus anyagmozgás Cl- ozmotikus anyagmozgás H+ Elektrokémiai potenciál integráció Inaktiváció ? IP3 Ca2+ felszabadítás, szignál-transzdukció Nukleotidok K+, X- PM, T, M Auxin X- Kalmodulin Ca2+, M+, X-, N ingerelhetőség, turgor- és ozmoreguláció, szignál-transzdukció + foszforiláció, és diszulfid hidak kialakítása
: Ca2+ K+, Cl- PM. In-/aktiváció. ingerelhetőség, ozmotikus anyagmozgás. T. ozmotikus anyagmozgás, szignál-transzdukció. K+ Feszültségfüggő aktiváció. Elektrokémiai potenciál integráció, ozmotikus anyagmozgás. Cl- ozmotikus anyagmozgás. H+ Elektrokémiai potenciál integráció. Inaktiváció. IP3. Ca2+ felszabadítás, szignál-transzdukció. Nukleotidok. K+, X- PM, T, M. Auxin. X- Kalmodulin. Ca2+, M+, X-, N ingerelhetőség, turgor- és ozmoreguláció, szignál-transzdukció. + foszforiláció, és diszulfid hidak kialakítása.")
42
Patch clamp mérési elrendezések
A membrántranszport alapjai 5. Ioncsatornák Patch clamp mérési elrendezések -10 mV -140 mV +60 mV

43
A membrántranszport alapjai
5. Ioncsatornák Patch clamp labor

44
A membrántranszport alapjai
5. Ioncsatornák Patch clamp labor

45
Sztómaműködés, a zárósejt
A membrántranszport alapjai Sztómamozgás Sztómaműködés, a zárósejt Működésének célja: a növény CO2 igényét a vízveszteség minimalizálása mellett elégítse ki Víz és gázcsere, levélhőmérséklet, ionfelvétel és transzport Hidroaktív nyitódás és záródás (fény, hőmérséklet, CO2, páratartalom, növényi hormonok)
")
46
A membrántranszport alapjai
Sztómamozgás A sztómanyílás méretének, a zárósejt K+ és szacharóz-tartalmának alakulása a nap folyamán K+ Szacharóz

47
Befelé és kifelé mutató kálium áramok zárósejtekben
A membrántranszport alapjai Sztómamozgás Befelé és kifelé mutató kálium áramok zárósejtekben K+ H2O K+ H2O (KAT1) K+out áram Áram t K+in áram Feszültség -140 mV + 60 mV
 K+out áram. Áram. t. K+in áram. Feszültség mV mV.")
48
Sztómanyitódás Sztómazáródás Kék fény R S 1. H+-pumpa aktiváció
ATP H+ ADP Pi + ATP H+ ADP Pi + Kék fény Ca2+ R Ca2+ Cl- S Cl- H+ depolarizáció [K+] 100 mM 800 mM Cl- K+ K+ 1. H+-pumpa aktiváció 2. membrán hiperpolarizáció 3. K+, Cl- és Ca2+ influx K+ 1. H+-pumpa inaktiváció 2. S és R-típusú anion-csatorna aktiváció 3. hosszan tartó PM depolarizáció 4. K+ efflux

49
Anyagtranszport a gyökérben
A membrántranszport alapjai Anyagtranszport a gyökérben Anyagtranszport a gyökérben

50
Apoplaszt és szimplaszt
A membrántranszport alapjai Anyagtranszport a gyökérben Apoplaszt és szimplaszt

51
A membrántranszport alapjai
Anyagtranszport a gyökérben A plazmodezmák olyan 40 nm-es átmérőjű citoplazma-hidak, melyek a víz és kisebb oldott anyagok diffúzióját teszik lehetővé a sejtek között. Sűrűségük a membránban 15 db/μm2.

52
Passzív diffúziós modell
A membrántranszport alapjai Anyagtranszport a gyökérben Passzív diffúziós modell

53
A membrántranszport alapjai
Anyagtranszport a gyökérben A xilém feltöltés passzív folyamat, vagy irányított transzport-mechanizmus? Válasz: xilém parenchima PM szabályozott transzport: K-specifikus és nem-szelektív kation efflux csatornák, melyek Vm és Ca2+-függőek a szabályozás független az epidermisz és kortikális sejtek aktív anyagfelvételétől Izotóppal jelölt transzport

Hasonló előadás